Assessment of the Public Health Threats Posed by Vector-Borne Disease in the United Kingdom (UK)

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Mosquitoes & Mosquito-Borne Disease
2.1. Chikungunya, Dengue and Zika Viruses and Invasive Aedes Mosquitoes
2.1.1. Importance of Aedes aegypti
2.1.2. Aedes aegypti in Europe
2.1.3. Aedes aegypti in the UK—Past, Present, and Future
2.1.4. Aedes albopictus in Europe
2.1.5. Aedes albopictus—Risks for the UK
2.2. West Nile Virus and Culex Mosquitoes
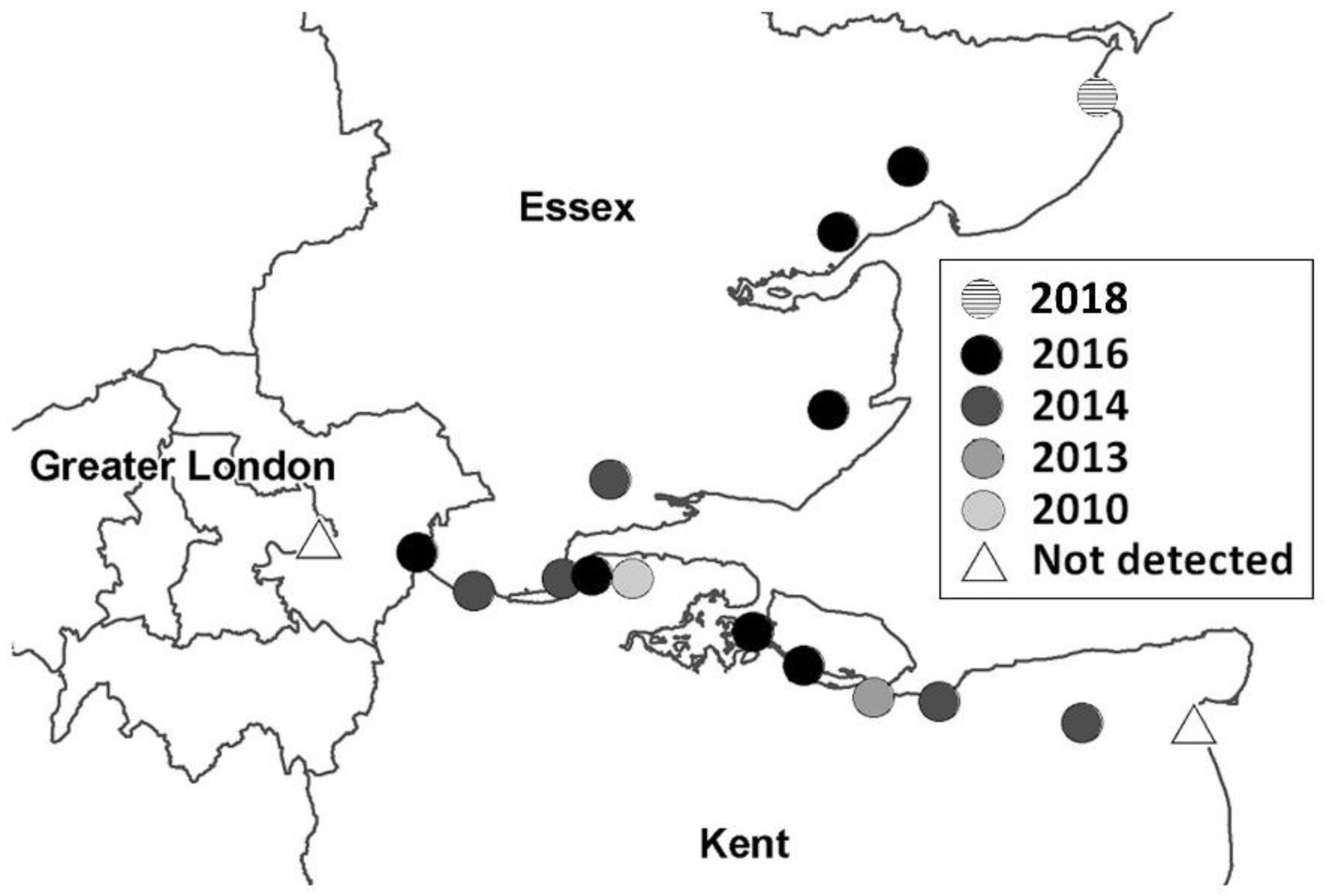
2.2.1. The Detection and Expansion of Culex modestus in the UK
2.2.2. Environmental Change and British Mosquitoes
2.3. Other Mosquito-Borne Infections of Potential UK Concern
2.3.1. Malaria Risk for the UK
2.3.2. Other Arboviruses
2.3.3. Dirofilaria
3. Ticks & Tick-Borne Diseases
3.1. Ixodes ricinus and Lyme borreliosis
3.1.1. Expansion of Ixodes ricinus Ticks
3.1.2. Urban Tick Issues
3.1.3. Borrelia Prevalence in Ticks & Impacts of Environmental Change & Wildlife Management
3.2. Ixodes ricinus and Other Known or Potential Pathogens
3.2.1. Anaplasma, Borrelia miyamotoi & Rickettsia
3.2.2. Tick-Borne Encephalitis and Congo-Crimean Haemorrhagic Fever Viruses
3.3. Dermacentor reticulatus and Associated Infections
3.4. Haemaphysalis punctata and Potential Disease Issues
3.5. Imported Ticks: Rhipicephalus sanguineus s.l.
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Napp, S.; Petrić, D.; Busquets, N. West Nile virus and other mosquito-borne viruses present in Eastern Europe. Pathog. Glob. Health 2018, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Venturi, G.; Di Luca, M.; Fortuna, C.; Remoli, M.E.; Riccardo, F.; Severini, F.; Toma, L.; Del Manso, M.; Benedetti, E.; Caporali, M.G.; et al. Detection of chikungunya outbreak in Central Italy, August to September 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, J.; Recker, M. The 2012 Madeira Dengue Outbreak: Epidemiological Determinants and Future Epidemic Potential. PLoS Negl. Trop. Dis. 2014, 8, e3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andriopoulos, P.; Economopoulou, A.; Spanakos, G.; Assimakopoulos, G. A local outbreak of autochthonous Plasmodium vivax malaria in Laconia, Greece—A re-emerging infection in the southern borders of Europe? Int. J. Infect. Dis. 2013, 17, e125–e128. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Hauffe, H.; Carpi, G.; Vourc, H.G.; Neteler, M.; Rosa, R. Lyme borreliosis in Europe. Eurosurveillance 2011, 16, 19906. [Google Scholar] [PubMed]
