Multisensory Integration Strategy for Modality-Specific Loss of Inhibition Control in Older Adults


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Stimulus Characteristics
2.2.1. Auditory Stimuli
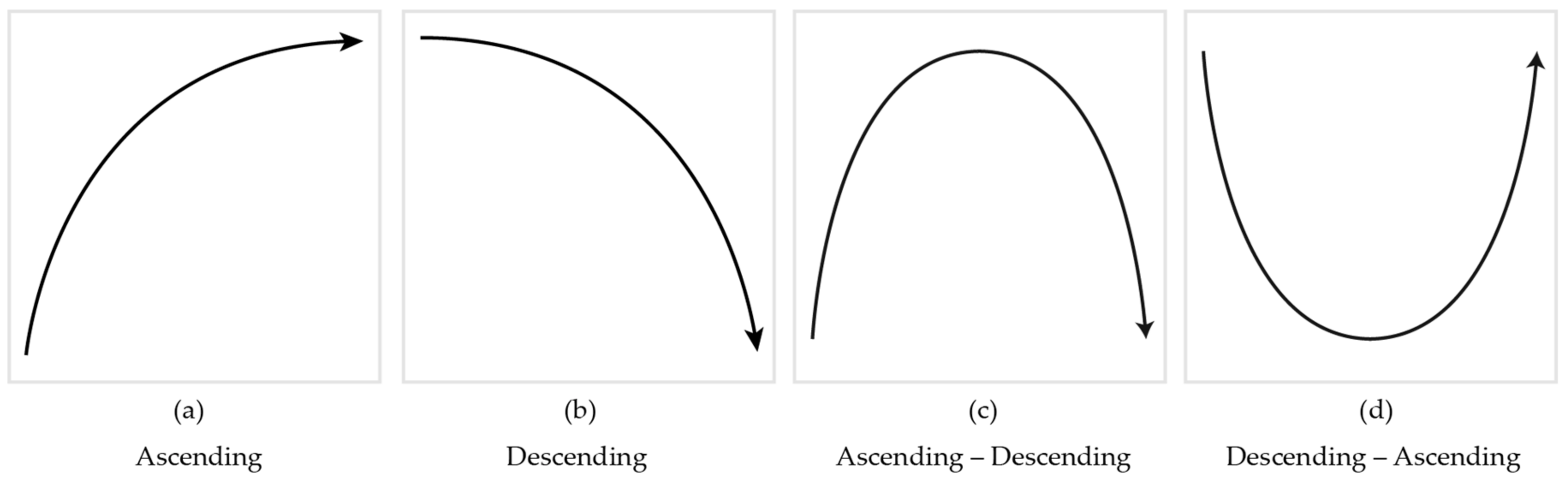
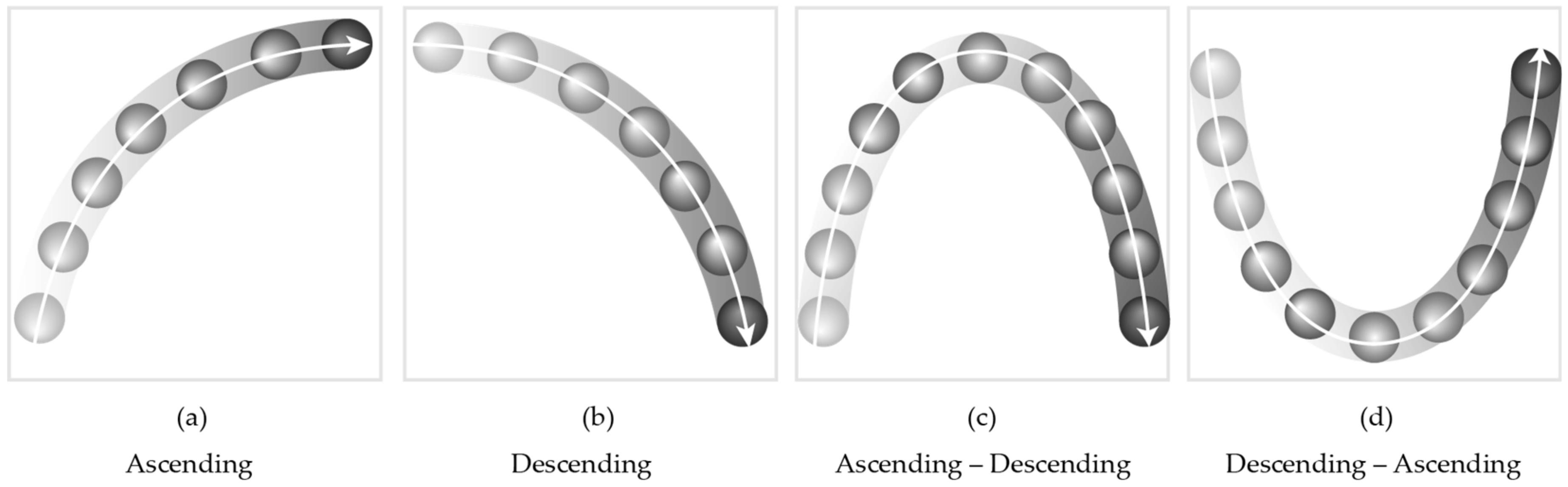
2.2.2. Visual Stimuli
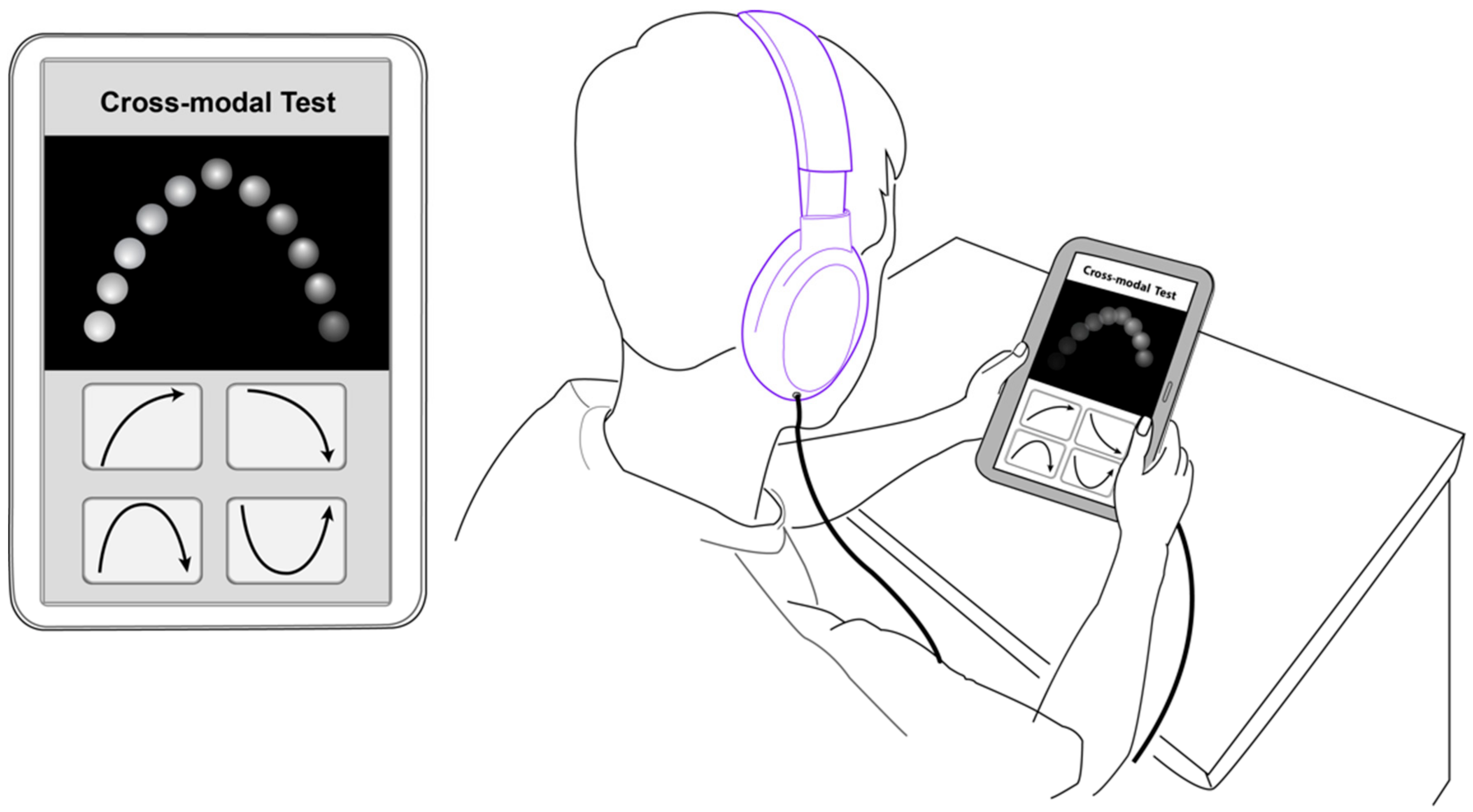
2.3. Audio-Visual Contour Identification Task
2.4. Procedure
2.5. Data Analysis
3. Results
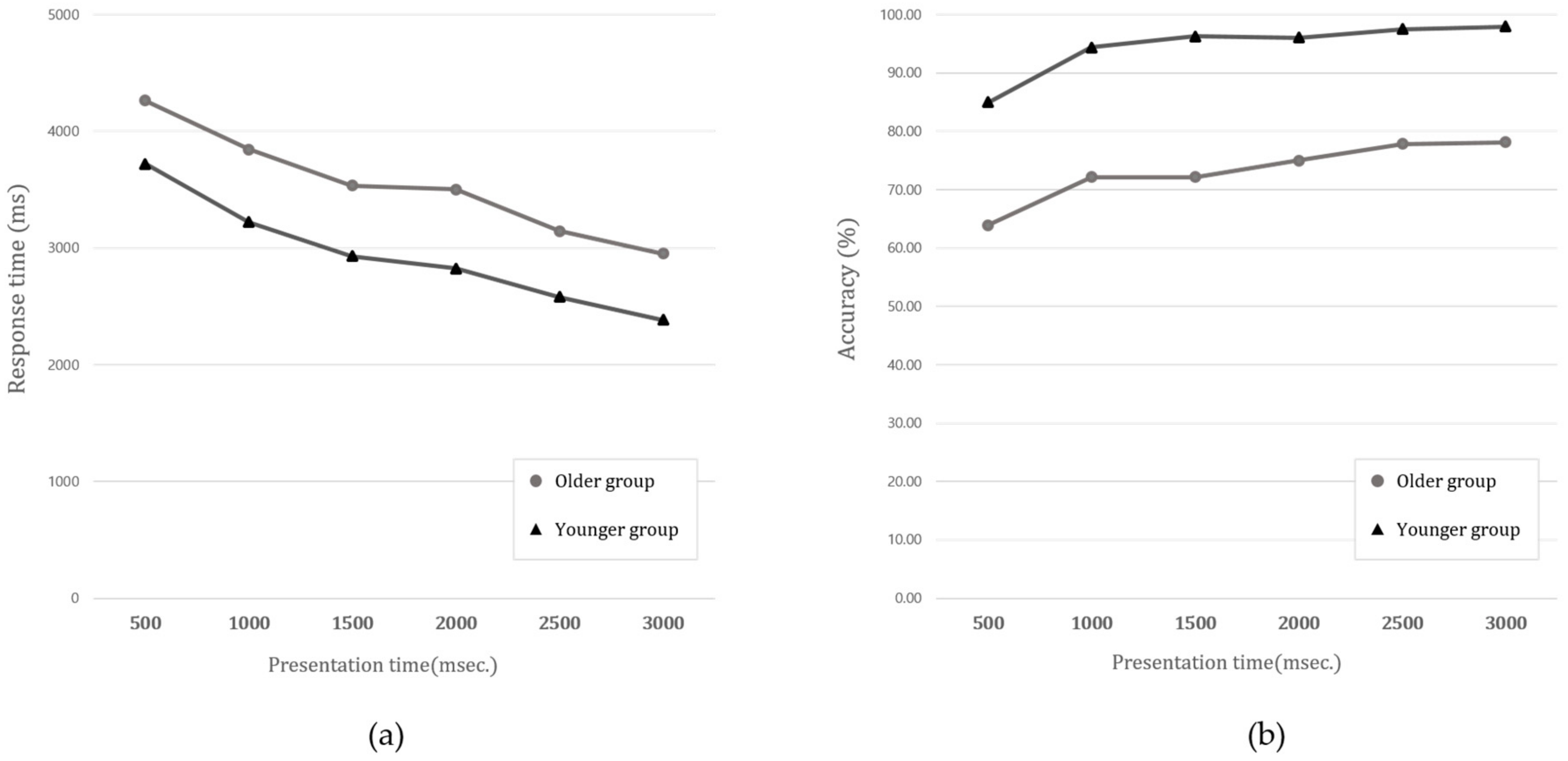
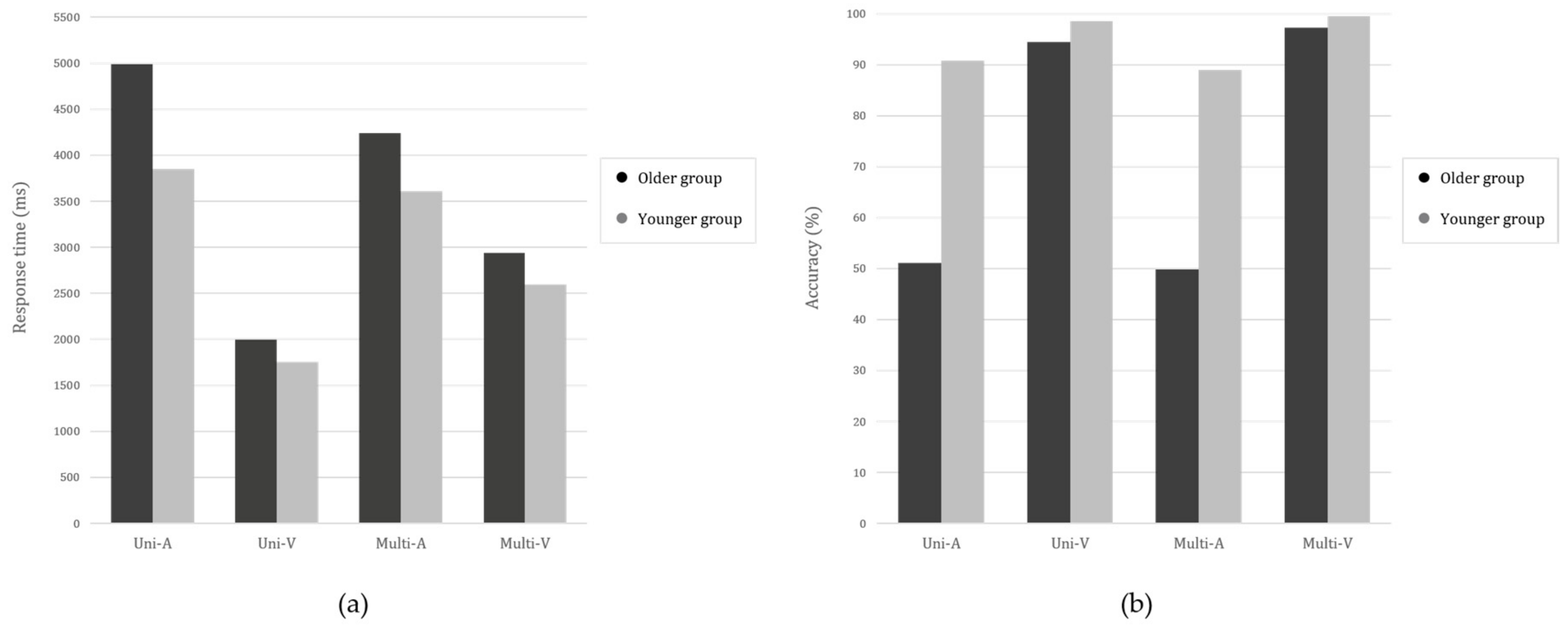
3.1. Comparing the Performance of the Four Audio-Visual Contour Identification Tasks
3.2. Examining the Role of Unattended Stimuli in Multisensory Integration: Reference or Distraction?
4. Discussion
4.1. Vision is Dominant, So Auditory Distraction Is Less Prominent in the Visual Target Identification
4.2. Visual Distraction as a Negative Reference for Auditory Target Identification
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Glisky, E.L. Changes in cognitive function in human aging. In Brain Aging: Models, Methods, and Mechanisms; CRC Press: Boca Raton, FL, USA, 2007; pp. 3–20. [Google Scholar]
- Alain, C.; Quan, J.; McDonald, K.; Van Roon, P. Noise-induced increase in human auditory evoked neuromagnetic fields. Eur. J. Neurosci. 2009, 30, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Ostroff, J.M.; McDonald, K.L.; Schneider, B.A.; Alain, C. Aging and the processing of sound duration in human auditory cortex. Hearing Res. 2003, 181, 1–7. [Google Scholar] [CrossRef]
