Changes in Soil Microbial Community and Its Effect on Carbon Sequestration Following Afforestation on the Loess Plateau, China


Abstract
:1. Introduction
2. Materials and Methods
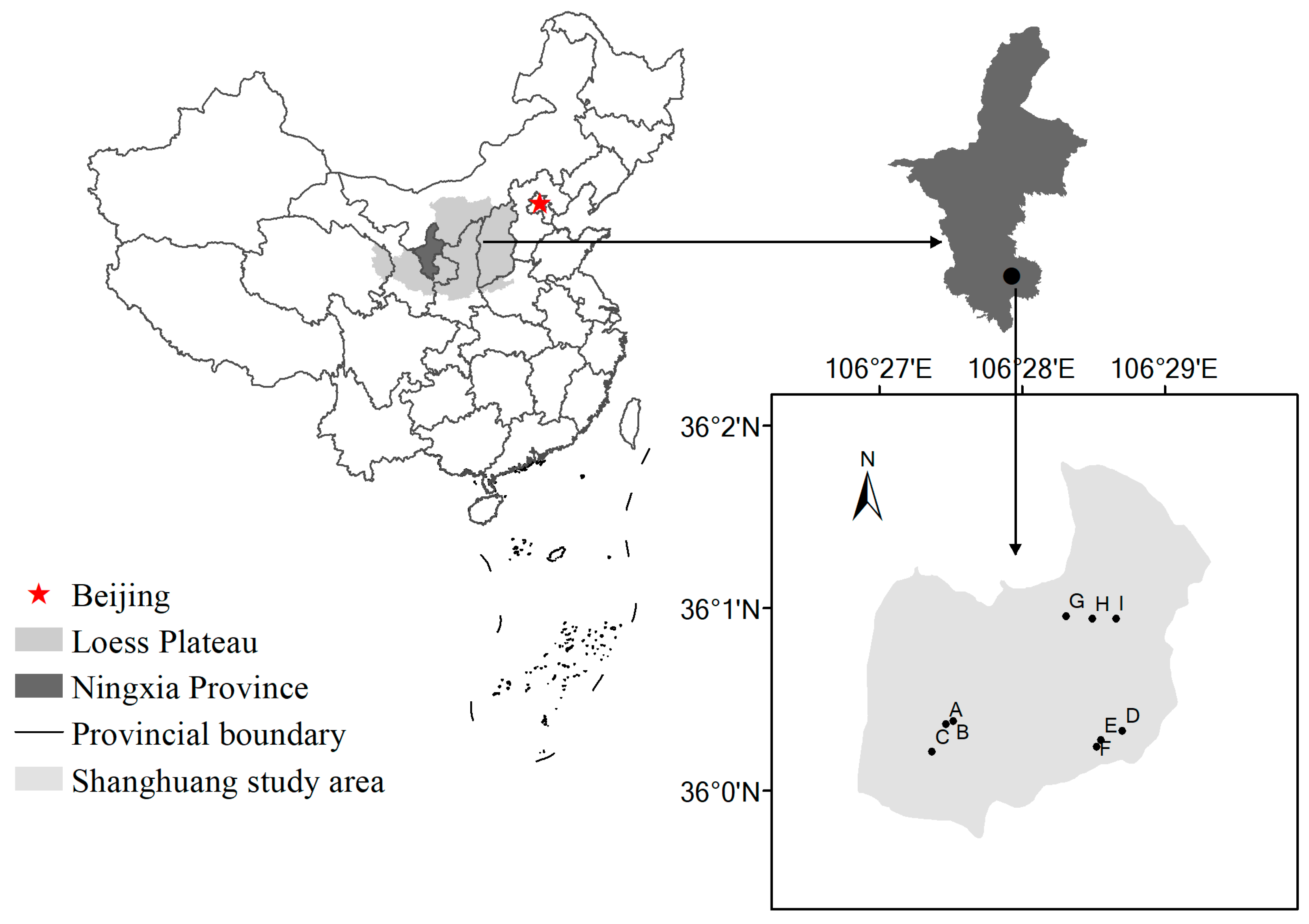
2.1. Study Site and Experimental Design
2.2. Soil Sampling and Preparation
2.3. Analysis of Soil Chemical Properties, Carbon Fractions, and Phospholipid Fatty Acid (PLFA) Profiles
2.4. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
3.2. Soil Carbon Fraction and Its Proportion
3.3. Soil Microbial Community Composition
3.4. Correlation of Soil Chemical Properties, Carbon Fractions, and Microbial Community
4. Discussion
4.1. Effect of Afforestation on Soil Carbon Sequestration
4.2. Effect of Afforestation on Soil Microbial Community
4.3. Changes in Soil Microbial Community Control Soil Carbon Sequestration
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grigal, D.F. Effect of extensive forest management on soil productivity. Forest Ecol. Manag. 2000, 138, 167–185. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Milori, D.M.B.P.; Martin-Neto, L.; Bayer, C.; Mielniczuk, J.; Bagnato, V.S. Humification degree of soil humic acids determined by fluorescence spectroscopy. Soil Sci. 2002, 167, 739–749. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Bailey, V.L.; Smith, J.L.; Bolton, H., Jr. Fungal-to-bacterial ratios in soils investigated for enhanced C sequestration. Soil Biol. Biochem. 2002, 34, 997–1007. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and Fungal Contributions to Carbon Sequestration in Agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Jia, G.M.; Liu, B.R.; Wang, G.; Zhang, B. The microbial biomass and activity in soil with shrub Caragana korshinskii K.) plantation in the semi-arid loess plateau in China. Eur. J. Soil Biol. 2010, 46, 6–10. [Google Scholar] [CrossRef]
- An, S.S.; Mentler, A.; Mayer, H.; Blum, W.E.H. Soil aggregation, aggregate stability, organic carbon and nitrogen in different soil aggregate fractions under forest and shrub vegetation on the Loess Plateau, China. Catena 2010, 81, 226–233. [Google Scholar] [CrossRef]
- Wei, X.R.; Shao, M.A.; Fu, X.L.; Horton, R. Changes in soil organic carbon and total nitrogen after 28 years grassland afforestation: Effects of tree species, slope position, and soil order. Plant Soil 2009, 331, 165–179. [Google Scholar] [CrossRef]
- Staff, S.S. Keys to Soil Taxonomy, 11th ed.; U.S. Department of Agriculture: Washington, DC, USA, 2010.
- Walkley, A.J.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Yan, C.S. The Soil Fertility Research Methods; China Agriciltural Press: Beijing, China, 1988. [Google Scholar]