- Jaenson, T.G.; Hjertqvist, M.; Bergström, T.; Lundkvist, A. Why is tick-borne encephalitis increasing? A review of the key factors causing the increasing incidence of human TBE in Sweden. Parasites Vectors 2012, 5, 184. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.J.; Mellor, P.S. Bluetongue in Europe: Past, present and future. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2669–2681. [Google Scholar] [CrossRef] [PubMed]
- Afonso, A.; Abrahantes, J.C.; Conraths, F.; Veldhuis, A.; Elbers, A.; Roberts, H.; Van der Stede, Y.; Méroc, E.; Gache, K.; Richardson, J. The Schmallenberg virus epidemic in Europe-2011-2013. Prev. Vet. Med. 2014, 116, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Fonville, M.; Jahfari, S.; Sprong, H.; Medlock, J.M. Borrelia miyamotoi in host-seeking Ixodes ricinus ticks in England. Epidemiol. Infect. 2015, 143, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Tijsse-Klasen, E.; Jameson, L.J.; Fonville, M.; Leach, S.; Sprong, H.; Medlock, J.M. First detection of spotted fever group rickettsiae in Ixodes ricinus and Dermacentor reticulatus ticks in the UK. Epidemiol. Infect. 2011, 139, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsky, H.C.; Matranga, C.B.; Wohl, S.; Schaffner, S.F.; Freije, C.A.; Winnicki, S.M.; West, K.; Qu, J.; Baniecki, M.L.; Gladden-Young, A.; et al. Zika virus evolution and spread in the Americas. Nature 2017, 546, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.F.; Choi, G.K.; Yip, C.C.; Cheng, V.C.; Yuen, K.Y. Zika fever and congenital Zika syndrome: An unexpected emerging arboviral disease. J. Infect. 2016, 72, 507–524. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Avenell, D.; Barrass, I.; Leach, S. Analysis of the potential for survival and seasonal activity of Aedes albopictus (Diptera: Culicidae) in the United Kingdom. J. Vector Ecol. 2006, 31, 292–304. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauchemez, S.; Ledrans, M.; Poletto, C.; Quenel, P.; de Valk, H.; Colizza, V.; Boëlle, P.Y. Local and regional spread of chikungunya fever in the Americas. Eurosurveillance 2014, 19, 20854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- CARPHA 2017. Available online: http://carpha.org/articles/ID/167/Integrated-Vector-Management-a-Critical-Strategy-for-Fighting-Vector-Borne-Diseases-in-the-Region (accessed on 26 September 2018).
