Geospatial Analysis and Seasonal Distribution of West Nile Virus Vectors (Diptera: Culicidae) in Southern Ontario, Canada


Abstract
:1. Introduction
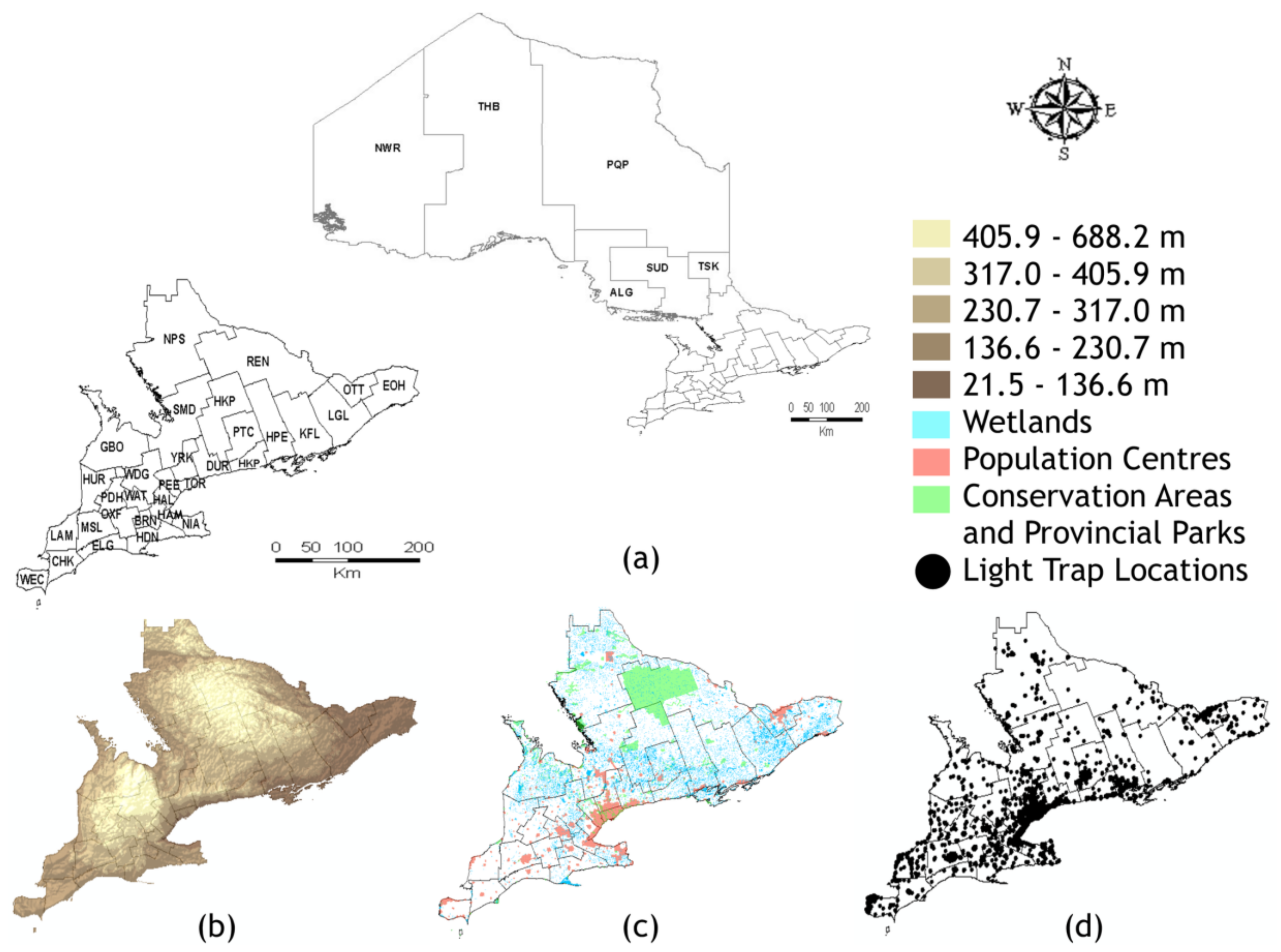
2. Materials and Methods
3. Results
3.1. Analyses Including All 36 HUs
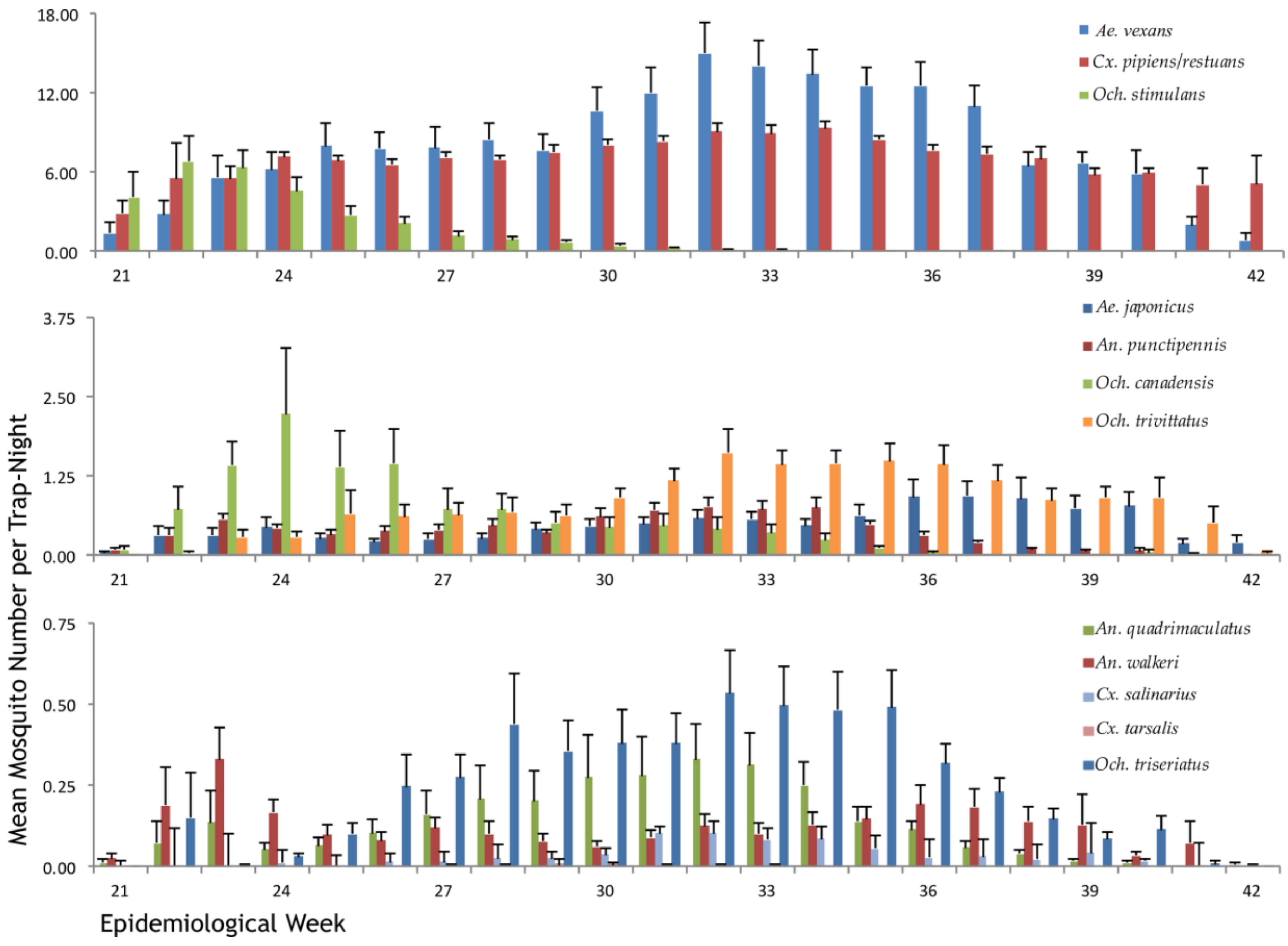
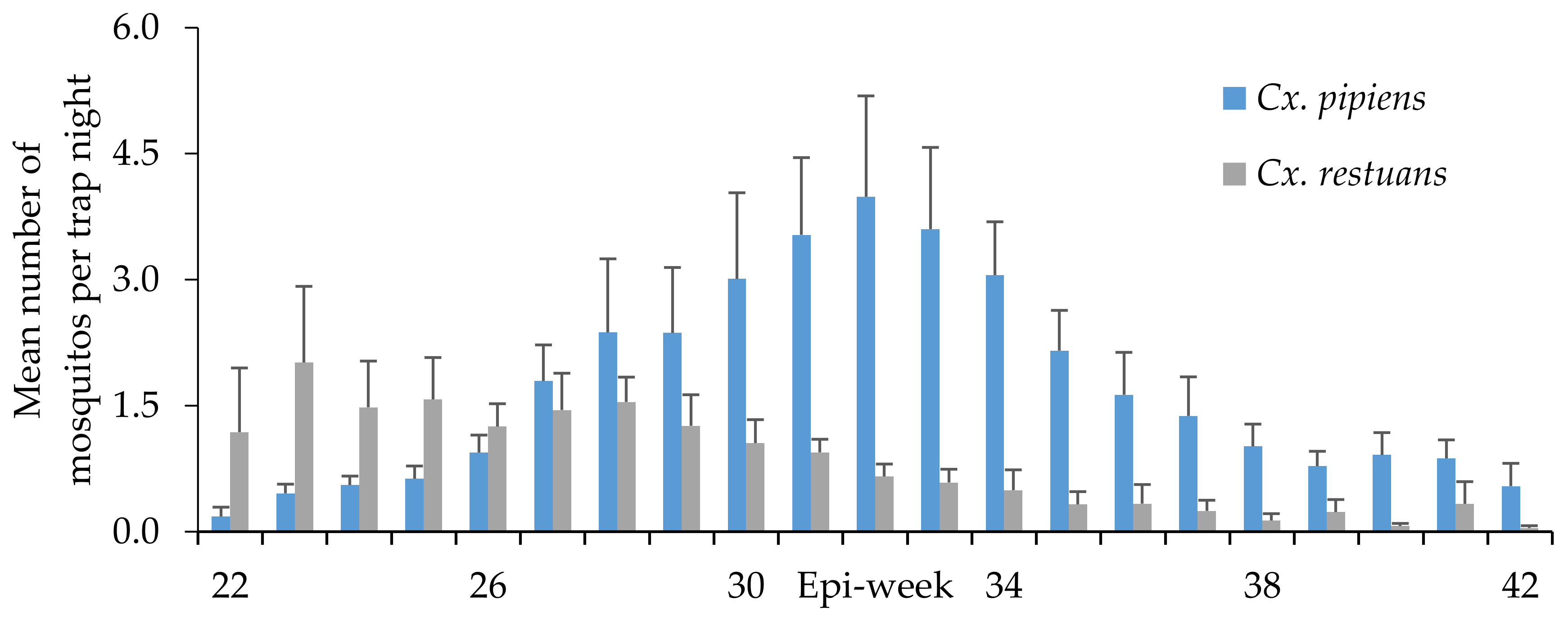
Seasonal Distribution of WNV Vectors in Ontario
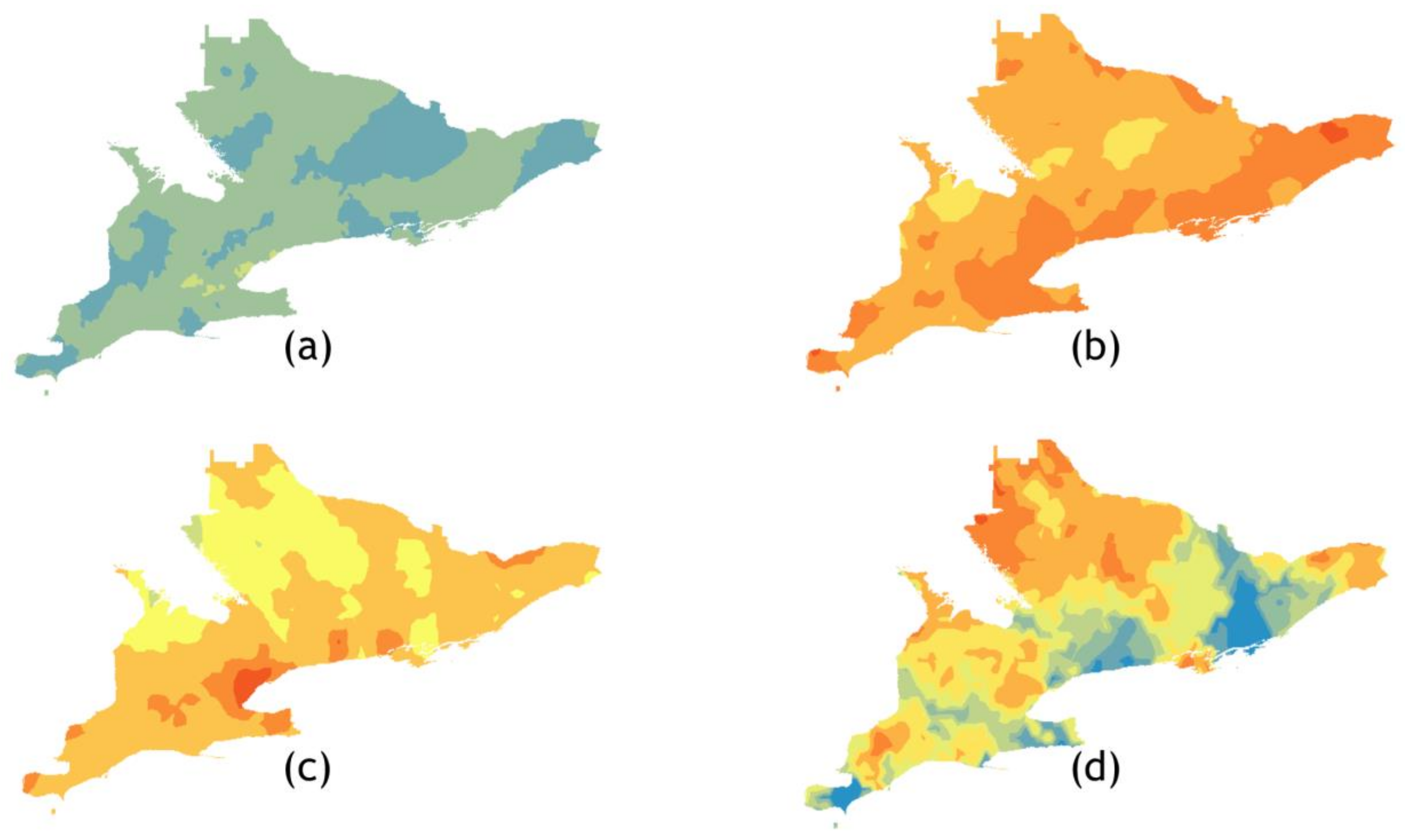
3.2. Geospatial Analyses of the 30 Southern Ontario HUs
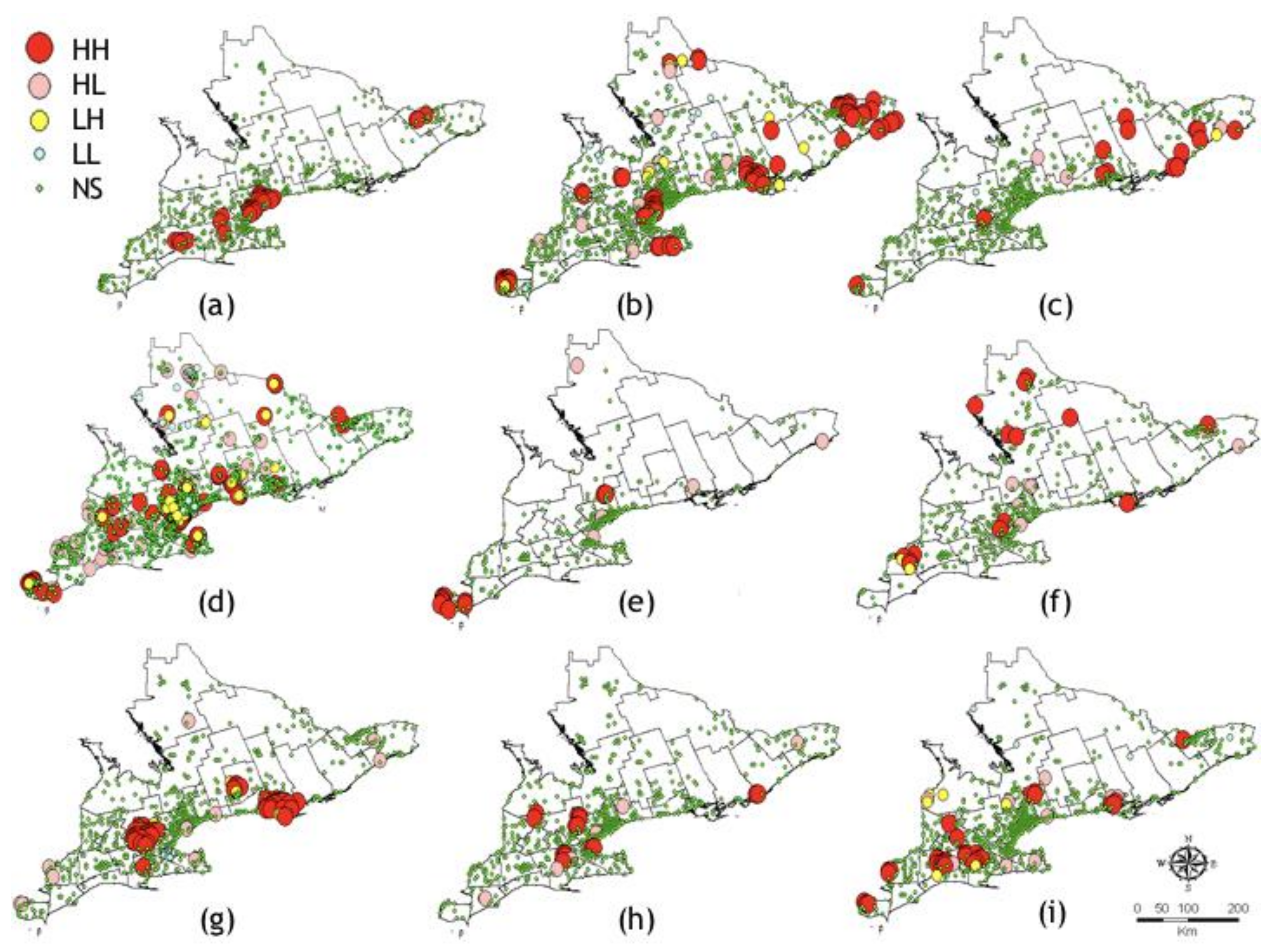
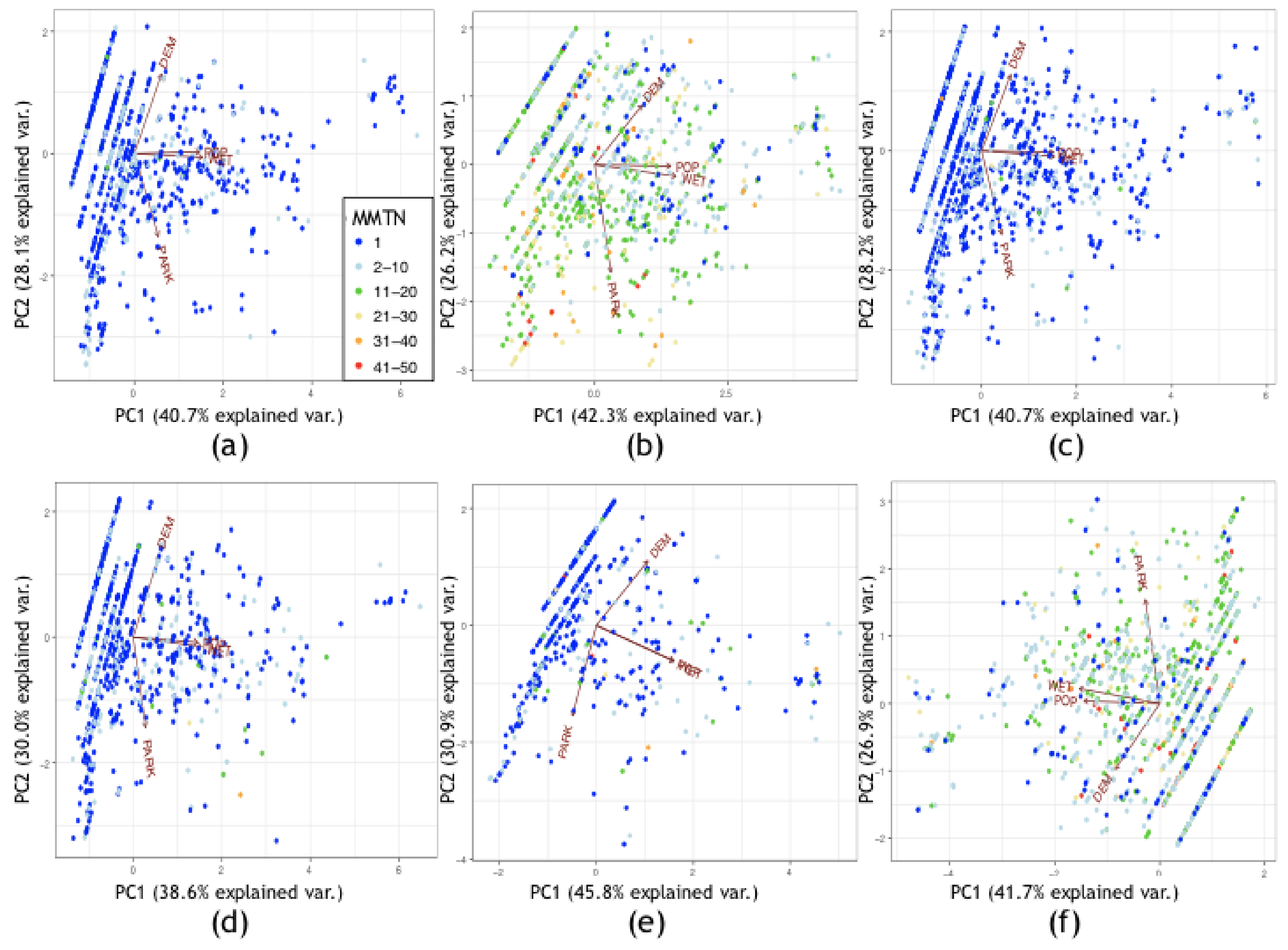
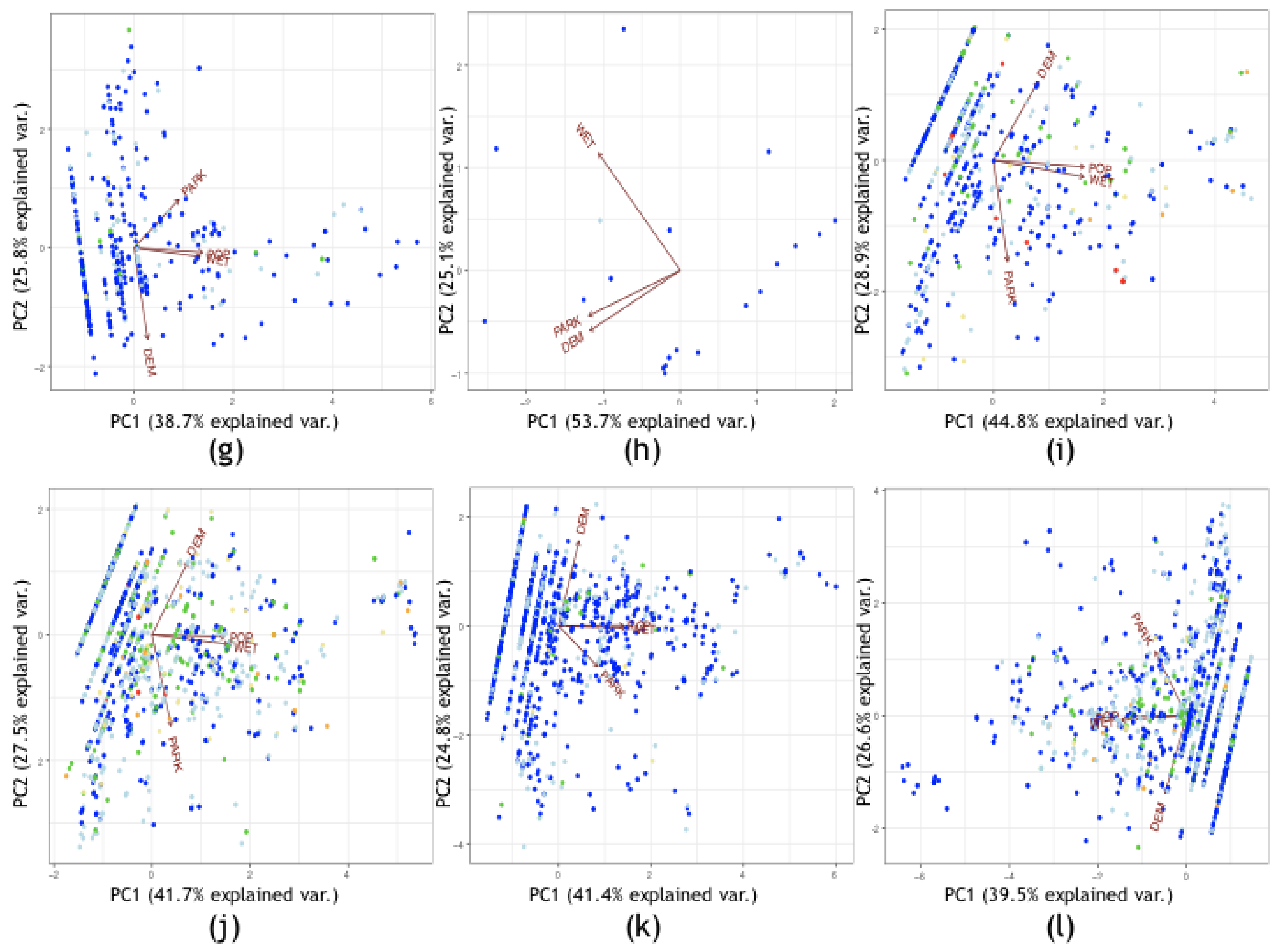
3.2.1. Exploratory Data Analysis
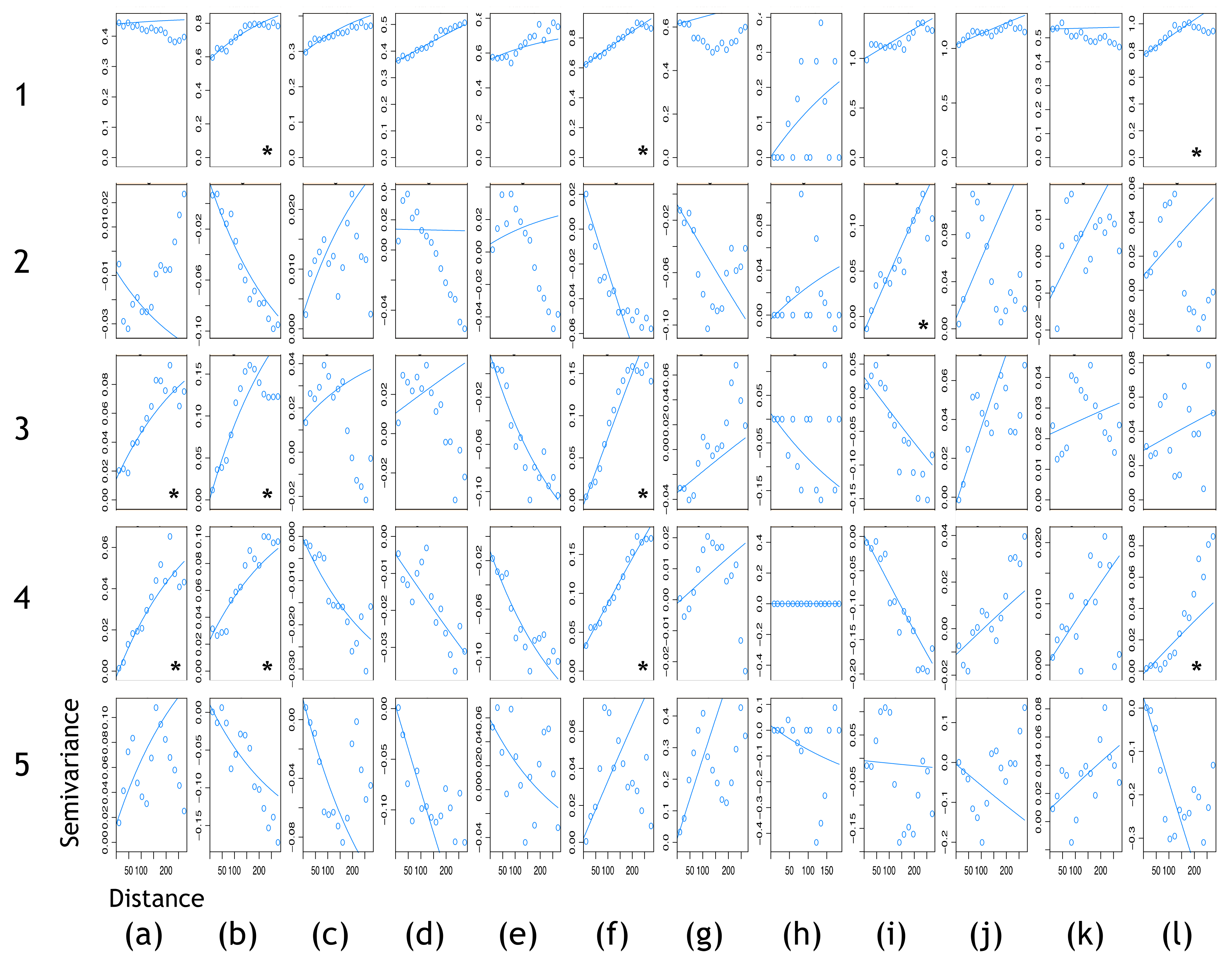
3.2.2. Kriging/Co-Kriging