- Sun, C.U.; Liu, J.S.; Wang, Y.; Zheng, N.; Wu, X.Q.; Liu, Q. Effect of long-term cultivation on soil organic carbon fractions and metal distribution in humic and fulvic acid in black soil, Northeast China. Soil Res. 2012, 50, 562–569. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, S.K. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Eco. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Deng, Q.; Cheng, X.; Yang, Y.; Zhang, Q.; Luo, Y. Carbon–nitrogen interactions during afforestation in central China. Soil Biol. Biochem. 2014, 69, 119–122. [Google Scholar] [CrossRef]
- Throop, H.; Archer, S.R. Spatial patterns of soil organic carbon relative to tree size and canopy distribution in a semidesert grassland. Glob. Chang. Biol. 2008, 14, 2420–2431. [Google Scholar] [CrossRef]
- Balesdent, J.C.C.; Balabane, M. Relationship of soil organic matter dynamics to physical protection and tillage. Soil Till. Res. 2000, 53, 215–230. [Google Scholar] [CrossRef]
- Qi, W.D.; Chen, Y.M.; Wang, L.L.; Zhang, F.; Zhang, X.W. Dynamics of soil organic carbon in Caragana microphylla forest and its relationship with environment factors in loess hilly region. Chin. Sci. Soil Water Conserv. 2011, 9, 72–77. [Google Scholar]
- Guimarães, D.V.; Gonzaga, M.I.S.; Da Silva, T.O.; Da Silva, T.L.; Da Silva Dias, N.; Matias, M.I.S. Soil organic matter pools and carbon fractions in soil under different land uses. Soil Till. Res. 2013, 126, 177–182. [Google Scholar] [CrossRef]
- Reddy, S.B.; Nagaraja, M.S.; Punish Raj, T.S.; Dhumgond, P.; Vignesh, N.S. Soil humic and fulvic acid fractions under different land use systems. Madras Agric. J. 2012, 99, 507–510. [Google Scholar]
- Stevenson, F.J. Humus Chemistry; John Wiley & Sons: New York, UY, USA, 1982. [Google Scholar]
- Cao, C.; Jiang, D.; Teng, X.; Jiang, Y.; Liang, W.; Cui, Z. Soil chemical and microbiological properties along a chronosequence of Caragana microphylla Lam. plantations in the Horqin sandy land of Northeast China. Appl. Soil Ecol. 2008, 40, 78–85. [Google Scholar] [CrossRef]
- Hortal, S.; Bastida, F.; Armas, C.; Lozano, Y.M.; Moreno, J.L.; García, C.; Pugnaire, F.I. Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol. Biochem. 2013, 64, 139–146. [Google Scholar] [CrossRef]
- Bossio, D.A.; Girvan, M.S.; Verchot, L.; Bullimore, J.; Borelli, T.; Albrecht, A.; Scow, K.M.; Ball, A.S.; Pretty, J.N.; Osborn, A.M. Soil microbial community response to land use change in an agricultural landscape of western Kenya. Microb. Ecol. 2005, 49, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.; Chen, C.; Xu, Z.; Ghadiri, H. Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. J. Soil Sediment. 2010, 10, 1267–1277. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Thomas, N.; Robinson, L.; Tate, K.R.; Ross, D.J.; Dando, J.; Singh, B.K. Physiological, biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata. Soil Biol. Biochem. 2009, 41, 1642–1651. [Google Scholar] [CrossRef]