- Benelli, G.; Mehlhorn, H. Declining malaria, rising of dengue and Zika virus: Insights for mosquito control. Para. Res. 2016, 115, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.B.; Beier, J.C.; Benelli, G. Transgenic mosquitoes—Fact or fiction? Trends Parasitol. 2018, 34, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F.; Mathis, A. Dengue and dengue vectors in the WHO European region: Past, present, and scenarios for the future. Lancet Infect. Dis. 2014, 14, 1271–1280. [Google Scholar] [CrossRef]
- Rosen, L. Dengue in Greece in 1927 and 1928 and the pathogenesis of dengue hemorrhaguc fever: New data and a different conclusion. Am. J. Trop. Med. Hyg. 1986, 35, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Akiner, M.A.; Demirci, B.; Babuadze, G.; Robert, V.; Schaffner, F. Spread of the Invasive Mosquitoes Aedes aegypti and Aedes albopictus in the Black Sea Region Increases Risk of Chikungunya, Dengue, and Zika Outbreaks in Europe. PLoS. Negl. Trop. Dis. 2016, 10, e0004664. [Google Scholar] [CrossRef]
- Meers, P.D. Yellow fever in Swansea, 1865. J. Hyg. 1986, 97, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Dallimore, T.; Hunter, A.; Medlock, J.M.; Vaux, A.G.C.; Harbach, R.E.; Strode, C. Discovery of a Single Male Aedes aegypti (L.) in Merseyside, England. Parasites Vectors 2017, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Ibañez-Justicia, A.; Gloria-Soria, A.; den Hartog, W.; Dik, M.; Jacobs, F.; Stroo, A. The first detected airline introductions of yellow fever mosquitoes (Aedes aegypti) to Europe, at Schiphol International airport, the Netherlands. Parasit Vectors 2017, 10, 603. [Google Scholar] [CrossRef] [PubMed]
- Vaux, A.G.C.; Dallimore, T.; Cull, B.; Schaffner, F.; Strode, C.; Pflueger, V.; Murchie, A.; Newham, Z.; Catton, M.; Medlock, J.M. The challenge of invasive mosquito vectors in the United Kingdom 2016–2017: A summary of surveillance and control efforts. in press.
- Schaffner, F.; Medlock, J.M.; Van Bortel, W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. 2013, 19, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Romi, R.; Di Luca, M.; Majori, G. Current status of Aedes albopictus and Aedes atropalpus in Italy. J. Am. Mosquito Control Assoc. 1999, 15, 425–427. [Google Scholar]
- Osório, H.C.; Zé-Zé, L.; Neto, M.; Silva, S.; Marques, F.; Silva, A.S.; Alves, M.J. Detection of the Invasive Mosquito Species Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Portugal. Int. J. Environ. Res. Public Health 2018, 15, 820. [Google Scholar] [CrossRef] [PubMed]
- ECDC 2018. Aedes albopictus Maps. Available online: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps. (accessed on 26 September 2018).
- Romi, R.; Majori, G. An overview of the lesson learned in almost 20 years of fight against the “tiger” mosquito. Parassitologia 2008, 50, 117–119. [Google Scholar] [PubMed]
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes albopictus as an epidemic vector of chikungunya virus: Another emerging problem? Lancet Infect. Dis. 2006, 6, 463–464. [Google Scholar] [CrossRef]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Calba, C.; Guerbois-Galla, M.; Franke, F.; Jeannin, C.; Auzet-Caillaud, M.; Grard, G.; Pigaglio, L.; Decoppet, A.; Weicherding, J.; Savaill, M.C.; et al. Preliminary report of an autochthonous chikungunya outbreak in France, July to September 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ECDC 2009. Development of Aedes albopictus Risk Maps. Available online: https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/0905_TER_Development_of_Aedes_Albopictus_Risk_Maps.pdf (accessed on 26 September 2018).
