Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture



Abstract
:1. Introduction
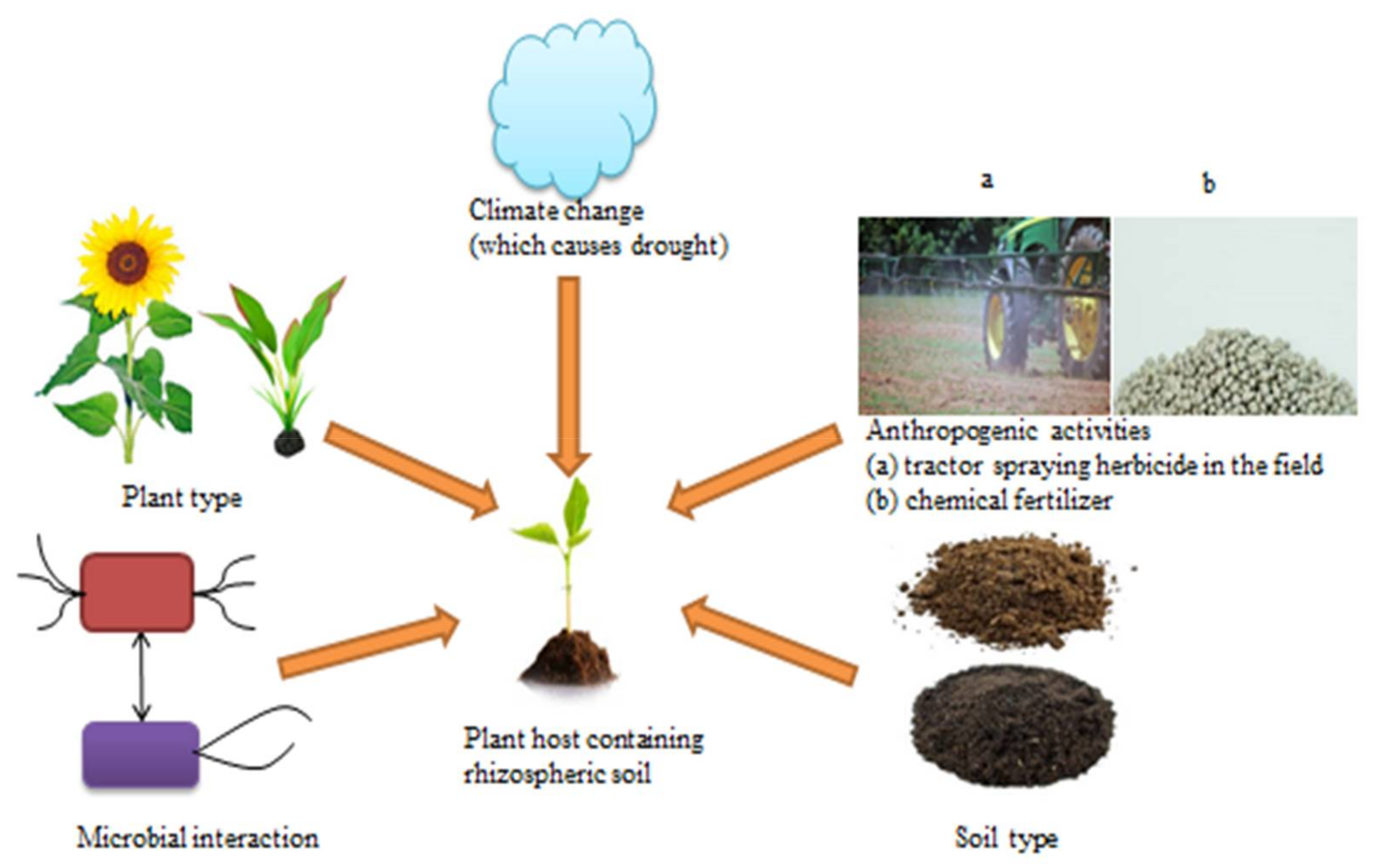
2. Modulators of Rhizosphere Microbiome of Agricultural Crops
Other Rhizosphere Microbiome Modulators: Climate Change and Anthropogenic Activities
3. Plant Endophytes and Their Ability to Fix Atmospheric Nitrogen
4. Rhizobiome as Plant Growth Promoters
5. The Effects of Rhizosphere Microbiome on Sustainable Agriculture and Food Security
5.1. Impact of Rhizobium Inoculation on Leguminous Crops Productivity
5.2. Impact of Rhizobium Inoculation on Mineral Nutrients Absorption by Leguminous Crops
5.3. Impact of Rhizobium Inoculation on Chlorophyll Concentration and Photosynthetic Activities of Leguminous Crops
6. Nexus of PGPR, Fe Acquisition, Plant Productivity and Pathogens Eradication
7. New PGPR That Are Related to Human Opportunistic Pathogens
8. Nexus of Rhizobia, Nodule Formation and SYM Pathway
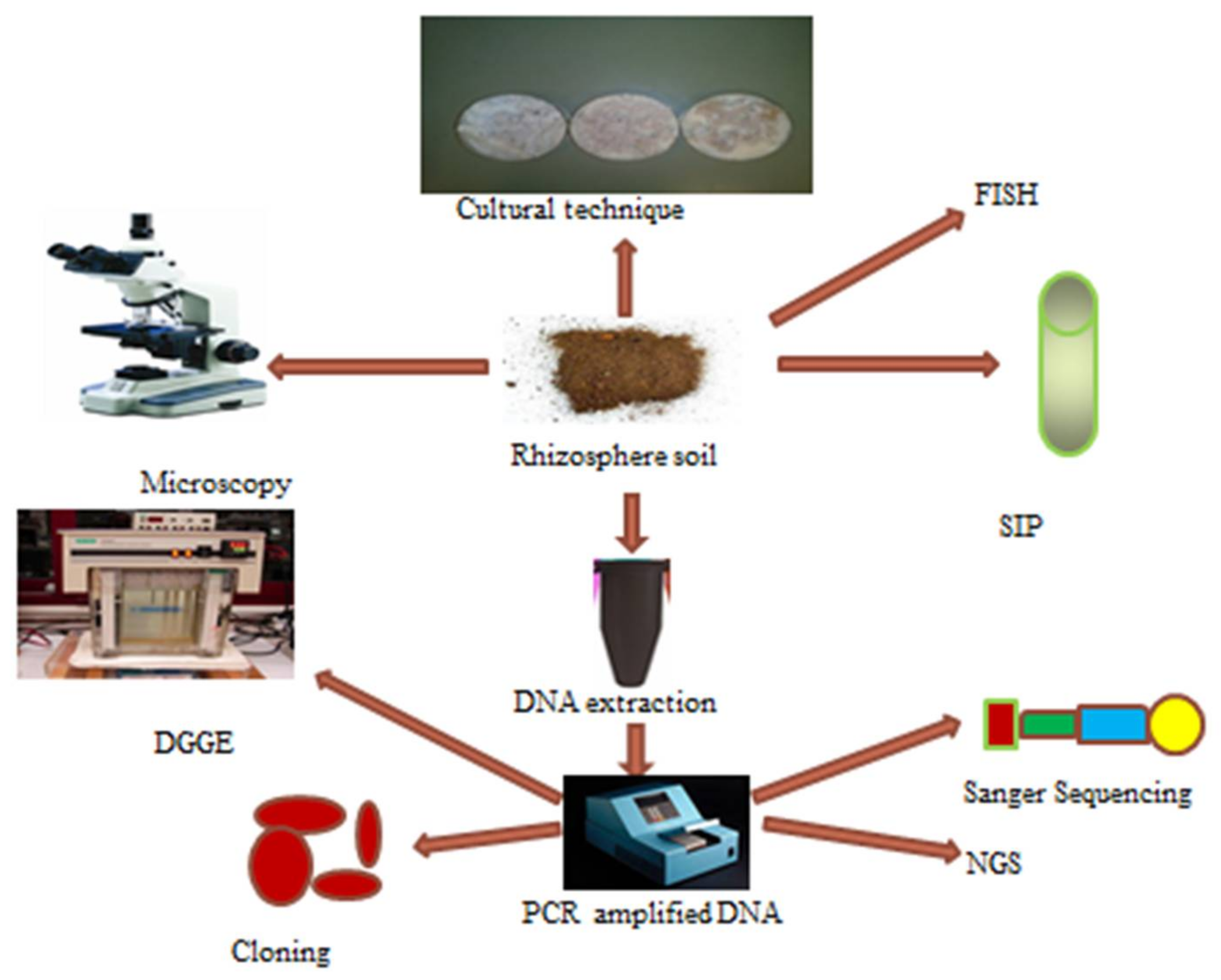
9. Techniques Use for Investigation of Rhizosphere Microbial Community Structure
9.1. Traditional Techniques
9.2. Molecular Techniques
10. Future Direction
11. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and sustainable agriculture: Exploring arbuscular mycorrhizal fungi. Appl. Microbiol. Biotechnol. 2017, 101, 4871–4881. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Bais, H. Probiotics for plants: Rhizospheric microbiome and plant fitness. Mol. Microb. Ecol. Rhiz. 2013, 1, 713–721. [Google Scholar]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- East, R. Microbiome: Soil science comes to life. Nature 2013, 501, S18–S19. [Google Scholar] [CrossRef] [PubMed]
- Dubois, O. The State of the World’s Land and Water Resources for Food and Agriculture: Managing Systems at Risk; Earthscan: London, UK, 2011. [Google Scholar]
- Hou, M.P.; Babalola, O.O. Evaluation of plant growth promoting potential of four rhizobacterial species for indigenous system. J. Cent. South Univ. 2013, 20, 164–171. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl.Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Bouffaud, M.L.; Kyselková, M.; Gouesnard, B.; Grundmann, G.; Muller, D.; Moenne-Loccoz, Y. Is diversification history of maize influencing selection of soil bacteria by roots? Mol. Ecol. 2012, 21, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional soil microbiome: Belowground solutions to an aboveground problem. Plant Physiol. 2014, 166, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E. Revealing structure and assembly cues for Arabidopsis root inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O.; Sanni, A.I.; Odhiambo, G.D.; Torto, B. Plant growth-promoting rhizobacteria do not pose any deleterious effect on cowpea and detectable amounts of ethylene are produced. World J. Microbiol. Biotechnol. 2007, 23, 747–752. [Google Scholar] [CrossRef]
- De Ridder-Duine, A.S.; Kowalchuk, G.A.; Gunnewiek, P.J.K.; Smant, W.; van Veen, J.A.; de Boer, W. Rhizosphere bacterial community composition in natural stands of Carex arenaria (sand sedge) is determined by bulk soil community composition. Soil Biol. Biochem. 2005, 37, 349–357. [Google Scholar] [CrossRef]
- Santos-González, J.C.; Nallanchakravarthula, S.; Alström, S.; Finlay, R.D. Soil, but not cultivar, shapes the structure of arbuscular mycorrhizal fungal assemblages associated with strawberry. Microb. Ecol. 2011, 62, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.R.; Fitak, R.R.; Munguia-Vega, A.; Racolta, A.; Martinson, V.G.; Dontsova, K. Abiotic factors shape microbial diversity in Sonoran Desert soils. Appl. Environ. Microbiol. 2012, 78, 7527–7537. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.; Götz, M.; Mrotzek, N.; Lottmann, J.; Berg, G.; Smalla, K. Effects of site and plant species on rhizosphere community structure as revealed by molecular analysis of microbial guilds. FEMS Microbiol. Ecol. 2006, 56, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Renoud, S.; Muller, D.; Babalola, O.O.; Prigent-Combaret, C. Alleviation of abiotic and biotic stresses in plants by Azospirillum. In Handbook for Azospirillum; Springer: Berlin/Heidelberg, Germany, 2015; pp. 333–365. [Google Scholar]
- Isobe, K.; Ohte, N. Ecological perspectives on microbes involved in N-cycling. Microbes Environ. 2014, 29, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Giraldo, L.; Bhattarai, K.; Pislariu, C.I.; Nakashima, J.; Jikumaru, Y.; Kamiya, Y.; Udvardi, M.K.; Monteros, M.J.; Dixon, R.A. Lignin modification leads to increased nodule numbers in alfalfa. Plant Physiol. 2014, 164, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Bahena, M.H.; Tejedor, C.; Martín, I.; Velázquez, E.; Peix, A. Endobacter medicaginis gen. nov., sp. nov., isolated from alfalfa nodules in an acidic soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.E.; Alonso-Vega, P.; Rodríguez, R.; Carro, L.; Cerda, E.; Alonso, P.; Martinez-Molina, E. The genus Micromonospora is widespread in legume root nodules: The example of Lupinus angustifolius. ISME J. 2010, 4, 1265–1281. [Google Scholar] [CrossRef] [PubMed]