- Yan, H.; Huang, Y.M.; Jiang, Y.L.; Zhao, T. Soil mineralization under two kinds of shrub lands in mountainous area of sourthern Ningxia, Northwest China. Acta Sci. Cirumstantiae 2014, 34, 2111–2120. [Google Scholar]
- Nsabimana, D.; Haynes, R.J.; Wallis, F.M. Size, activity and catabolic diversity of the soil microbial biomass as affected by land use. Appl. Soil Ecol. 2004, 26, 81–92. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim. Chang. 2006, 80, 5–23. [Google Scholar] [CrossRef]


{kind=link}
| Vegetation Type | Depth (cm) | SOC | Total·N | NH4-N | NO3-N | Available p | C/N | Bulk Density |
|---|---|---|---|---|---|---|---|---|
| g·kg−1 | g·kg−1 | mg·kg−1 | mg·kg−1 | mg·kg−1 | g·cm−3 | |||
| Cropland | 0–20 | 7.66 ± 0.10Ca | 0.89 ± 0.19Ba | 3.75 ± 0.50Ba | 13.10 ± 0.05Aa | 9.37 ± 0.54Aa | 8.61 ± 0.29Aa | 1.30 |
| 20–40 | 6.35 ± 0.28Cb | 0.67 ± 0.02Ca | 3.69 ± 0.26Ca | 8.08 ± 0.38Ab | 2.55 ± 0.13Bb | 9.48 ± 0.57Aa | 1.51 | |
| shrubland I | 0–20 | 12.39 ± 0.15Aa | 1.45 ± 0.06Aa | 6.02 ± 0.12Aa | 7.46 ± 0.18Ba | 4.89 ± 0.21Ba | 8.55 ± 0.27Aa | 1.04 |
| 20–40 | 7.43 ± 0.18 Bb | 0.92 ± 0.07Bb | 5.71 ± 0.03Aa | 6.41 ± 0.20Bb | 3.57 ± 0.20Ab | 8.14 ± 0.84ABa | 1.14 | |
| shrubland II | 0–20 | 10.38 ± 0.37Ba | 1.59 ± 0.07Aa | 5.07 ± 0.23Ba | 7.67 ± 0.20Ba | 4.24 ± 0.13Ba | 6.52 ± 0.37Ba | 1.17 |
| 20–40 | 8.80 ± 0.02Ab | 1.37 ± 0.09Ab | 4.37 ± 0.06Ba | 5.96 ± 0.19Bb | 3.31 ± 0.19Ab | 6.43 ± 0.25Ba | 1.30 |
| Vegetation Type | Depth (cm) | FA-C (Fulvic Acid) | HA-C (Humic Acid) | HA-C/FA-C | ||
|---|---|---|---|---|---|---|
| Concentration (g·kg−1) | Proportion to Total SOC (%) | Concentration (g·kg−1) | Proportion to Total SOC (%) | |||
| Cropland | 0–20 | 0.76 ± 0.13Ba | 9.9C | 0.48 ± 0.01Aa | 6.3A | 0.63 ± 0.09Aa |
| 20–40 | 0.63 ± 0.13Ba | 10.0B | 0.28 ± 0.01Bb | 4.4A | 0.45 ± 0.10Aa | |
| shrubland I | 0–20 | 1.64 ± 0.06Aa | 13.2B | 0.44 ± 0.01Aa | 3.5B | 0.27 ± 0.01Ca |
| 20–40 | 0.78 ± 0.02Bb | 10.4B | 0.13 ± 0.01Cb | 1.7B | 0.17 ± 0.02Ca | |
| shrubland II | 0–20 | 1.61 ± 0.05Aa | 15.6A | 0.54 ± 0.01Aa | 5.2A | 0.34 ± 0.08Ba |
| 20–40 | 1.17 ± 0.03Ab | 13.3A | 0.41 ± 0.03Ab | 4.7A | 0.35 ± 0.01Ba | |
| Vegetation Type | Depth (cm) | Total PLFAs | G+ Bacterial PLFAs a | G− Bacterial PLFAs b | Bacterial PLFAs c | Fungal PLFA d | Actinomycetic PLFAs e | G+:G− Bacterial PLFAs | Fungal:Bacterial PLFAs |
|---|---|---|---|---|---|---|---|---|---|
| Cropland | 0–20 | 29.17 ± 0.99Ca | 4.79 ± 0.34Ca | 7.42 ± 0.21Ca | 20.30 ± 0.89Ca | 3.83 ± 0.12Ba | 1.47 ± 0.03Ca | 0.64 ± 0.03Ab | 2.61 ± 0.14Ab |
| 20–40 | 10.89 ± 0.48Cb | 1.53 ± 0.16Cb | 1.40 ± 0.09Cb | 7.12 ± 0.23Cb | 2.20 ± 0.11Ab | 0.42 ± 0.01Cb | 1.09 ± 0.05Aa | 5.25 ± 0.16Aa | |