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface 2012, 9, 2708–2717. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Leach, S.A. Effect of climate change on vector-borne disease risk in the UK. Lancet Infect Dis. 2015, 15, 721–730. [Google Scholar] [CrossRef]
- Murphy, G.; Vaux, A.; Medlock, J. Challenges in undertaking mosquito surveillance at UK seaports and airports to prevent the entry and establishment of invasive vector species. Int. J. Environ. Health Res. 2013, 23, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Vaux, A.G.; Medlock, J.M. Current status of invasive mosquito surveillance in the UK. Parasites Vectors 2015, 8, 351. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.; Cull, B.; Schaffner, F.; Gillingham, E.; Pfluger, V.; Leach, S. Detection of the invasive mosquito species Aedes albopictus in southern England. Lancet Infect Dis. 2017, 17, 140. [Google Scholar] [CrossRef]
- Murray, K.O.; Mertens, E.; Despres, P. West Nile virus and its emergence in the United States of America. Vet. Res. 2010, 41, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, J.M.; Snow, K.R.; Leach, S. Potential transmission of West Nile virus in the British Isles: An ecological review of candidate mosquito bridge vectors. Med. Vet. Entomol. 2005, 19, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Vazeille, M.; Reiter, P.; Schaffner, F.; Zeller, H.; Bicout, D.J. Evidence of laboratory vector competence of Culex modestus for West Nile virus. J. Am. Mosq. Control. Assoc. 2007, 23, 233–236. [Google Scholar] [CrossRef]
- Medlock, J.M.; Vaux, A.G.C. Aedes (Aedes) geminus Peus (Diptera, Culicidae)—An addition to the British mosquito fauna. Dipter. Dig. 2009, 16, 1–4. [Google Scholar]
- Golding, N.; Nunn, M.A.; Medlock, J.M.; Purse, B.V.; Vaux, A.G.; Schäfer, S.M. West Nile virus vector Culex modestus established in southern England. Parasites Vectors 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbach, R.E.; Dallimore, T.; Briscoe, A.G.; Culverwell, C.L.; Vaux, A.G.C.; Medlock, J.M. Aedes nigrinus (Eckstein, 1918) (Diptera, Culicidae), a new country record for England, contrasted with Aedes sticticus (Meigen, 1838). Zookeys 2017, 119–130. [Google Scholar] [CrossRef]
- Hutchinson, R.A.; West, P.A.; Lindsay, S.W. Suitability of two carbon dioxide-baited traps for mosquito surveillance in the United Kingdom. Bull. Entomol. Res. 2007, 97, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G. Distribution of West Nile virus vector, Culex modestus, in England. Vet. Rec. 2012, 171, 278. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.; Gibson, G.; Hawkes, F.M.; Cheke, R.A. Potential vector for West Nile virus prevalent in Kent. Vet. Rec. 2014, 175, 284–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaux, A.G.C.; Gibson, G.; Hernandez-Triana, L.M.; Cheke, R.A.; McCracken, F.; Jeffries, C.L.; Horton, D.L.; Springate, S.; Johnson, N.; Fooks, A.R.; et al. Enhanced West Nile virus surveillance in the North Kent marshes, UK. Parasites Vectors 2015, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Cull, B.; Vaux, A.G.; Medlock, J.M.; Abbott, A.; Gibson, G. Expansion of the range of the West Nile virus vector in Essex. Vet. Rec. 2016, 179, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G. Colonization of UK coastal realignment sites by mosquitoes: Implications for design, management, and public health. J. Vector Ecol. 2013, 38, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Brugman, V.A.; England, M.E.; Stoner, J.; Tugwell, L.; Harrup, L.E.; Wilson, A.J.; Medlock, J.M.; Logan, J.G.; Fooks, A.R.; Mertens, P.P.C.; et al. How often do mosquitoes bite humans in southern England? A standardised summer trial at four sites reveals spatial, temporal and site-related variation in biting rates. Parasites Vectors 2017, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Brugman, V.A.; Hernández-Triana, L.M.; England, M.E.; Medlock, J.M.; Mertens, P.P.C.; Logan, J.G.; Wilson, A.J.; Fooks, A.R.; Johnson, N.; Carpenter, S. Blood-feeding patterns of native mosquitoes and insights into their potential role as pathogen vectors in the Thames estuary region of the United Kingdom. Parasites Vectors 2017, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phipps, L.P.; Duff, J.P.; Holmes, J.P.; Gough, R.E.; McCracken, F.; McElhinney, L.M.; Johnson, N.; Hughes, L.; Chantrey, J.; Pennycott, T.; et al. Surveillance for West Nile virus in British birds (2001 to 2006). Vet. Rec. 2008, 162, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Brugman, V.A.; Horton, D.L.; Phipps, L.P.; Johnson, N.; Cook, A.J.; Fooks, A.R.; Breed, A.C. Epidemiological perspectives on West Nile virus surveillance in wild birds in Great Britain. Epidemiol. Infect. 