- Pini, F.; Frascella, A.; Santopolo, L.; Bazzicalupo, M.; Biondi, E.G.; Scotti, C.; Mengoni, A. Exploring the plant-associated bacterial communities in Medicago sativa L. BMC Microbiol. 2012, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.Y.; Alsyeeh, A.-M.; Almalki, M.A.; Saleh, F.A. Characterization of the plant growth promoting bacterium, Enterobacter cloacae MSR1, isolated from roots of non-nodulating Medicago sativa. Saudi J. Biol. Sci. 2016, 23, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.; Piceno, Y.M.; Desantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; Von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Rincon-Florez, V.A.; Carvalhais, L.C.; Schenk, P.M. Culture-independent molecular tools for soil and rhizosphere microbiology. Diversity 2013, 5, 581–612. [Google Scholar] [CrossRef]
- Schlaeppi, K.; Dombrowski, N.; Oter, R.G.; van Themaat, E.V.L.; Schulze-Lefert, P. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Sci. USA 2014, 111, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Sanschagrin, S.; Maynard, C.; St-Arnaud, M.; Greer, C.W. Microbial expression profiles in the rhizosphere of willows depend on soil contamination. ISME J. 2014, 8, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Turner, T.R.; Ramakrishnan, K.; Walshaw, J.; Heavens, D.; Alston, M.; Swarbreck, D.; Osbourn, A.; Grant, A.; Poole, P.S. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013, 7, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Minz, D.; Ofek, M.; Hadar, Y. Plant rhizosphere microbial communities. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 56–84. [Google Scholar]
- Abbott, L.; Robsson, A. The effect of soil on the formation of VA mycorrhizas by two species of Glomus. Soil Res. 1985, 23, 253–261. [Google Scholar] [CrossRef]
- İnceoğlu, Ö.; Salles, J.F.; van Elsas, J.D. Soil and cultivar type shape the bacterial community in the potato rhizosphere. Microb. Ecol. 2012, 63, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Cooperband, L.; Stone, A. Dynamic relationships between soil properties and foliar disease as affected by annual additions of organic amendment to a sandy-soil vegetable production system. Soil Biol. Biochem. 2005, 37, 1343–1357. [Google Scholar] [CrossRef]
- Toljander, J.F.; Santos-González, J.C.; Tehler, A.; Finlay, R.D. Community analysis of arbuscular mycorrhizal fungi and bacteria in the maize mycorrhizosphere in a long-term fertilization trial. FEMS Microbiol. Ecol. 2008, 65, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Relative roles of niche and neutral processes in structuring a soil microbial community. ISME J. 2010, 4, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.V.; Van Veen, J.; Van Elsas, J. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Gelsomino, A.; Keijzer-Wolters, A.C.; Cacco, G.; van Elsas, J.D. Assessment of bacterial community structure in soil by polymerase chain reaction and denaturing gradient gel electrophoresis. J. Microbiol. Methods 1999, 38, 1–15. [Google Scholar] [CrossRef]
- Latour, X.; Corberand, T.; Laguerre, G.; Allard, F.; Lemanceau, P. The composition of fluorescent pseudomonad populations associated with roots is influenced by plant and soil type. Appl. Environ. Microbiol. 1996, 62, 2449–2456. [Google Scholar] [PubMed]
- Hardoim, P.R.; Hardoim, C.C.; Van Overbeek, L.S.; Van Elsas, J.D. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef] [PubMed]
- Lareen, A.; Burton, F.; Schäfer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Bever, J.D.; Platt, T.G.; Morton, E.R. Microbial population and community dynamics on plant roots and their feedbacks on plant communities. Annu. Rev. Microbiol. 2012, 66, 265–283. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Mathesius, U. The role of flavonoids in root–rhizosphere signalling: Opportunities and challenges for improving plant–microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Montaño, F.; Guasch-Vidal, B.; González-Barroso, S.; López-Baena, F.J.; Cubo, T.; Ollero, F.J.; Gil-Serrano, A.M.; Rodríguez-Carvajal, M.Á.; Bellogín, R.A.; Espuny, M.R. Nodulation-gene-inducing flavonoids increase overall production of autoinducers and expression of N-acyl homoserine lactone synthesis genes in rhizobia. Res. Microbiol. 2011, 162, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Bressan, M.; Roncato, M.-A.; Bellvert, F.; Comte, G.; Haichar, F.Z.; Achouak, W.; Berge, O. Exogenous glucosinolate produced by Arabidopsis thaliana has an impact on microbes in the rhizosphere and plant roots. ISME J. 2009, 3, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Quintana, N.; El Kassis, E.G.; Kim, H.K.; Choi, Y.H.; Sugiyama, A.; Verpoorte, R.; Martinoia, E.; Manter, D.K.; Vivanco, J.M. An ABC transporter mutation alters root exudation of phytochemicals that provoke an overhaul of natural soil microbiota. Plant Physiol. 2009, 151, 2006–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, N.; Piceno, Y.; Ding, G.-C.; Meincke, R.; Heuer, H.; Berg, G.; Schloter, M.; Andersen, G.; Smalla, K. PhyloChip hybridization uncovered an enormous bacterial diversity in the rhizosphere of different potato cultivars: Many common and few cultivar-dependent taxa. FEMS Microbiol. Ecol. 2011, 75, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.B.; Lutz, M.P.; Frapolli, M.; Péchy-Tarr, M.; Rochat, L.; Keel, C.; Défago, G.; Maurhofer, M. Interplay between wheat cultivars, biocontrol pseudomonads, and soil. Appl. Environ. Microbiol. 2010, 76, 6196–6204. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.-H.; Mazzola, M. Modification of fluorescent pseudomonad community and control of apple replant disease induced in a wheat cultivar-specific manner. Appl. Soil Ecol. 2003, 24, 57–72. [Google Scholar] [CrossRef]
