Spatial Pattern and Population Structure of Artemisia ordosica Shrub in a Desert Grassland under Enclosure, Northwest China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Design
2.3. Data Analysis
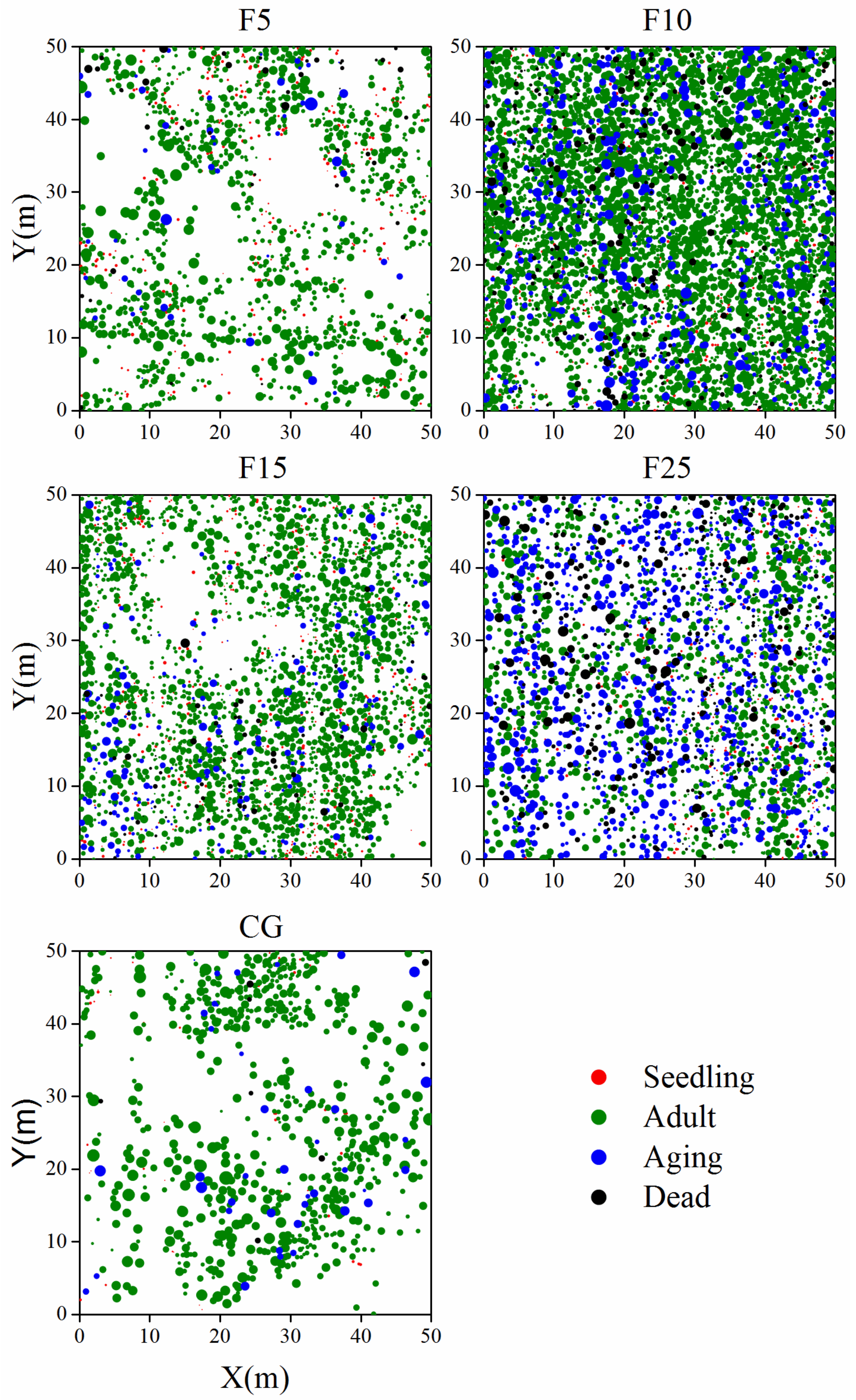