2013, 141, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; Dawson, A.; Gould, E.A. Detection of seroconversion to West Nile virus, Usutu virus and Sindbis virus in UK sentinel chickens. Virol. J. 2006, 3, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, J.M.; Vaux, A.G. Colonization of a newly constructed urban wetland by mosquitoes in England: Implications for nuisance and vector species. J. Vector Ecol. 2014, 39, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G. Seasonal dynamics and habitat specificity of mosquitoes in an English wetland: Implications for UK wetland management and restoration. J. Vector Ecol. 2015, 40, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.C. Impacts of the creation, expansion and management of English wetlands on mosquito presence and abundance—Developing strategies for future disease mitigation. Parasites Vectors 2015, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Blagrove, M.S.; Sherlock, K.; Chapman, G.E.; Impoinvil, D.E.; McCall, P.J.; Medlock, J.M.; Lycett, G.; Soloman, S.; Baylis, M. Evaluation of the vector competence of a native UK mosquito Ochlerotatus detritus (Aedes detritus) for dengue, chikungunya and West Nile viruses. Parasites Vectors 2016, 9, 452. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, S.W.; Hole, D.G.; Hutchinson, R.A.; Richards, S.A.; Willis, S.G. Assessing the future threat from vivax malaria in the United Kingdom using two markedly different modelling approaches. Malar. J. 2010, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caminade, C.; Kovats, S.; Rocklov, J.; Topkins, A.M.; Morse, A.P.; Colon-Gonzalez, F.J.; Stenlund, H.; Martens, P.; Lloyd, S.J. Impact of climate change on global malaria distribution. Proc. Natl. Acad. Sci. USA 2014, 111, 3286–3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, S.W.; Thomas, C.J. Global warming and risk of vivax malaria in Great Britain. Glob. Chang. Hum. Health 2001, 2, 80–84. [Google Scholar] [CrossRef]
- Deboninck, W.; Hendrickx, F.; Van Nortel, W.; Versteirt, V.; Coosemans, M.; Damiens, D.; Hance, T.; De Clercq, E.M.; Hendrickx, G.; Schaffner, F.; et al. 2011 Human-induced expanded distribution of Anopheles plumbeus, experimental vector of West Nile virus and a potential vector of human malaria in Belgium. J. Med. Entomol. 2011, 48, 924–928. [Google Scholar] [CrossRef]
- Medlock, J.M.; Snow, K.R.; Leach, S. Possible ecology and epidemiology of medically important mosquito-borne arboviruses in Great Britain. Epidemiol. Infect. 2007, 135, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; Lawson, B.; Egbetade, A.; Jeffries, C.; Johnson, N.; Cunningham, A.A.; Fooks, A.R. Targeted surveillance for Usutu virus in British birds (2005–2011). Vet. Rec. 2013, 172, 17. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Triana, L.M.; de Marco, M.F.; Mansfield, K.L.; Thorne, L.; Lumley, S.; Marston, D.; Fooks, A.A.; Johnson, N. Assessment of vector competence of UK mosquitoes for Usutu virus of African origin. Parasit Vectors 2018, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Cull, B.; Vaux, A.G.C.; Irwin, A.G. The mosquito Aedes vexans in England. Vet. Rec. 2017, 181, 243. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.; Hernández-Triana, L.M.; Horton, D.L.; Fernández de Marco, M.D.M.; Medlock, J.M.; Hewson, R.; Fooks, A.R.; Johnson, N. Competence of mosquitoes native to the United Kingdom to support replication and transmission of Rift Valley fever virus. Parasites Vectors 2018, 11, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancrini, G.; Scaramozzino, P.; Gabrielli, S.; Di Paolo, M.; Toma, L.; Romi, R. Aedes albopictus and Culex pipiens implicated as natural vectors of Dirofilaria repens in central Italy. J. Med. Entomol. 2007, 44, 1064–1066. [Google Scholar] [CrossRef]