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HU | ‘02 | ‘03 | ‘04 | ‘05 | ‘06 | ‘07 | ‘08 | ‘09 | ‘10 | ‘11 | ‘12 | ‘13 | ‘14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Algoma District | 8 | 103 | 210 | 136 | 138 | 161 | 123 | 103 | 121 | 61 | 52 | 25 | 45 |
| Brant County | 76 | 47 | 139 | 120 | 117 | 105 | 139 | 150 | 123 | 117 | 98 | 143 | 114 |
| Chatham-Kent | 74 | 78 | 166 | 121 | 143 | 155 | 136 | 127 | 121 | 122 | 103 | 137 | 106 |
| Durham Region | 103 | 77 | 222 | 191 | 178 | 177 | 228 | 159 | 145 | 151 | 152 | 162 | 132 |
| Elgin-St. Thomas | 21 | 32 | 61 | 57 | 51 | 49 | 69 | 23 | 19 | 13 | 18 | 15 | 12 |
| Eastern Ontario | 64 | 46 | 100 | 119 | 104 | 115 | 119 | 76 | 66 | 73 | 70 | 92 | 92 |
| Grey Bruce 1 | 0 | 41 | 89 | 89 | 36 | 8 | 1 | 2 | 0 | 9 | 1 | 0 | 0 |
| Halton Region | 160 | 336 | 302 | 354 | 318 | 335 | 347 | 192 | 185 | 175 | 228 | 212 | 208 |