- Okubara, P.A.; Bonsall, R.F. Accumulation of Pseudomonas-derived 2, 4-diacetylphloroglucinol on wheat seedling roots is influenced by host cultivar. Biol. Control 2008, 46, 322–331. [Google Scholar] [CrossRef]
- Blee, K.; Hein, J.; Wolfe, G.V. Arabidopsis thaliana: A useful but limited model to investigate stress impacts on rhizosphere community composition and function. Mol. Microbial Ecol. Rhiz. 2013, 1, 265–270. [Google Scholar]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 2010, 8, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Crowther, T.W.; Thomas, S.M.; Maynard, D.S.; Baldrian, P.; Covey, K.; Frey, S.D.; Van Diepen, L.T.; Bradford, M.A. Biotic interactions mediate soil microbial feedbacks to climate change. Proc. Natl. Acad. Sci. USA 2015, 112, 7033–7038. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Philippot, L.; Neumann, G.; Marhan, S.; Kandeler, E. Local response of bacterial densities and enzyme activities to elevated atmospheric CO2 and different N supply in the rhizosphere of Phaseolus vulgaris L. Soil Biol. Biochem. 2008, 40, 1225–1234. [Google Scholar] [CrossRef]
- Stevnbak, K.; Scherber, C.; Gladbach, D.J.; Beier, C.; Mikkelsen, T.N.; Christensen, S. Interactions between above-and belowground organisms modified in climate change experiments. Nat. Clim. Chang. 2012, 2, 805–808. [Google Scholar] [CrossRef]
- Drigo, B.; Kowalchuk, G.A.; Knapp, B.A.; Pijl, A.S.; Boschker, H.T.; Veen, J.A. Impacts of 3 years of elevated atmospheric CO2 on rhizosphere carbon flow and microbial community dynamics. Glob. Chang. Biol. 2013, 19, 621–636. [Google Scholar] [CrossRef] [PubMed]
- De Vries, F.T.; Liiri, M.E.; Bjørnlund, L.; Bowker, M.A.; Christensen, S.; Setälä, H.M.; Bardgett, R.D. Land use alters the resistance and resilience of soil food webs to drought. Nat. Clim. Chang. 2012, 2, 276–280. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of warming and drought on grassland microbial communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Giller, K.E. Nitrogen Fixation in Tropical Cropping Systems; Cabi: Wallingford, UK, 2001. [Google Scholar]
- Serraj, R.; Adu-Gyamfi, J. Role of symbiotic nitrogen fixation in the improvement of legume productivity under stressed environments. West Afr. Appl. Ecol. 2004, 6, 95–109. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Pellizari, V.H.; Mueller, R.; Baek, K.; Jesus, E.D.C.; Paula, F.S.; Mirza, B.; Hamaoui, G.S.; Tsai, S.M.; Feigl, B. Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2013, 110, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Labbate, M.; Seymour, J.R.; Lauro, F.; Brown, M.V. Anthropogenic impacts on the microbial ecology and function of aquatic environments. Front. Microbiol. 2016, 7, 1044. [Google Scholar] [CrossRef] [PubMed]
- Gyamfi, S.; Pfeifer, U.; Stierschneider, M.; Sessitsch, A. Effects of transgenic glufosinate-tolerant oilseed rape (Brassica napus) and the associated herbicide application on eubacterial and Pseudomonas communities in the rhizosphere. FEMS Microbiol. Ecol. 2002, 41, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, B.; Caul, S.; Thompson, J.; Hackett, C.; Cortet, J.; Pernin, C.; Krogh, P.H. Soil microbial and faunal responses to herbicide tolerant maize and herbicide in two soils. Plant Soil 2008, 308, 93–103. [Google Scholar] [CrossRef]
- Haney, R.; Senseman, S.; Krutz, L.; Hons, F. Soil carbon and nitrogen mineralization as affected by atrazine and glyphosate. Biol. Fertil. Soils 2002, 35, 35–40. [Google Scholar]
- Lupwayi, N.; Harker, K.; Clayton, G.; Turkington, T.; Rice, W.; O’Donovan, J. Soil microbial biomass and diversity after herbicide application. Can. J. Plant Sci. 2004, 84, 677–685. [Google Scholar] [CrossRef]
- Kremer, R.J.; Means, N.E. Glyphosate and glyphosate-resistant crop interactions with rhizosphere microorganisms. Eur. J. Agron. 2009, 31, 153–161. [Google Scholar] [CrossRef]
- Pessagno, R.C.; Sánchez, R.M.T.; Dos Santos Afonso, M. Glyphosate behavior at soil and mineral–water interfaces. Environ. Pollut. 2008, 153, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Barja, B.; Dos Santos Afonso, M. Aminomethylphosphonic acid and glyphosate adsorption onto goethite: A comparative study. Environ. Sci. Technol. 2005, 39, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R. Herbicides and herbicide degradates in shallow groundwater and the Cedar River near a municipal well field, Cedar Rapids, Iowa. Sci. Total Environ. 2000, 248, 241–253. [Google Scholar] [CrossRef]