- Medlock, J.M.; Barrass, I.; Kerrod, E.; Taylor, M.A.; Leach, S. Analysis of climatic predictions for extrinsic incubation of Dirofilaria in the United kingdom. Vector Borne Zoonotic Dis. 2007, 7, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, P.D. Ticks of North-West Europe; Synopses of the British Fauna (New, Series); Barnes, R.S.K., Crothers, J.H., Eds.; CABI: Wallingford, UK, 1996; p. 178. [Google Scholar]
- Medlock, J.M.; Jameson, L.J.; Pietzsch, M.E.; Gilbert, L. British Mosquitoes. Br. Wildl. 2009, 20, 258–266. [Google Scholar]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, L.J.; Medlock, J.M. Tick surveillance in Great Britain. Vector Borne Zoonotic Dis. 2011, 11, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Cull, B.; Pietzsch, M.E.; Hansford, K.M.; Gillingham, E.L.; Medlock, J.M. Surveillance of British ticks: An overview of species records, host associations, and new records of Ixodes ricinus distribution. Ticks Tick Borne Dis. 2018, 9, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Vaux, A.G.C.; Cull, B.; Abdullah, S.; Pietzsch, M.E.; Wall, R.; Johnson, N.; Phipps, L.P. Distribution of the tick Dermacentor reticulatus in the United Kingdom. Med. Vet. Entomol. 2017, 31, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, J.; Vaux, A.G.C.; Cull, B.; Pietzsch, M.E.; Gillingham, E.; Johnson, N.; Phipps, L.P. Has the red sheep tick, Haemaphysalis punctata, recently expanded its range in England? Med. Vet. Entomol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Martyn, K.P. Provisional Atlas of the Ticks (Ixodoidea) of the British Isles; Biological Records Centre: Cumbria, UK, 1988. [Google Scholar]
- Craine, N.G.; Randolph, S.E.; Nuttall, P.A. Seasonal variation in the role of grey squirrels as hosts of Ixodes ricinus, the tick vector of the Lyme disease spirochaete, in a British woodland. Folia Parasitol. 1995, 42, 73–80. [Google Scholar] [PubMed]
- Craine, N.G.; Nuttall, P.A.; Marriott, A.C.; Randolph, S.E. Role of grey squirrels and pheasants in the transmission of Borrelia burgdorferi sensu lato, the Lyme disease spirochaete, in the UK. Folia Parasitol. 1997, 44, 155–160. [Google Scholar] [PubMed]
- Hoodless, A.N.; Kurtenbach, K.; Peacey, M.; Nuttall, P.A.; Randolph, S.E. The role of pheasants (Phasianus colchicus) as hosts for ticks (Ixodes ricinus) and Lyme disease spirochaetes (Borrelia burgdorferi) in southern England. Game Wildl. 1998, 15, 477–489. [Google Scholar]
- Kurtenbach, K.; Kampen, H.; Dizij, A.; Arndt, S.; Seitz, H.M.; Schaible, U.E.; Simon, M.M. Infestation of rodents with larval Ixodes ricinus (Acari: Ixodidae) is an important factor in the transmission cycle of Borrelia burgdorferi s.l. in German woodlands. J. Med. Entomol. 1995, 32, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; Carey, D.; Hoodless, A.N.; Nuttall, P.A.; Randolph, S.E. Competence of pheasants as reservoirs for Lyme disease spirochetes. J. Med. Entomol. 1998, 35, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; Peacey, M.; Rijpkema, S.G.T.; Hoodless, A.N.; Nuttall, P.A.; Randolph, S.E. Differential transmission of the genospecies of Borrelia burgdorferi sensu lato by game birds and small rodents in England. Appl. Environ. Microbiol. 1998, 64, 1169–1174. [Google Scholar] [PubMed]
- Millins, C.; Magierecka, A.; Gilbert, L.; Edoff, A.; Brereton, A.; Kilbride, E.; Denwood, M.; Birtles, R.; Biek, R. An invasive mammal (Gray squirrel, Sciurus carolinensis) commonly hosts diverse and atypical genotypes of the zoonotic pathogen Borrelia burgdorferi senu lato. Appl. Environ. Microbiol. 2015, 81, 4236–4245. [Google Scholar] [CrossRef] [PubMed]
- Pietzsch, M.E.; Medlock, J.M.; Jones, L.; Avenell, D.; Abbott, J.; Harding, P.; Leach, S. Distribution of Ixodes ricinus in the British Isles: Investigation of historical records. Med. Vet. Entomol. 2005, 19, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Public Health England [PHE] LYME Borreliosis Epidemiology and Surveillance. 2018. Available online: https://www.gov.uk/government/publications/lyme-borreliosis-epidemiology/lyme-borreliosis-epidemiology-and-surveillance (accessed on 1 June 2018).