| City of Hamilton | 104 | 138 | 611 | 572 | 504 | 575 | 530 | 488 | 516 | 463 | 344 | 371 | 418 |
| Haldimand-Norfolk | 35 | 43 | 214 | 227 | 211 | 255 | 230 | 185 | 100 | 37 | 20 | 41 | 33 |
| Haliburton-Kawartha-Pine Ridge | 32 | 86 | 74 | 343 | 299 | 292 | 316 | 123 | 98 | 36 | 117 | 136 | 166 |
| Hastings and Prince Edward Counties | 64 | 42 | 174 | 173 | 185 | 190 | 87 | 63 | 58 | 131 | 88 | 81 | 134 |
| Huron County | 0 | 85 | 94 | 114 | 128 | 122 | 131 | 130 | 122 | 103 | 53 | 74 | 106 |
| Kingston-Frontenac and Lennox & Addington | 0 | 34 | 35 | 33 | 33 | 25 | 27 | 24 | 63 | 46 | 50 | 41 | 53 |
| Lambton County | 40 | 66 | 122 | 158 | 185 | 168 | 179 | 124 | 134 | 128 | 131 | 126 | 115 |
| Leegs-Grenville and Lanark District | 63 | 41 | 105 | 103 | 100 | 98 | 98 | 44 | 39 | 29 | 45 | 138 | 100 |