- Skeel, R.L.; Schutte, C.; van Voorst, W.; Nagra, A. The relationship between visual contrast sensitivity and neuropsychological performance in a healthy elderly sample. J. Clin. Exp. Neuropsychol. 2006, 28, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Alain, C. Age-related changes in neural activity associated with concurrent vowel segregation. Cognit. Brain Res. 2005, 24, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.C.; Jin, J.X.; Gunasekera, G.M.; Abel, R.; Lee, E.R.; Dhar, S. Aging and cortical mechanisms of speech perception in noise. Neuropsychologia 2009, 47, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, M.J.; Murphy, D.R.; Van Gerven, P.W. The role of sensory modality in age-related distraction: A critical review and a renewed view. Psychol. Bull. 2010, 136, 975–1022. [Google Scholar] [CrossRef] [PubMed]
- Hasher, L.; Lustig, C.; Zacks, R. Inhibitory mechanisms and the control of attention. In Variation in Working Memory; Oxford University Press: New York, NY, USA, 2007; Volume 19, pp. 227–249. [Google Scholar]
- Lustig, C.; Hasher, L. Interference. In Encyclopedia of Aging, 3rd ed.; Maddox, G., Ed.; Springer: New York, NY, USA, 2001; pp. 553–555. [Google Scholar]
- Stein, B.E.; Stanford, T.R.; Ramachandran, R.; Perrault, T.J.; Rowland, B.A. Challenges in quantifying multisensory integration: Alternative criteria, models, and inverse effectiveness. Exp. Brain Res. 2009, 198, 113. [Google Scholar] [CrossRef] [PubMed]