- Hansford, K.M.; Fonville, M.; Gillingham, E.L.; Coipan, E.C.; Pietzsch, M.E.; Krawczyk, A.I.; Vaux, A.G.C.; Cull, B.; Sprong, H.; Medlock, J.M. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick Borne Dis. 2017, 8, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Leonardi, G.S.; White, M.P.; Medlock, J.; Alcock, I.; Macintyre, H.L.; Maguire, K.; Nichols, G.; Wheeler, B.W.; Morris, G.; et al. Beyond climate change and health: Integrating broader environmental change and natural environments for public health protection and promotion in the UK. Atmposhere 2018, 9, 245. [Google Scholar] [CrossRef]
- Millins, C.; Gilbert, L.; Medlock, J.; Hansford, K.; Thompson, D.B.; Biek, R. Effects of conservation management of landscapes and vertebrate communities on Lyme borreliosis risk in the United Kingdom. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, J.M.; Shuttleworth, H.; Copley, V.; Hansford, K.M.; Leach, S. Woodland biodiversity management as a tool for reducing human exposure to Ixodes ricinus ticks: A preliminary study in an English woodland. J. Vector Ecol. 2012, 37, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Sprong, H.; Jansen, P.A.; Prins, H.H.T.; van Wieren, S.E. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in Dutch forests. Parasit Vectors 2017, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Bettridge, J.; Renard, M.; Zhao, F.; Bown, K.J.; Birtles, R.J. Distribution of Borrelia burgdorferi sensu lato in Ixodes ricinus populations across central Britain. Vector Borne Zoonotic Dis. 2013, 13, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S. The ecology of ticks transmitting Lyme borreliosis. Exp. Appl. Acarol. 1998, 22, 249–258. [Google Scholar] [CrossRef]
- Smith, F.D.; Wall, L.E. Prevalence of Babesia and Anaplasma in ticks infesting dogs in Great Britain. Vet. Parasitol. 2013, 198, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L. Louping ill virus in the UK: A review of the hosts, transmission and ecological consequences of control. Exp. Appl. Acarol. 2016, 68, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Bown, K.J.; Begon, M.; Bennett, M.; Birtles, R.J.; Burthe, S.; Lambin, X.; Telfer, S.; Woldehiwet, Z.; Ogden, N.H. Sympatric Ixodes trianguliceps and Ixodes ricinus ticks feeding on field voles (Microtus agrestis): Potential for increased risk of Anaplasma phagocytophilum in the United Kingdom? Vector Borne Zoonotic Dis. 2006, 6, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Stuen, S. Anaplasma phagocytophilum—The most widespread tick-borne infection in animals in Europe. Vet. Res. Commun. 2007, 31, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Tijsse-Klasen, E.; Hansford, K.M.; Jahfari, S.; Phipps, P.; Sprong, H.; Medlock, J.M. Spotted fever group rickettsiae in Dermacentor reticulatus and Haemaphysalis punctata ticks in the UK. Parasites Vectors 2013, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; de Vries, A.; Rijks, J.M.; Van Gucht, S.; Vennema, H.; Sprong, H.; Rockx, B. Tick-borne encephalitis virus in ticks and roe deer, the Netherlands. Emerg. Infect. Dis. 2017, 23, 1028–1030. [Google Scholar] [CrossRef] [PubMed]
- Pietzsch, M.E.; Mitchell, R.; Jameson, L.J.; Morgan, C.; Medlock, J.M.; Collins, D.; Chamberlain, J.C.; Gould, E.A.; Hewson, R.; Taylor, M.A.; et al. Preliminary evaluation of exotic tick species and exotic pathogens imported on migratory birds into the British Isles. Vet. Parasitol. 2008, 155, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Jameson, L.J.; Morgan, P.J.; Medlock, J.M.; Watola, G.; Vaux, A.G. Importation of Hyalomma marginatum, vector of Crimean-Congo haemorrhagic fever virus, into the United Kingdom by migratory birds. Ticks Tick Borne Dis. 2012, 3, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Jameson, L.J.; Phipps, L.P. Status of Dermacentor reticulatus in the UK. Vet. Rec. 2011, 168, 386–387. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Medlock, J.M.; Swainsbury, C.; Phipps, L.P.; De Marco Mdel, M.; Hernández-Triana, L.M.; Johnson, N.; Fooks, A.R. Babesia canis infection in ticks in Essex. Vet. Rec. 2016, 178, 323. [Google Scholar] [CrossRef] [PubMed]