| Middlesex-London | 172 | 175 | 221 | 209 | 241 | 368 | 400 | 277 | 360 | 379 | 327 | 351 | 378 |
| Niagara region | 115 | 149 | 224 | 189 | 185 | 162 | 151 | 246 | 274 | 308 | 295 | 364 | 350 |
| North Bay Perry Sound District | 5 | 89 | 124 | 120 | 127 | 131 | 131 | 71 | 81 | 66 | 69 | 47 | 91 |
| Northwestern | 0 | 14 | 74 | 56 | 105 | 90 | 61 | 46 | 44 | 21 | 19 | 26 | 61 |
| City of Ottawa | 10 | 417 | 481 | 532 | 462 | 419 | 436 | 419 | 338 | 307 | 299 | 427 | 476 |
| Oxford County | 114 | 49 | 144 | 137 | 97 | 122 | 125 | 115 | 114 | 123 | 121 | 148 | 132 |
| Perth District | 82 | 34 | 134 | 125 | 118 | 118 | 130 | 55 | 68 | 48 | 49 | 78 | 84 |
| Peel Region | 225 | 430 | 482 | 480 | 493 | 442 | 474 | 452 | 473 | 464 | 472 | 495 | 435 |
| Porcupine | 0 | 62 | 131 | 93 | 147 | 155 | 138 | 125 | 54 | 25 | 39 | 126 | 110 |