| shrubland I | 0–20 | 35.98 ± 0.38Ba | 6.55 ± 0.34Ba | 9.23 ± 0.35Ba | 24.95 ± 0.49Ba | 3.50 ± 0.35Ba | 2.50 ± 0.05Ba | 0.71 ± 0.01Ab | 1.41 ± 0.17Ba |
| 20–40 | 19.04 ± 0.50Bb | 3.93 ± 0.03Bb | 3.89 ± 0.25Bb | 13.31 ± 0.21Bb | 1.78 ± 0.20Bb | 1.44 ± 0.08Bb | 1.01 ± 0.07Aa | 1.24 ± 0.21Ba | |
| shrubland II | 0–20 | 51.02 ± 1.57Aa | 9.33 ± 0.59Aa | 14.18 ± 0.06Aa | 36.14 ± 0.99Aa | 4.54 ± 0.11Aa | 3.19 ± 0.34Aa | 0.66 ± 0.04Ab | 1.43 ± 0.12Ba |
| 20–40 | 28.62 ± 1.03Ab | 6.48 ± 0.40Ab | 5.77 ± 0.36Ab | 19.86 ± 0.78Ab | 2.53 ± 0.19Ab | 2.22 ± 0.19Aa | 1.12 ± 0.02Aa | 1.15 ± 0.18Ba |
| SOC | FA-C | HA-C | HA-C/FA-C | |
|---|---|---|---|---|
| Soil Chemical and Physical Properties | ||||
| TN | 0.869 * | 0.957 ** | 0.622 | –0.376 |
| C/N | –0.358 | –0.554 | –0.414 | 0.270 |
| NO3-N | –0.222 | –0.325 | 0.396 | 0.866 * |
| NH4-N | 0.682 | 0.613 | –0.177 | –0.853 * |
| Available P | 0.054 | –0.082 | 0.470 | 0.671 |
| Bulk density | –0.758 | –0.855 * | –0.610 | 0.309 |
| Soil Microbial Community | ||||
| Total PLFAs | 0.766 | 0.861 * | 0.790 | –0.074 |
| G+ Bacterial | 0.763 | 0.875 * | 0.711 | –0.206 |
| G− Bacterial | 0.747 | 0.831 * | 0.782 | –0.041 |
| Bacterial | 0.758 | 0.855 * | 0.783 | –0.078 |
| Fungi | 0.587 | 0.636 | 0.916 * | 0.362 |
| Actinomycetes | 0.832 * | 0.921 ** | 0.643 | –0.321 |
| G+/G− | –0.552 | –0.490 | –0.679 | –0.292 |
| F/B | –0.604 | –0.607 | –0.183 | 0.522 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, Y.; Cheng, M.; Huang, Y.; An, S.; Darboux, F. Changes in Soil Microbial Community and Its Effect on Carbon Sequestration Following Afforestation on the Loess Plateau, China. Int. J. Environ. Res. Public Health 2017, 14, 948. https://doi.org/10.3390/ijerph14080948
Xiang Y, Cheng M, Huang Y, An S, Darboux F. Changes in Soil Microbial Community and Its Effect on Carbon Sequestration Following Afforestation on the Loess Plateau, China. International Journal of Environmental Research and Public Health. 2017; 14(8):948. https://doi.org/10.3390/ijerph14080948
Chicago/Turabian StyleXiang, Yun, Man Cheng, Yimei Huang, Shaoshan An, and Frédéric Darboux. 2017. "Changes in Soil Microbial Community and Its Effect on Carbon Sequestration Following Afforestation on the Loess Plateau, China" International Journal of Environmental Research and Public Health 14, no. 8: 948. https://doi.org/10.3390/ijerph14080948
APA StyleXiang, Y., Cheng, M., Huang, Y., An, S., & Darboux, F. (2017). Changes in Soil Microbial Community and Its Effect on Carbon Sequestration Following Afforestation on the Loess Plateau, China. International Journal of Environmental Research and Public Health, 14(8), 948. https://doi.org/10.3390/ijerph14080948