- Foxe, J.J.; Schroeder, C.E. The case for feedforward multisensory convergence during early cortical processing. Neuroreport 2005, 16, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Giard, M.H.; Peronnet, F. Auditory-visual integration during multimodal object recognition in humans: A behavioral and electrophysiological study. J. Cognit. Neurosci. 1999, 11, 473–490. [Google Scholar] [CrossRef]
- Molholm, S.; Ritter, W.; Murray, M.M.; Javitt, D.C.; Schroeder, C.E.; Foxe, J.J. Multisensory auditory–visual interactions during early sensory processing in humans: A high-density electrical mapping study. Cognit. Brain Res. 2002, 14, 115–128. [Google Scholar] [CrossRef]
- Cappe, C.; Barone, P. Heteromodal connections supporting multisensory integration at low levels of cortical processing in the monkey. Eur. J. Neurosci. 2005, 22, 2886–2902. [Google Scholar] [CrossRef] [PubMed]
- Falchier, A.; Clavagnier, S.; Barone, P.; Kennedy, H. Anatomical evidence of multimodal integration in primate striate cortex. J. Neurosci. 2002, 22, 5749–5759. [Google Scholar] [PubMed]
- Rockland, K.S.; Ojima, H. Multisensory convergence in calcarine visual areas in macaque monkey. Int. J. Psychophysiol. 2003, 50, 19–26. [Google Scholar] [CrossRef]
- McGurk, H.; MacDonald, J. Hearing lips and seeing voices. Nature 1976, 264, 746–748. [Google Scholar] [CrossRef] [PubMed]
- Freiherr, J.; Lundström, J.N.; Habel, U.; Reetz, K. Multisensory integration mechanisms during aging. Front. Hum. Neurosci. 2013, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hairston, W.D.; Letowski, T.R.; McDowell, K. Task-related suppression of the brainstem frequency following response. PLoS ONE 2013, 8, e55215. [Google Scholar] [CrossRef] [PubMed]