- Satsuma, K. Complete biodegradation of atrazine by a microbial community isolated from a naturally derived river ecosystem (microcosm). Chemosphere 2009, 77, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Toyota, K.; Ritz, K.; Kuninaga, S.; Kimura, M. Impact of fumigation with metam sodium upon soil microbial community structure in two Japanese soils. Soil Sci. Plant Nutr. 1999, 45, 207–223. [Google Scholar] [CrossRef]
- Ratcliff, A.W.; Busse, M.D.; Shestak, C.J. Changes in microbial community structure following herbicide (glyphosate) additions to forest soils. Appl. Soil Ecol. 2006, 34, 114–124. [Google Scholar] [CrossRef]
- Hernandez, A.; Garcia-Plazaola, J.I.; Becerril, J.M. Glyphosate effects on phenolic metabolism of nodulated soybean (Glycine max L. Merr.). J. Agric. Food Chem. 1999, 47, 2920–2925. [Google Scholar] [CrossRef] [PubMed]
- Moorman, T.B.; Becerril, J.M.; Lydon, J.; Duke, S.O. Production of hydroxybenzoic acids by Bradyrhizobium japonicum strains after treatment with glyphosate. J. Agric. Food Chem. 1992, 40, 289–293. [Google Scholar] [CrossRef]
- Liu, C.-M.; McLean, P.; Sookdeo, C.; Cannon, F. Degradation of the herbicide glyphosate by members of the family Rhizobiaceae. Appl. Environ. Microbiol. 1991, 57, 1799–1804. [Google Scholar] [PubMed]
- Bohm, G.M.; Alves, B.J.; Urquiaga, S.; Boddey, R.M.; Xavier, G.R.; Hax, F.; Rombaldi, C.V. Glyphosate-and imazethapyr-induced effects on yield, nodule mass and biological nitrogen fixation in field-grown glyphosate-resistant soybean. Soil Biol. Biochem. 2009, 41, 420–422. [Google Scholar] [CrossRef]
- Zobiole, L.; Oliveira, R., Jr.; Kremer, R.; Constantin, J.; Yamada, T.; Castro, C.; Oliveira, F.A.; Oliveira, A., Jr. Effect of glyphosate on symbiotic N2 fixation and nickel concentration in glyphosate-resistant soybeans. Appl. Soil Ecol. 2010, 44, 176–180. [Google Scholar] [CrossRef]
- King, C.A.; Purcell, L.C.; Vories, E.D. Plant growth and nitrogenase activity of glyphosate-tolerant soybean in response to foliar glyphosate applications. Agron. J. 2001, 93, 179–186. [Google Scholar] [CrossRef]
- Powell, J.R.; Levy Booth, D.J.; Gulden, R.H.; Asbil, W.L.; Campbell, R.G.; Dunfield, K.E.; Hamill, A.S.; Hart, M.M.; Lerat, S.; Nurse, R.E.; et al. Effects of genetically modified, herbicide tolerant crops and their management on soil food web properties and crop litter decomposition. J. Appl. Ecol. 2009, 46, 388–396. [Google Scholar] [CrossRef]
- Crouzet, O.; Batisson, I.; Besse-Hoggan, P.; Bonnemoy, F.; Bardot, C.; Poly, F.; Bohatier, J.; Mallet, C. Response of soil microbial communities to the herbicide mesotrione: A dose-effect microcosm approach. Soil Biol. Biochem. 2010, 42, 193–202. [Google Scholar] [CrossRef]
- Dick, R.; Lorenz, N.; Wojno, M.; Lane, M. Microbial dynamics in soils under long-term glyphosate tolerant cropping systems. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010. [Google Scholar]
- Ikeda, S.; Okubo, T.; Anda, M.; Nakashita, H.; Yasuda, M.; Sato, S.; Kaneko, T.; Tabata, S.; Eda, S.; Momiyama, A. Community-and genome-based views of plant-associated bacteria: Plant–bacterial interactions in soybean and rice. Plant Cell Physiol. 2010, 51, 1398–1410. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Okubo, T.; Kaneko, T.; Inaba, S.; Maekawa, T.; Eda, S.; Sato, S.; Tabata, S.; Mitsui, H.; Minamisawa, K. Community shifts of soybean stem-associated bacteria responding to different nodulation phenotypes and N levels. ISME J. 2010, 4, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Sasaki, K.; Okubo, T.; Yamashita, A.; Terasawa, K.; Bao, Z.; Liu, D.; Watanabe, T.; Murase, J.; Asakawa, S. Low nitrogen fertilization adapts rice root microbiome to low nutrient environment by changing biogeochemical functions. Microbes Environ. 2014, 29, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.; Poole, P.S. Transport and metabolism in legume-rhizobia symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed]
- Tkacz, A.; Poole, P. Role of root microbiota in plant productivity. J. Exp. Bot. 2015, 66, 2167–2175. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ping, S.; Peng, J.; Han, Y.; Li, L.; Yang, J.; Dou, Y.; Li, Y.; Fan, H.; Fan, Y.; et al. Global transcriptional analysis of nitrogen fixation and ammonium repression in root-associated Pseudomonas stutzeri A1501. BMC Genom. 2010, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Reinhold-Hurek, B. Transcriptional profiling of nitrogen fixation and the role of NifA in the diazotrophic endophyte Azoarcus sp. strain BH72. PLoS ONE 2014, 9, e86527. [Google Scholar] [CrossRef] [PubMed]
- Crook, M.B.; Mitra, S.; Ané, J.-M.; Sadowsky, M.J.; Gyaneshwar, P. Complete genome sequence of the Sesbania symbiont and rice growth-promoting endophyte Rhizobium sp. strain IRBG74. Genome Announc. 2013, 1, e00934-13. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhu, H. Are common symbiosis genes required for endophytic rice-rhizobial interactions? Plant Signal Behav. 2013, 8, e25453. [Google Scholar] [CrossRef] [PubMed]