| Peterborough County-City | 24 | 28 | 70 | 75 | 73 | 63 | 76 | 50 | 46 | 54 | 67 | 78 | 71 |
| Renfrew County and District | 21 | 42 | 68 | 66 | 73 | 74 | 72 | 67 | 41 | 30 | 36 | 46 | 43 |
| Simcoe Muskoka District | 92 | 137 | 261 | 272 | 301 | 299 | 270 | 131 | 200 | 172 | 177 | 128 | 181 |
| Sudbury and District | 0 | 96 | 213 | 183 | 220 | 354 | 349 | 252 | 150 | 54 | 111 | 108 | 72 |
| Thunder Bay District | 0 | 60 | 162 | 154 | 36 | 73 | 47 | 18 | 32 | 38 | 17 | 26 | 16 |
| City of Toronto | 310 | 627 | 965 | 798 | 673 | 515 | 651 | 380 | 630 | 601 | 678 | 665 | 641 |
| Timiskaming | 0 | 63 | 51 | 57 | 41 | 63 | 63 | 63 | 47 | 34 | 27 | 36 | 77 |
| Region of Waterloo | 193 | 45 | 174 | 174 | 228 | 217 | 278 | 225 | 221 | 216 | 199 | 208 | 195 |
| Wellington-Dufferin-Guelph | 52 | 41 | 151 | 150 | 150 | 147 | 147 | 148 | 130 | 106 | 87 | 159 | 109 |
| Windsor-Essex County | 155 | 185 | 169 | 189 | 249 | 387 | 333 | 347 | 224 | 218 | 231 | 195 | 171 |
| York Region | 137 | 253 | 416 | 544 | 574 | 561 | 409 | 361 | 422 | 324 | 345 | 510 | 497 |
References
- Staples, J.E.; Shankar, M.B.; Sejvar, J.J.; Meltzer, M.I.; Fischer, M. Initial and long-term costs of patients hospitalized with West Nile virus disease. Am. J. Trop. Med. Hyg. 2014, 90, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Centres for Disease Control and Prevention. West Nile Virus Disease Cases Reported to CDC by State, 1999–2013; CDC Centres for Disease Control and Prevention: Atlanta, GA, USA, 2014.
- Public Health Agency of Canada. Surveillance of West Nile Virus; PHAC Public Health Agency of Canada: Ottawa, ON, Canada, 2015.