- Andersen, G.J.; Ni, R. Aging and visual processing: Declines in spatial not temporal integration. Vis. Res. 2008, 48, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Bennett, P.J.; Sekuler, R.; Sekuler, A.B. The effects of aging on motion detection and direction identification. Vis. Res. 2007, 47, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Betts, L.R.; Sekuler, A.B.; Bennett, P.J. The effects of aging on orientation discrimination. Vis. Res. 2007, 47, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Del Viva, M.M.; Agostini, R. Visual spatial integration in the elderly. Investig. Ophthalmol. Visual Sci. 2007, 48, 2940–2946. [Google Scholar] [CrossRef] [PubMed]
- Delahunt, P.B.; Hardy, J.L.; Werner, J.S. The effect of senescence on orientation discrimination and mechanism tuning. J. Vis. 2008, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Diaconescu, A.O.; Hasher, L.; McIntosh, A.R. Visual dominance and multisensory integration changes with age. Neuroimage 2013, 65, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Hasher, L.; Zacks, R.T. Aging and a benefit of distractibility. Psychon. Bull. Rev. 2007, 14, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Weeks, J.C.; Hasher, L. The disruptive–and beneficial–effects of distraction on older adults’ cognitive performance. Front. Psychol. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gazzaley, A.; Clapp, W.; Kelley, J.; McEvoy, K.; Knight, R.T.; D’Esposito, M. Age-related top-down suppression deficit in the early stages of cortical visual memory processing. Proc. Natl. Acad. Sci. USA 2008, 105, 13122–13126. [Google Scholar] [CrossRef] [PubMed]
- Gazzaley, A.; Cooney, J.W.; Rissman, J.; D‘esposito, M. Top-down suppression deficit underlies working memory impairment in normal aging. Nat. Neurosci. 2005, 8, 1298. [Google Scholar] [CrossRef] [PubMed]
- Vallesi, A.; Stuss, D.T.; McIntosh, A.R.; Picton, T.W. Age-related differences in processing irrelevant information: Evidence from event-related potentials. Neuropsychologia 2009, 47, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, M.J.; Van Gerven, P.W. Now you see it, now you don‘t: Evidence for age-dependent and age-independent cross-modal distraction. Psychol. Aging 2011, 26, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, M.J.; Murphy, D.R.; Van Gerven, P.W. Making sense of age-related distractibility: The critical role of sensory modality. Acta Psychol. (Amst) 2013, 142, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Čeponienė, R.; Westerfield, M.; Torki, M.; Townsend, J. Modality-specificity of sensory aging in vision and audition: Evidence from event-related potentials. Brain Res. 2008, 1215, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; vom Berg, I.; Rupp, A.; Seidl, U.; Schröder, J.; Roesch-Ely, D.; Kreisel, S.H.; Mundt, C.; Weisbrod, M. P50 gating deficit in alzheimer dementia