- Venkateshwaran, M.; Volkening, J.D.; Sussman, M.R.; Ané, J.-M. Symbiosis and the social network of higher plants. Curr. Opin. Plant Biol. 2013, 16, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, F.O.; Monteiro, R.A.; Wassem, R.; Cruz, L.M.; Ayub, R.A.; Colauto, N.B.; Fernandez, M.A.; Fungaro, M.H.P.; Grisard, E.C.; Hungria, M. Genome of Herbaspirillum seropedicae strain SmR1, a specialized diazotrophic endophyte of tropical grasses. PLoS Genet. 2011, 7, e1002064. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.A.; Balsanelli, E.; Faoro, H.; Cruz, L.M.; Wassem, R.; de Baura, V.A.; Weiss, V.; Yates, M.G.; Madeira, H.M.; Pereira-Ferrari, L.; et al. The type III secretion system is necessary for the development of a pathogenic and endophytic interaction between Herbaspirillum rubrisubalbicans and Poaceae. BMC Microbiol. 2012, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, S.; Kaneko, T.; Sato, S.; Saeki, K. Hijacking of leguminous nodulation signaling by the rhizobial type III secretion system. Proc. Natl. Acad. Sci. USA 2013, 110, 17131–17136. [Google Scholar] [CrossRef] [PubMed]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Schroth, M.N. Plant growth-promoting rhizobacteria on radishes. In Proceedings of the 4th International Conference on Plant Pathogenic Bacteria, Gilbert Clarey, Tours, France, 27 August–2 September 1978. [Google Scholar]
- Conway, G.R.; Pretty, J.N. Unwelcome Harvest: Agriculture and Pollution; Routledge: Abingdon, UK, 2013; 676p. [Google Scholar]
- Rascovan, N.; Carbonetto, B.; Perrig, D.; Díaz, M.; Canciani, W.; Abalo, M.; Alloati, J.; González-Anta, G.; Zquez, M.P. Integrated analysis of root microbiomes of soybean and wheat from agricultural fields. Sci. Rep. 2016, 6, 28084. [Google Scholar] [CrossRef] [PubMed]
- Bevivino, A.; Sarrocco, S.; Dalmastri, C.; Tabacchioni, S.; Cantale, C.; Chiarini, L. Characterization of a free-living maize-rhizosphere population of Burkholderia cepacia: Effect of seed treatment on disease suppression and growth promotion of maize. FEMS Microbiol. Ecol. 1998, 27, 225–237. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Babikova, Z.; Johnson, D.; Bruce, T.; Pickett, J.; Gilbert, L. Underground allies: How and why do mycelial networks help plants defend themselves? BioEssays 2014, 36, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Wani, P.A.; Khan, M.S. Nickel detoxification and plant growth promotion by multi metal resistant plant growth promoting Rhizobium species RL9. Bull. Environ. Contam. Toxicol. 2013, 91, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. Biotecholology 2015, 5, 355–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uma, C.; Sivagurunathan, P.; Sangeetha, D. Performance of bradyrhizobial isolates under drought conditions. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 228–232. [Google Scholar]
- Rodrigues, E.P.; Rodrigues, L.S.; de Oliveira, A.L.M.; Baldani, V.L.D.; dos Santos Teixeira, K.R.; Urquiaga, S.; Reis, V.M. Azospirillum amazonense inoculation: Effects on growth, yield and N2 fixation of rice (Oryza sativa L.). Plant Soil. 2008, 302, 249–261. [Google Scholar] [CrossRef]
- Radwan, S.; Dashti, N.; El-Nemr, I. Enhancing the growth of Vicia faba plants by microbial inoculation to improve their phytoremediation potential for oily desert areas. Int. J. Phytoremediat. 2005, 7, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Zamioudis, C.; Pieterse, C.M. Modulation of host immunity by beneficial microbes. Mol. Plant-Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Shaharoona, B.; Jamro, G.; Zahir, Z.; Arshad, M.; Memon, K. Effectiveness of various Pseudomonas spp. and Burkholderia caryophylli containing ACC-Deaminase for improving growth and yield of wheat (Triticum aestivum I.). J. Microbiol. Biotechnol. 2007, 17, 1300. [Google Scholar] [PubMed]
- Mfilinge, A.; Mtei, K.; Ndakidemi, P. Effect of Rhizobium inoculation and supplementation with phosphorus and potassium on growth and total leaf chlorophyll (Chl) content of bush bean Phaseolus vulgaris, L. Agric. Sci. 2014, 5, 1413. [Google Scholar] [CrossRef]
- Ravikumar, R. Growth effects of Rhizobium inoculation in some legume plants. Int. J.Curr. Sci. 2012, 1, 1–6. [Google Scholar]
- Fan, L.-M.; Ma, Z.-Q.; Liang, J.-Q.; Li, H.-F.; Wang, E.-T.; Wei, G.-H. Characterization of a copper-resistant symbiotic bacterium isolated from Medicago lupulina growing in mine tailings. Bioresour. Technol. 2011, 102, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Khan, M.S. Effect of tebuconazole tolerant and plant growth promoting Rhizobium isolate MRP1 on pea Rhizobium symbiosis. Sci. Hortic. 2011, 129, 266–272. [Google Scholar] [CrossRef]
- Gupta, V.V. Beneficial microorganisms for sustainable agriculture. Microbiol. Aust. 2012, 113, 113–115. [Google Scholar]
- Bhattacharyya, P.; Jha, D. Plant growth promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sust. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Lau, J.A.; Lennon, J.T. Evolutionary ecology of plant microbe interactions: Soil microbial structure alters selection on plant traits. New Phytol. 2011, 192, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; Klironomos, J.N.; HilleRisLambers, J.; Kinkel, L.L.; Reich, P.B.; Xiao, K.; Rillig, M.C.; Sikes, B.A.; Callaway, R.M.; Mangan, S.A. Soil microbes drive the classic plant diversity productivity pattern. Ecology 2011, 92, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Jansa, J.; Schmid, B.; van der Heijden, M.G. Below ground biodiversity effects of plant symbionts support aboveground productivity. Ecol. Lett. 2011, 14, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.R.; Sahin, S.; Düzdemir, O.; Kandemir, N. Selection of chickpea cultivars with agronomic phosphorus (P) utilization characters as influenced by Rhizobium inoculation. Sci. Res. Essays 2013, 8, 676–681. [Google Scholar]