- Giordano, B.V.; Gasparotto, A.; Hunter, F.F. A checklist of the 67 mosquito species of Ontario, Canada. J. Am. Mosq. Control Assoc. 2015, 31, 101–103. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector Borne Zoonotic Dis. 2004, 2, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Popov, V.; Wen, J.; Han, V.; Higgs, S. Ultrastructure study of West Nile virus pathogenesis in Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2005, 3, 429–444. [Google Scholar] [CrossRef]
- Andreadis, T.G.; Anderson, J.F.; Vossbrinck, C.R. Mosquito surveillance for West Nile virus in Connecticut, 2000: Isolation of Culex pipiens, Cx. restuans, Cx. salinarius, and Culiseta melanura. Emerg. Infect. Dis. 2001, 7, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Kulasekera, V.L.; Kramer, L.; Nasci, R.S.; Mostashari, F.; Cherry, B.; Trock, S.C.; Glaser, C.; Miller, J.R. West Nile virus infection in mosquitoes, birds, horses, and humans, Staten Island, New York, 2000. Emerg. Infect. Dis. 2001, 7, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’Guinn, M.L.; Andreadis, T.G. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.; O’Guinn, M.; Dohm, D.; Jones, J. Vector competency of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Drebot, M.A.; Lindsay, R.; Barker, I.K.; Buck, P.A.; Fearon, M.; Hunter, F.; Sockett, P.; Artsob, H. West Nile virus surveillance and diagnostics: A Canadian perspective. Can. J. Infect. Dis. Med. 2003, 14, 105–114. [Google Scholar] [CrossRef]
- Public Health Ontario. Guide for Public Health Units: Considerations for Adult Mosquito Control; PHO Public Health Ontario: Toronto, ON, Canada, 2013. [Google Scholar]
- Statistics Canada. The Changing Landscape of Canadian Metropolitan Areas. Statistics Canada, No. 16201X; Statistics Canada: Ottawa, ON, Canada, 2016.
- Anderson, S.L.; Richards, S.L.; Tabachnick, W.J.; Smartt, C.T. Effects of West Nile virus dose and extrinsic incubation temperature on temporal progression of vector competence in Culex pipiens quinquefasciatus. J. Am. Mosq. Control Assoc. 2010, 26, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.M.; Dang, P.T.; Ellis, R.A. The Mosquitoes of Canada (Diptera: Culicidae): The Insects and Arachnids of Canada, Part VI; Agriculture Canada: Ottawa, ON, Canada, 1979.
- Darsie, R.F.; Ward, R.A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University Press of Florida: Gainesville, FL, USA, 2005. [Google Scholar]
- Statistics Canada. Health Region Boundary Files. Health Indicators, 82-221-X.; No. 1; Statistics Canada: Ottawa, ON, Canada, 2015.
- Ontario Ministry of Natural Resources. Provincial Digital Elevation Model; Ontario Ministry of Natural Resources: Peterborough, ON, Canada, 2012.
- Statistics Canada. Population Centres, Digital Boundary File- 2016 Census; Statistics Canada: Ottawa, ON, Canada, 2016.
- Statistics Canada. 2011 Census Boundary File; Statistics Canada: Ottawa, ON, Canada, 2011.
- Ontario Ministry of Natural Resources. Wetland Unit. 2011. Available online: http://geo.scholarsportal.info/#r/details/_uri@=2555853800&_add:true_nozoom:true (accessed on 10 April 2017).
- Ontario Ministry of Natural Resources. Land Information Ontario. 2014. Available online: https://www.ontario.ca/page/land-information-ontario (accessed on 10 April 2017).
- Thielman, A.; Hunter, F. A Photographic Key to Adult Female Mosquito Species of Canada (Diptera: Culicidae). Can. J. Arthropod Identif. 2007, 4, 1–117. [Google Scholar]
- Public Health Ontario. Vector-Borne Disease Surveillance Reports; PHO Public Health Ontario: Toronto, ON, Canada, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Service, M.W. Mosquito (Diptera: Culicidae) dispersal—The long and short of it. J. Med. Entomol. 1997, 3, 579–588. [Google Scholar] [CrossRef]
- Chun, Y.; Griffith, D.A. Spatial Statistics and Geostatistics: Theory and Applications for Geographic Information Science and Technology; Sage Publications Ltd.: Thousands Oaks, CA, USA, 2013. [Google Scholar]
- Gräler, B.; Pebesma, E.; Heuvelink, G. Spatio-Temporal Interpolation using gstat. R J. 2016, 8, 204–218. [Google Scholar]
- Pebesma, E.J. Multivariable geostatistics in R: The gstat package. Comput. Geosci. 2004, 30, 683–691. [Google Scholar] [CrossRef]
- Pardo-Igúzquiza, E.; Dowd, P.A. Empirical maximum likelihood kriging: The general case. Math. Geol. 2005, 37, 477–492. [Google Scholar] [CrossRef]
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot. R Package Version 0.55. 2011. Available online: http://github.com/vqv/ggbiplot (accessed on 5 December 2017).
- Reeves, W.; Korecki, J. Ochlerotatus japonicus (Theobald) (Diptera: Culicidae) a new invasive mosquito for Georgia and South Carolina. Proc. Entomol. Soc. Wash. 2004, 106, 233–234. [Google Scholar]
- Thielman, A.; Hunter, F. Establishment of Ochlerotatus japonicus (Diptera: Culicidae) in Ontario, Canada. J. Med. Entomol. 2006, 43, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Belton, P.; McMahon, S.; Hart, M.; McCann, S.; Azevedo, D.; Hurteau, L. The first record of Aedes (Hulecoeteomyia) japonicus (Diptera: Culicidae) and its establishment in western Canada. J. Med. Entomol. 2016, 53, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Fielden, M.A.; Chaulk, A.C.; Bassett, K.; Wiersma, Y.F.; Erbland, M.; Whitney, H.; Chapman, T.W. Aedes japonicus japonicus (Diptera: Culicidae) arrives at the most easterly point in North America. Can. Entomol. 2015, 147, 737–740. [Google Scholar] [CrossRef]