correlates to frontal neuropsychological function. Neurobiol. Aging 2010, 31, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Lijffijt, M.; Lane, S.D.; Meier, S.L.; Boutros, N.N.; Burroughs, S.; Steinberg, J.L.; Gerard Moeller, F.; Swann, A.C. P50, n100, and p200 sensory gating: Relationships with behavioral inhibition, attention, and working memory. Psychophysiology 2009, 46, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, T.W.; Cheng, F.H.; De Rosa, E. Failing to ignore: Paradoxical neural effects of perceptual load on early attentional selection in normal aging. J. Neurosci. 2010, 30, 14750–14758. [Google Scholar] [CrossRef] [PubMed]
- Lutfi-Proctor, D.A.; Elliott, E.M.; Cowan, N. The role of visual stimuli in cross-modal stroop interference. Psych J. 2014, 3, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Leiva, A.; Parmentier, F.B.; Andres, P. Aging increases distraction by auditory oddballs in visual, but not auditory tasks. Psychol. Res. 2015, 79, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Stothart, G.; Kazanina, N. Auditory perception in the aging brain: The role of inhibition and facilitation in early processing. Neurobiol. Aging 2016, 47, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E. Psychometric validation of a music-based attention assessment: Revised for patients with traumatic brain injury. J. Music Ther. 2013, 50, 66–92. [Google Scholar] [CrossRef] [PubMed]
- Ibm Spss Statistics for Windows; 21.0; IBM Corp, Released; IBM Corp: Armonk, NY, USA, 2012.