- Nyoki, D.; Ndakidemi, P.A. Effects of phosphorus and Bradyrhizobium japonicum on growth and chlorophyll content of cowpea (Vigna unguiculata (L) Walp). Am. J. Exp. Agric. 2014, 4, 1120–1136. [Google Scholar] [CrossRef]
- Tairo, E.V.; Ndakidemi, P.A. Bradyrhizobium japonicum inoculation and phosphorus supplementation on growth and chlorophyll accumulation in soybean (Glycine max L.). Am. J. Plant Sci. 2013, 4, 2281. [Google Scholar] [CrossRef]
- Remans, R.; Ramaekers, L.; Schelkens, S.; Hernandez, G.; Garcia, A.; Reyes, J.L.; Mendez, N.; Toscano, V.; Mulling, M.; Galvez, G.; Vanderleyden, J. Effect of Rhizobium–Azospirillum coinoculation on nitrogen fixation and yield of two contrasting Phaseolus vulgaris L. genotypes cultivated across different environments in Cuba. Plant Soil 2008, 312, 25–37. [Google Scholar] [CrossRef]
- Tchebotar, V.; Kang, U.; Asis, C., Jr.; Akao, S. The use of GUS-reporter gene to study the effect of Azospirillum-Rhizobium coinoculation on nodulation of white clover. Biol. Fertil. Soils 1998, 27, 349–352. [Google Scholar] [CrossRef]
- Cassan, F.; Perrig, D.; Sgroy, V.; Masciarelli, O.; Penna, C.; Luna, V. Azospirillum brasilense Az39 and Bradyrhizobium japonicum E109, inoculated singly or in combination, promote seed germination and early seedling growth in corn (Zea mays L.) and soybean (Glycine max L.). Eur. J. Soil Biol. 2009, 45, 28–35. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; de Cordoba, F.J.F.; Espuny, M.R.; Carvajal, M.A.R.; Díaz, M.E.S.; Serrano, A.M.G.; Okon, Y.; Megias, M. Effect of Azospirillum brasilense coinoculated with Rhizobium on Phaseolus vulgaris flavonoids and Nod factor production under salt stress. Soil Biol. Biochem. 2008, 40, 2713–2721. [Google Scholar] [CrossRef]
- Akhtar, N.; Qureshi, M.; Iqbal, A.; Ahmad, M.; Khan, K. Influence of Azotobacter and IAA on symbiotic performance of Rhizobium and yield parameters of lentil. J. Agric. Res. 2012, 50, 361–372. [Google Scholar]
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Bisht, J.; Kundu, S.; Gupta, H.S. Coinoculation of Bacillus thuringeinsis-KR1 with Rhizobium leguminosarum enhances plant growth and nodulation of pea (Pisum sativum L.) and lentil (Lens culinaris L.). World J. Microbiol. Biotechnol. 2009, 25, 753–761. [Google Scholar] [CrossRef]
- Stajkovic, O.; Delic, D.; Josic, D.; Kuzmanovic, D.; Rasulic, N.; Knezevic-Vukcevic, J. Improvement of common bean growth by co-inoculation with Rhizobium and plant growth-promoting bacteria. Rom. Biotechnol. Lett. 2011, 16, 5919–5926. [Google Scholar]
- Singh, G.; Sekhon, H.; Sharma, P. Effect of irrigation and biofertilizer on water use, nodulation, growth and yield of chickpea (Cicer arietinum L.). Arch. Agron. Soil Sci. 2011, 57, 715–726. [Google Scholar] [CrossRef]
- Elkoca, E.; Kantar, F.; Sahin, F. Influence of nitrogen fixing and phosphorus solubilizing bacteria on the nodulation, plant growth, and yield of chickpea. J. Plant Nutr. 2007, 31, 157–571. [Google Scholar] [CrossRef]
- Rajendran, G.; Sing, F.; Desai, A.J.; Archana, G. Enhanced growth and nodulation of pigeon pea by co-inoculation of Bacillus strains with Rhizobium spp. Bioresour. Technol. 2008, 99, 4544–4550. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.L.; O’Hara, G.W.; Bräu, L. Enhanced nodulation and symbiotic effectiveness of Medicago truncatula when co-inoculated with Pseudomonas fluorescens WSM3457 and Ensifer (Sinorhizobium) medicae WSM419. Plant Soil 2011, 348, 245. [Google Scholar] [CrossRef]
- Verma, J.P.; Yadav, J.; Tiwari, K.N.; Kumar, A. Effect of indigenous Mesorhizobium spp. and plant growth promoting rhizobacteria on yields and nutrients uptake of chickpea (Cicer arietinum L.) under sustainable agriculture. Ecol. Eng. 2013, 51, 282–286. [Google Scholar] [CrossRef]
- Verma, J.P.; Yadav, J.; Tiwari, K.N.; Jaiswal, D.K. Evaluation of plant growth promoting activities of microbial strains and their effect on growth and yield of chickpea (Cicer arietinum L.) in India. Soil Biol. Biochem. 2014, 70, 33–37. [Google Scholar] [CrossRef]
- Vance, C.P.; Graham, P.H.; Allan, D.L. Biological nitrogen fixation: Phosphorus-A critical future need? In Nitrogen Fixation: From Molecules to Crop Productivity; Springer: Dordrecht, The Netherlands, 2000; pp. 509–514. [Google Scholar]
- Rudresh, D.; Shivaprakash, M.; Prasad, R. Effect of combined application of Rhizobium, phosphate solubilizing bacterium and Trichoderma spp. on growth, nutrient uptake and yield of chickpea (Cicer aritenium L.). Appl. Soil Ecol. 2005, 28, 139–146. [Google Scholar] [CrossRef]