- Ledesma, N.; Harrington, L. Mosquito vectors of dog heartworm in the United States: Vector status and factors influencing transmission efficiency. Top. Companion. Anim. Med. 2011, 26, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Helbing, C.M.; Moorhead, D.L.; Mitchell, L. Population dynamics of Culex restuans and Culex pipiens (Diptera: Culicidae) related to climatic factors in northwest Ohio. Environ. Entomol. 2015, 44, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLoS Biol. 2006, 4, e82. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.; Hunter, F.F. Culex pipiens (Culicidae) is attracted to humans in southern Ontario, but will it serve as a bridge vector of West Nile virus? Can. Entomol. 2012, 144, 667–671. [Google Scholar] [CrossRef]
- Giordano, B.V.; Kaur, S.; Hunter, F.F. West Nile virus in Ontario, Canada: A twelve-year analysis of human case prevalence, mosquito surveillance, and climate data. PLoS ONE 2017, 12, e0183568. [Google Scholar] [CrossRef] [PubMed]
- Roth, D.; Henry, B.; Mak, S.; Fraser, M.; Taylor, M.; Li, M.; Cooper, K.; Furnell, A.; Wong, Q.; Morshed, M.; et al. West Nile virus range expansion into British Columbia. Emerg. Infect. Dis. 2010, 16, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.; Lothrop, H.; Chiles, R.; Madon, M.; Cossen, C.; Woods, L.; Husted, S.; Kramer, V.; Edman, J. West Nile virus in California. Emerg. Infect. Dis. 2004, 10, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, M.; Turell, M.J.; Lindsay, L.R. Potential for Canadian mosquitoes to transmit Rift Valley fever virus. J. Am. Mosq. Control Assoc. 2011, 27, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Andreadis, T.G. Eastern equine encephalitis virus in mosquitoes and their role as bridge vectors. Emerg. Infect. Dis. 2010, 1, 1869–1874. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.; Hardy, J.; Reisen, W.; Milby, M.M. Potential effect of global warming on mosquito-borne arboviruses. J. Med. Entomol. 1994, 31, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ogden, N.; Zhu, H. The impact of weather conditions on Culex pipiens and Culex restuans (Diptera: Culicidae) abundance: A case study in Peel region. J. Med. Entomol. 2011, 48, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Diuk-Wasser, M.A.; Brown, H.E.; Andreadis, T.G.; Fish, D. Modeling the spatial distribution of mosquito vectors for West Nile virus in Connecticut, USA. Vector Borne Zoonotic Dis. 2006, 6, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Yoo, E.H.; Chen, D.; Diao, C.; Russell, C. The effects of weather and environmental factors on West Nile virus mosquito abundance in greater Toronto area. Earth Interact. 2016, 20, 1–22. [Google Scholar] [CrossRef]







| Species | 5 km | 10 km | 15 km | 20 km |
|---|---|---|---|---|
| Ae. japonicus | 0.03 (0.386) | 0.04 (0.109) | 0.06 (0.010) 1 | 0.05 (0.014) |
| Ae. vexans | 0.20 (<0.001) 1 | 0.20 (<0.001) 1 | 0.17 (<0.001) 1 | 0.17 (<0.001) 1 |
| An. punctipennis | 0.04 (0.207) | 0.02 (0.506) | 0.03 (0.521) | 0.02 (0.351) |
| An. quadrimaculatus | 0.04 (0.241) | 0.06 (0.040) 1 | 0.04 (0.108) | 0.07 (<0.001) 1 |
| An. walkeri | 0.00 (0.992) | 0.04 (0.315) | 0.04 (0.324) | 0.03 (0.344) |
| Cx. pipiens/restuans | 0.06 (0.030) 1 | 0.02 (0.389) | 0.04 (0.058) 1 | 0.03 (0.131) |
| Cx. salinarius | 0.10 (0.042) 1 | 0.08 (0.057) 1 | 0.05 (0.149) | 0.05 (0.131) |
| Cx. tarsalis | −0.03 (0.401) | −0.03 (0.437) | −0.02 (0.423) | −0.02 (0.280) |
| Och. canadensis | 0.07 (0.092) 1 | 0.09 (0.018) 1 | 0.11 (<0.001) 1 | 0.13 (<0.001) 1 |
| Och. stimulans | 0.12 (<0.001) 1 | 0.11 (<0.001) 1 | 0.11 (<0.001) 1 | 0.13 (<0.001) 1 |
| Och. triseriatus | 0.10 (0.005) 1 | 0.06 (0.075) 1 | 0.02 (0.420) | 0.02 (0.250) |
| Och. trivittatus | 0.11 (0.002) 1 | 0.13 (<0.001) 1 | 0.12 (<0.001) 1 | 0.10 (<0.001) 1 |
| Species | Variable(s) | Kriging Type | Model | RMSS | MS | RMS | ASE |
|---|---|---|---|---|---|---|---|
| Ae. japonicus | MMTN, POP | Universal | Stable | 0.872 | −0.005 | 2.153 | 2.470 |
| MMTN, WET 1 | Simple | Stable | 0.952 | 0.002 | 2.175 | 2.325 | |
| Ae. vexans | MMTN | Simple | Stable | 0.984 | 0.019 | 8.521 | 8.645 |
| MMTN, POP 1 | Simple | Stable | 0.995 | 0.001 | 8.556 | 8.686 | |
| MMTN, WET | Simple | Stable | 1.018 | −0.009 | 8.652 | 8.633 | |
| Cx. pipiens/restuans | MMTN | Universal | Stable | 0.940 | −0.001 | 9.880 | 10.629 |
| MMTN, POP | Simple | Spherical | 1.034 | −0.002 | 10.157 | 10.533 | |
| MMTN, WET 1 | Simple | Gaussian | 1.012 | −0.001 | 10.201 | 10.596 | |
| Och. canadensis | MMTN, DEM 1 | Universal | Stable | 0.942 | 0.003 | 9.050 | 9.644 |
| Och. trivittatus | MMTN | Universal | Stable | 1.121 | −0.006 | 6.162 | 5.504 |
| MMTN, POP 1 | Universal | Gaussian | 1.121 | −0.006 | 6.162 | 5.504 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, B.V.; Turner, K.W.; Hunter, F.F. Geospatial Analysis and Seasonal Distribution of West Nile Virus Vectors (Diptera: Culicidae) in Southern Ontario, Canada. Int. J. Environ. Res. Public Health 2018, 15, 614. https://doi.org/10.3390/ijerph15040614
Giordano BV, Turner KW, Hunter FF. Geospatial Analysis and Seasonal Distribution of West Nile Virus Vectors (Diptera: Culicidae) in Southern Ontario, Canada. International Journal of Environmental Research and Public Health. 2018; 15(4):614. https://doi.org/10.3390/ijerph15040614
Chicago/Turabian StyleGiordano, Bryan V., Kevin W. Turner, and Fiona F. Hunter. 2018. "Geospatial Analysis and Seasonal Distribution of West Nile Virus Vectors (Diptera: Culicidae) in Southern Ontario, Canada" International Journal of Environmental Research and Public Health 15, no. 4: 614. https://doi.org/10.3390/ijerph15040614
APA StyleGiordano, B. V., Turner, K. W., & Hunter, F. F. (2018). Geospatial Analysis and Seasonal Distribution of West Nile Virus Vectors (Diptera: Culicidae) in Southern Ontario, Canada. International Journal of Environmental Research and Public Health, 15(4), 614. https://doi.org/10.3390/ijerph15040614