- Winer, B.J. Statistical Principles in Experimental Design; American Psychological Association: Washington, DC, USA, 1962. [Google Scholar]
- Kirk, R.E. Experimental Design: Procedures for the Behavioral Sciences; Brooks/Cole: Belmont, CA, USA, 1968. [Google Scholar]
- Williams, R.L.; Stockdale, S.L. High-performing students with low critical thinking skills. J. Gen. Educ. 2003, 52, 199–225. [Google Scholar] [CrossRef]
- Ha, K.S.; Kwon, J.S.; Lyoo, I.K.; Kong, S.W.; Lee, D.W.; Youn, T. Development and standardization process, and factor analysis of the computerized cognitive function test system for korea adults. J. Korean Neuropsychiatr. Assoc. 2002, 41, 551–562. [Google Scholar]
- Amedi, A.; Malach, R.; Pascual-Leone, A. Negative bold differentiates visual imagery and perception. Neuron 2005, 48, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ramirez, M.; Higgins, B.A.; Rycroft, J.A.; Owen, G.N.; Mahoney, J.; Shpaner, M.; Foxe, J.J. The deployment of intersensory selective attention: A high-density electrical mapping study of the effects of theanine. Clin. Neuropharmacol. 2007, 30, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Molholm, S.; Ritter, W.; Javitt, D.C.; Foxe, J.J. Multisensory visual–auditory object recognition in humans: A high-density electrical mapping study. Cereb. Cortex 2004, 14, 452–465. [Google Scholar] [CrossRef] [PubMed]
- James, T.W.; Humphrey, G.K.; Gati, J.S.; Servos, P.; Menon, R.S.; Goodale, M.A. Haptic study of three-dimensional objects activates extrastriate visual areas. Neuropsychologia 2002, 40, 1706–1714. [Google Scholar] [CrossRef]
- Ernst, M.O.; Bülthoff, H.H. Merging the senses into a robust percept. Trends Cognit. Sci. 2004, 8, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Molholm, S.; Martinez, A.; Shpaner, M.; Foxe, J.J. Object-based attention is multisensory: Co-activation of an object’s representations in ignored sensory modalities. Eur. J. Neurosci. 2007, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Giard, M.-H.; Fort, A.; Mouchetant-Rostaing, Y.; Pernier, J. Neurophysiological mechanisms of auditory selective attention in humans. Front. Biosci. 2000, 5, 84–94. [Google Scholar] [CrossRef]
- Uhlig, C.H.; Gutschalk, A. Transient human auditory cortex activation during volitional attention shifting. PLoS ONE 2017, 12, e0172907. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.E.; Campbell, T.A. Processing complex sounds passing through the rostral brainstem: The new early filter model. Front. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Kanwisher, N.; Wojciulik, E. Visual attention: Insights from brain imaging. Nat. Rev. Neurosci. 2000, 1, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ciaramitaro, V.M.; Buračas, G.T.; Boynton, G.M. Spatial and cross-modal attention alter responses to unattended sensory information in early visual and auditory human cortex. J. Neurophysiol. 2007, 98, 2399–2413. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, B.; Wozny, D.R.; Shams, L. Biases in visual, auditory, and audiovisual perception of space. PLoS Comput. Biol. 2015, 11, e1004649. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.-C.; Prime, S.L. Clarifying the crossmodal stroop effect in an auditory-visual colour naming task with words and non-words stimuli. J. Vis. 2014, 14, 446. [Google Scholar] [CrossRef]
- Amer, T.; Campbell, K.L.; Hasher, L. Cognitive control as a double-edged sword. Trends Cognit. Sci. 2016, 20, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Biss, R.K.; Ngo, K.J.; Hasher, L.; Campbell, K.L.; Rowe, G. Distraction can reduce age-related forgetting. Psychol. Sci. 2013, 24, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.G.; Blumenfeld, R.S.; D’Esposito, M. Disruption of dorsolateral but not ventrolateral prefrontal cortex improves unconscious perceptual memories. J. Neurosci. 2013, 33, 13233–13237. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, J.S.; Maylor, E.A. When distraction holds relevance: A prospective memory benefit for older adults. Int. J. Environ. Res. Public Health 2015, 12, 6523–6541. [Google Scholar] [CrossRef] [PubMed]