- Clayton, G.; Rice, W.; Lupwayi, N.; Johnston, A.; Lafond, G.; Grant, C.; Walley, F. Inoculant formulation and fertilizer nitrogen effects on field pea: Nodulation, N2 fixation and nitrogen partitioning. Can. J. Plant Sci. 2004, 84, 79–88. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Hameeda, B.; Harini, G.; Rupela, O.; Wani, S.; Reddy, G. Growth promotion of maize by phosphate-solubilizing bacteria isolated from composts and macrofauna. Microbiol. Res. 2008, 163, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhakar, P.; Chattopadhyay, G.; Gangwar, S.; Ghosh, J. Effect of foliar application of Azotobacter, Azospirillum and Beijerinckia on leaf yield and quality of mulberry (Morus alba). J. Agric. Sci. 2000, 134, 227–234. [Google Scholar] [CrossRef]
- Mehnaz, S.; Lazarovits, G. Inoculation effects of Pseudomonas putida, Gluconacetobacter azotocaptans, and Azospirillum lipoferum on corn plant growth under greenhouse conditions. Microb. Ecol. 2006, 51, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Makoi, J.H.; Bambara, S.; Ndakidemi, P.A. Rhizobium inoculation and the supply of molybdenum and lime affect the uptake of macroelements in common bean (‘P. Vulgaris L.’) plants. Aust. J. Crop Sci. 2013, 7, 784. [Google Scholar]
- Kisinyo, P.; Gudu, S.; Othieno, C.; Okalebo, J.; Opala, P.; Maghanga, J.; Agalo, J.; Ng, W.; Kisinyo, J.; Osiyo, R. Effects of lime, phosphorus and rhizobia on Sesbania sesban performance in a Western Kenyan acid soil. Afr. J. Agric. Res. 2012, 7, 2800–2809. [Google Scholar] [CrossRef]
- Salisbury, F.B.; Ross, C.W. Plant Physiology, 4th ed.; Wadsworth: Belmont, CA, USA, 1992; pp. 23–52. [Google Scholar]
- Von Wirén, N.; Khodr, H.; Hider, R.C. Hydroxylated phytosiderophore species possess an enhanced chelate stability and affinity for iron (III). Plant Physiol. 2000, 124, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Bar-Ness, E.; Chen, Y.; Hadar, Y.; Marschner, H.; Römheld, V. Siderophores of Pseudomonas putida as an iron source for dicot and monocot plants. In Iron Nutrition and Interactions in Plants; Springer: Dordrecht, The Netherlands, 1991; pp. 271–281. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Duijff, B.; De Kogel, W.; Bakker, P.; Schippers, B. Influence of pseudobactin 358 on the iron nutrition of barley. Soil Biol. Biochem. 1994, 26, 1681–1688. [Google Scholar] [CrossRef]
- Masalha, J.; Kosegarten, H.; Elmaci, Ö.; Mengel, K. The central role of microbial activity for iron acquisition in maize and sunflower. Biol. Fertil. Soils 2000, 30, 433–439. [Google Scholar] [CrossRef]
- Bar-Ness, E.; Hadar, Y.; Chen, Y.; Römheld, V.; Marschner, H. Short-term effects of rhizosphere microorganisms on Fe uptake from microbial siderophores by maize and oat. Plant Physiol. 1992, 100, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Traxler, M.F.; Seyedsayamdost, M.R.; Clardy, J.; Kolter, R. Interspecies modulation of bacterial development through iron competition and siderophore piracy. Mol. Microbiol. 2012, 86, 628–644. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.B.; Das, S.; Dangar, T.K.; Adhya, T.K. ACC deaminase and IAA producing growth promoting bacteria from the rhizosphere soil of tropical rice plants. J. Basic Microbiol. 2013, 53, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Eberl, L.; Hartmann, A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 2005, 7, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Alavi, P.; Starcher, M.R.; Thallinger, G.G.; Zachow, C.; Müller, H.; Berg, G. Stenotrophomonas comparative genomics reveals genes and functions that differentiate beneficial and pathogenic bacteria. BMC Genom. 2014, 15, 482. [Google Scholar] [CrossRef] [PubMed]
- Suckstorff, I.; Berg, G. Evidence for dose dependent effects on plant growth by Stenotrophomonas strains from different origins. J. Appl. Microbiol. 2003, 95, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.; Bakker, P.; Pieterse, C. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Morales, G.; Escalante, R.; Campanario, E.; Sastre, L.; Martinez, J.L. Overexpression of the multidrug efflux pump SmeDEF impairs Stenotrophomonas maltophilia physiology. J. Antimicrob. Chem. 2004, 53, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.-W.; Rahme, L.G.; Sternberg, J.A.; Tompkins, R.G.; Ausubel, F.M. Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc. Natl. Acad. Sci. USA 1999, 96, 2408–2413. [Google Scholar] [CrossRef] [PubMed]
- Dörr, J.; Hurek, T.; Reinhold Hurek, B. Type IV pili are involved in plant–microbe and fungus–microbe interactions. Mol. Microbiol. 1998, 30, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Denton, M.; Kerr, K.G. Microbiological and clinical aspects of infection associated with Stenotrophomonas maltophilia. Clin. Microbiol. Rev. 1998, 11, 57–80. [Google Scholar] [PubMed]
- Gyaneshwar, P.; Hirsch, A.M.; Moulin, L.; Chen, W.-M.; Elliott, G.N.; Bontemps, C.; Estrada-de los Santos, P.; Gross, E.; dos Reis, F.B., Jr.; Sprent, J.I.; et al. Legume nodulating betaproteobacteria: Diversity, host range, and future prospects. Mol. Plant-Microbe Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef] [PubMed]