- Phipps, L.P.; De Marco, M.D.M.F.; Hernández-Triana, L.M.; Johnson, N.; Swainsbury, C.; Medlock, J.M.; Hansford, K.; Mitchell, S. Babesia canis detected in dogs and associated ticks from Essex. Vet. Rec. 2016, 178, 243–244. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.D.M.F.; Hernández-Triana, L.M.; Phipps, L.P.; Hansford, K.; Mitchell, E.S.; Cull, B.; Swainsbury, C.S.; Fooks, A.R.; Medlock, J.M.; Johnson, N. Emergence of Babesia canis in southern England. Parasites Vectors 2017, 10, 241. [Google Scholar] [CrossRef] [PubMed]
- Pietzsch, M.E.; Hansford, K.M.; Cull, B.; Jahfari, S.; Sprong, H.; Medlock, J.M. Detection of Dermacentor marginatus and a possible Rickettsia slovaca case in the United Kingdom—The risk of the visiting traveller. Travel Med. Infect. Dis. 2015, 13, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Foldvari, G.; Siroky, P.; Szekeres, S.; Majoros, G.; Sprong, H. Dermacentor reticulatus: A new vector on the rise. Parasites Vectors 2016, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautal, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Tick Tick-Borne Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Rainey, T.; Occi, J.L.; Robbins, R.G.; Egizi, A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) parasitizing a sheeo in New Jersey, United States. J. Med. Entomol. 2018, 55, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Pietzsch, M.E.; Cull, B.; Medlock, J.M.; Wall, R. Overwintering of the brown dog tick in residential properties in England–raising awareness. Vet. Rec. 2015, 177, 156. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Phipps, L.P.; Cull, B.; Pietzsch, M.E.; Medlock, J.M. Rhipicephalus sanguineus importation into the UK: Surveillance, risk, public health awareness and One Health response. Vet. Rec. 2017, 180, 119. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Pietzsch, M.; Cull, B.; Medlock, J.M. Brown dog tick infestation of a home in England. Vet. Rec. 2015, 176, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Jameson, L.J.; Medlock, J.M. Results of HPA tick surveillance in Great Britain. Vet. Rec. 2009, 165, 154. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Medlock, J.M.; Atkinson, B.; Santos-Silva, M.M. Importation of a Hyalomma lusitanicum tick into the UK on a dog. Vet. Rec. 2016, 179, 415. [Google Scholar] [CrossRef] [PubMed]
- Pietzsch, M.; Hansford, K.; Medlock, J.; Doggett, S.L. Australian paralysis tick imported on a traveller returning to the UK. Travel Med. Infect. Dis. 2014, 12, 196–197. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medlock, J.M.; Hansford, K.M.; Vaux, A.G.C.; Cull, B.; Gillingham, E.; Leach, S. Assessment of the Public Health Threats Posed by Vector-Borne Disease in the United Kingdom (UK). Int. J. Environ. Res. Public Health 2018, 15, 2145. https://doi.org/10.3390/ijerph15102145
Medlock JM, Hansford KM, Vaux AGC, Cull B, Gillingham E, Leach S. Assessment of the Public Health Threats Posed by Vector-Borne Disease in the United Kingdom (UK). International Journal of Environmental Research and Public Health. 2018; 15(10):2145. https://doi.org/10.3390/ijerph15102145
Chicago/Turabian StyleMedlock, Jolyon M., Kayleigh M. Hansford, Alexander G. C. Vaux, Ben Cull, Emma Gillingham, and Steve Leach. 2018. "Assessment of the Public Health Threats Posed by Vector-Borne Disease in the United Kingdom (UK)" International Journal of Environmental Research and Public Health 15, no. 10: 2145. https://doi.org/10.3390/ijerph15102145
APA StyleMedlock, J. M., Hansford, K. M., Vaux, A. G. C., Cull, B., Gillingham, E., & Leach, S. (2018). Assessment of the Public Health Threats Posed by Vector-Borne Disease in the United Kingdom (UK). International Journal of Environmental Research and Public Health, 15(10), 2145. https://doi.org/10.3390/ijerph15102145