- Mozolic, J.L.; Hugenschmidt, C.E.; Peiffer, A.M.; Laurienti, P.J. Multisensory integration and aging. In The Neural Bases of Multisensory Processes; Murray, M.M., Wallace, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Heuninckx, S.; Wenderoth, N.; Swinnen, S.P. Systems neuroplasticity in the aging brain: Recruiting additional neural resources for successful motor performance in elderly persons. J. Neurosci. 2008, 28, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.; Adamo, M.; Haist, F. Changing channels: An fmri study of aging and cross-modal attention shifts. Neuroimage 2006, 31, 1682–1692. [Google Scholar] [CrossRef] [PubMed]
- Venkatraman, V.K.; Aizenstein, H.; Guralnik, J.; Newman, A.B.; Glynn, N.W.; Taylor, C.; Studenski, S.; Launer, L.; Pahor, M.; Williamson, J. Executive control function, brain activation and white matter hyperintensities in older adults. Neuroimage 2010, 49, 3436–3442. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.O.; Adalı, T.; Calhoun, V.D. Estimating the number of independent components for functional magnetic resonance imaging data. Hum. Brain Mapp. 2007, 28, 1251–1266. [Google Scholar] [CrossRef] [PubMed]
- Grady, C.L.; Springer, M.V.; Hongwanishkul, D.; McIntosh, A.R.; Winocur, G. Age-related changes in brain activity across the adult lifespan. J. Cognit. Neurosci. 2006, 18, 227–241. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Presentation Time Mean (M)/Standard Deivation (SD) | |||||||
|---|---|---|---|---|---|---|---|
| 500 | 1000 | 1500 | 2000 | 2500 | 3000 | ||
| Response time (RT) (ms) | Younger | 3719.20 | 3220.79 | 2929.75 | 2825.18 | 2580.51 | 2385.46 |
| (N = 30) | (1436.89) | (1278.81) | (1054.31) | (1093.25) | (916.28) | (865.08) | |
| Older | 4262.778 | 3843.881 | 3536.023 | 3501.994 | 3145.986 | 2952.384 | |
| (N = 22) | (3421.64) | (1702.78) | (1484.28) | (1695.64) | (1465.81) | (1461.03) | |
| Accuracy (%) | Younger | 84.83 | 94.33 | 95.83 | 96.00 | 96.50 | 97.50 |
| (N = 30) | (23.36) | (14.60) | (10.03) | (12.07) | (9.35) | (6.91) | |
| Older | 62.05 | 68.18 | 70.91 | 73.18 | 75.91 | 75.45 | |
| (N = 22) | (33.04) | (30.49) | (33.80) | (33.71) | (29.78) | (30.58) | |
| AV-CITs M(SD) | |||||
|---|---|---|---|---|---|
| Uni-A | Uni-V | Multi-A | Multi-V | ||
| RT (msec) | Younger | 3849.80 | 1752.69 | 3608.48 | 2595.58 |
| (N = 30) | (746.51) | (383.34) | (569.12) | (307.56) | |
| Older | 4989.42 | 1993.77 | 4240.48 | 2938.36 | |
| (N = 22) | (1829.16) | (464.93) | (1027.51) | (587.92) | |
| Accuracy (%) | Younger | 90.83 | 98.61 | 89.03 | 99.58 |
| (N = 30) | (10.51) | (4.68) | (12.21) | (1.27) | |
| Older | 51.14 | 94.51 | 49.81 | 97.35 | |
| (N = 22) | (22.24) | (7.43) | (21.69) | (3.97) | |
| Subgroups on Response Time in the Multi-A Condition | ΔUni-A–Multi-A | |||
|---|---|---|---|---|
| Younger (N = 30) M(SD) | Older (N = 22) M(SD) | |||
| High | 392.66 | (183.91) | 474.99 | (311.43) |
| Middle | 270.66 | (506.92) | 624.14 | (660.79) |
| Low | 38.65 | (953.34) | 1230.85 | (2297.46) |
| Condition | Stroop Task | Uni-A | Uni-V | Multi-A | Multi-V |
|---|---|---|---|---|---|
| 1 | Color naming | 0.30 * | 0.34 * | 0.24 | 0.26 |
| 2 | Word reading | 0.63 ** | 0.37 ** | 0.56 ** | 0.48 ** |
| 3 | Word reading-Congruent | 0.19 | 0.33 * | 0.17 | 0.30 * |
| 4 | Word reading-Incongruent (Color inhibition) | 0.46 ** | 0.36 ** | 0.56 ** | 0.21 |
| 5 | Color reading-Incongruent (Word inhibition) | 0.32 * | 0.47 ** | 0.49 ** | 0.35 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.; Ryu, H.; Kim, J.-K.; Jeong, E. Multisensory Integration Strategy for Modality-Specific Loss of Inhibition Control in Older Adults. Int. J. Environ. Res. Public Health 2018, 15, 718. https://doi.org/10.3390/ijerph15040718
Lee A, Ryu H, Kim J-K, Jeong E. Multisensory Integration Strategy for Modality-Specific Loss of Inhibition Control in Older Adults. International Journal of Environmental Research and Public Health. 2018; 15(4):718. https://doi.org/10.3390/ijerph15040718
Chicago/Turabian StyleLee, Ahreum, Hokyoung Ryu, Jae-Kwan Kim, and Eunju Jeong. 2018. "Multisensory Integration Strategy for Modality-Specific Loss of Inhibition Control in Older Adults" International Journal of Environmental Research and Public Health 15, no. 4: 718. https://doi.org/10.3390/ijerph15040718
APA StyleLee, A., Ryu, H., Kim, J. -K., & Jeong, E. (2018). Multisensory Integration Strategy for Modality-Specific Loss of Inhibition Control in Older Adults. International Journal of Environmental Research and Public Health, 15(4), 718. https://doi.org/10.3390/ijerph15040718