- Gavrin, A.; Jansen, V.; Ivanov, S.; Bisseling, T.; Fedorova, E. ARP2/3-mediated actin nucleation associated with symbiosome membrane is essential for the development of symbiosomes in infected cells of Medicago truncatula root nodules. Mol. Plant-Microbe Interact. 2015, 28, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J. Phylogenetic perspectives on the origins of nodulation. Mol. Plant-Microbe Interact. 2011, 24, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Schornack, S.; Marsh, J.F.; Gobbato, E.; Schwessinger, B.; Eastmond, P.; Schultze, M.; Kamoun, S.; Oldroyd, G.E. A common signaling process that promotes mycorrhizal and oomycete colonization of plants. Curr. Biol. 2012, 22, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Taulé, C.; Mareque, C.; Barlocco, C.; Hackembruch, F.; Reis, V.M.; Sicardi, M.; Battistoni, F. The contribution of nitrogen fixation to sugarcane (Saccharum officinarum L.), and the identification and characterization of part of the associated diazotrophic bacterial community. Plant Soil 2012, 356, 35–49. [Google Scholar] [CrossRef]
- Berlec, A. Novel techniques and findings in the study of plant microbiota: Search for plant probiotics. Plant Sci. 2012, 193, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Dini-Andreote, F.; van Elsas, J.D. Back to the basics: The need for ecophysiological insights to enhance our understanding of microbial behaviour in the rhizosphere. Plant Soil 2013, 373, 1–15. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of next-generation sequencing systems. BioMed Res. Int. 2012, 2012, 251364. [Google Scholar] [CrossRef] [PubMed]
- Uhlik, O.; Leewis, M.-C.; Strejcek, M.; Musilova, L.; Mackova, M.; Leigh, M.B.; Macek, T. Stable isotope probing in the metagenomics era: A bridge towards improved bioremediation. Biotechnol. Adv. 2013, 31, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Barret, M.; Morrissey, J.P.; O’Gara, F. Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence. Biol. Fertil. Soils 2011, 47, 729. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Wilhartitz, I.C.; Schneider, T.; Roschitzki, B.; Schmid, E.; Eberl, L.; Riedel, K.; Zechmeister-Boltenstern, S. Soil metaproteomics—Comparative evaluation of protein extraction protocols. Soil Biol. Biochem. 2012, 54, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmeder, C.A.; de Vos, W.M. Metaproteomics of our microbiome—Developing insight in function and activity in man and model systems. J. Proteom. 2014, 97, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- De-la-Peña, C.; Loyola-Vargas, V.M. Biotic interactions in the rhizosphere: A diverse cooperative enterprise for plant productivity. Plant Physiol. 2014, 166, 701–719. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistic Division; FAO: Rome, Italy, 2015. [Google Scholar]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: Drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2016, 66, 35–42. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Rhizobacteria Species | Contributions towards Sustainable Agriculture | References |
|---|---|---|
| Azospirillum amazonense | Enhanced grain yield by increasing dry matter, panicle number and nitrogen content at maturation. | [118] |
| Pseudomonas aeruginosa | Enhanced the remediation capacity of broad bean plants cultivated in soil environment containing oil contaminants. It also helps to control plant pathogens. | [4,119,120] |
| Serratia liquefaciens | Enhanced the remediation capacity of broad bean plants cultivated in soil environment containing oil contaminants. | [2] |
| Bradyrhizobium spp. | Improved nodulation in leguminous plants as well as shoot and root growth. They also enhance plants resistance to drought and production of indole-3-acetic acid | [2,116,117,121] |
| Azospirillum spp. | Enhanced N content in Vicia sativa. | [122] |
| Rhizobium spp. | Enhanced significantly the height, pod number and length as well as seed weight of Vigna mungo and Vigna radiate. | [123] |
| Bacillus spp. | Help plants to develop resistance against pathogens and pest. | [4,120] |
| Sinorhizobium meliloti | Improved biomass diversity in black madic plant that was subjected to copper stress. | [116,124] |
| Rhizobium RL9 | Increased lentil plant development, nitrogen content, seed protein content and seed produced under heavy metal stressed environment. | [115] |
| Rhizobium MRPI | Promoted nodule formation, leghaemoglobin concentration, seed protein and seed harvest in pea plant. | [125] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Igiehon, N.O.; Babalola, O.O. Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture. Int. J. Environ. Res. Public Health 2018, 15, 574. https://doi.org/10.3390/ijerph15040574
Igiehon NO, Babalola OO. Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture. International Journal of Environmental Research and Public Health. 2018; 15(4):574. https://doi.org/10.3390/ijerph15040574
Chicago/Turabian StyleIgiehon, Nicholas Ozede, and Olubukola Oluranti Babalola. 2018. "Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture" International Journal of Environmental Research and Public Health 15, no. 4: 574. https://doi.org/10.3390/ijerph15040574
APA StyleIgiehon, N. O., & Babalola, O. O. (2018). Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture. International Journal of Environmental Research and Public Health, 15(4), 574. https://doi.org/10.3390/ijerph15040574