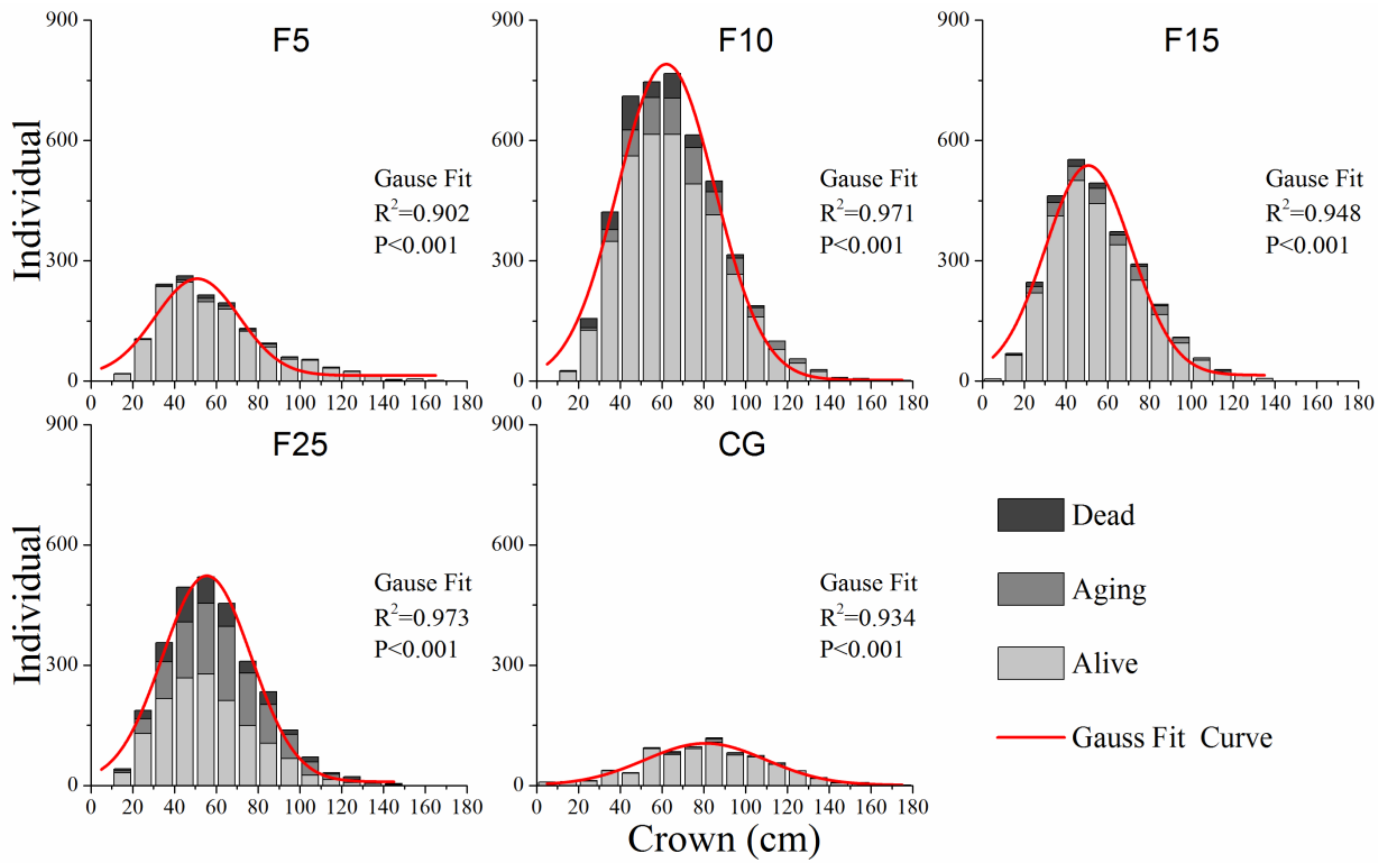
2.3.1. Population Structure
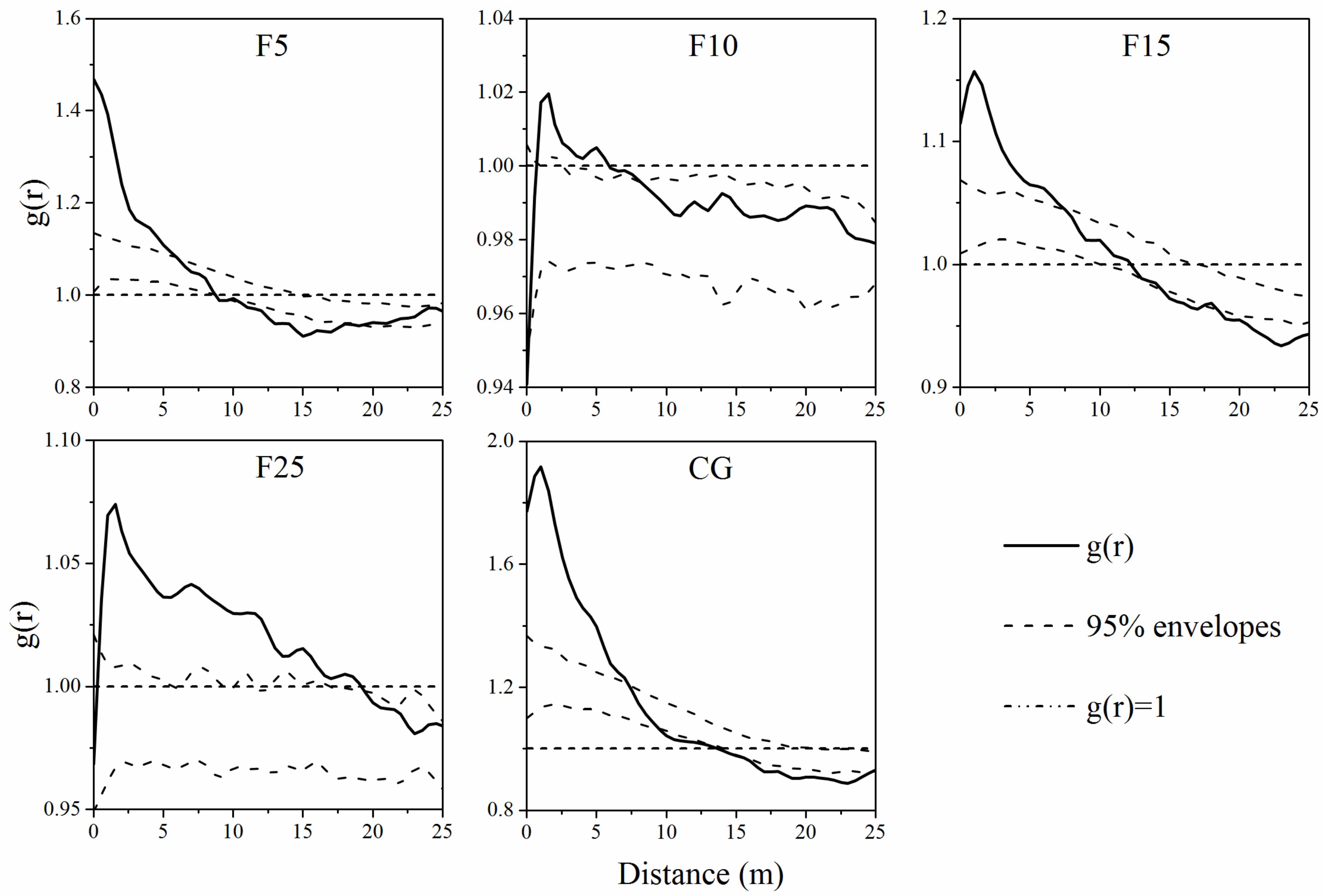
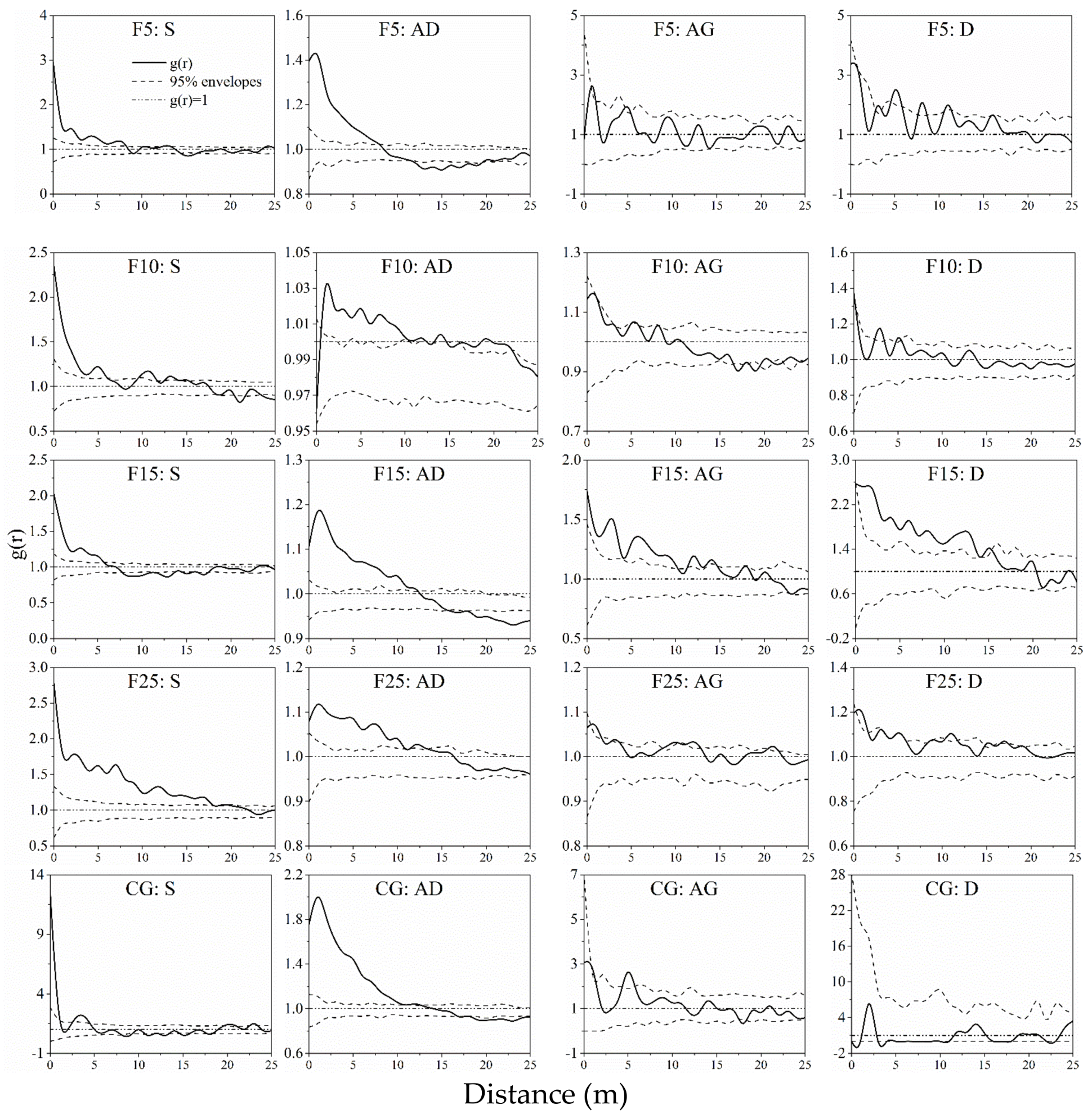
2.3.2. Spatial Pattern Analysis
2.3.3. Species Diversity Index
3. Results
3.1. Population Structure of A. ordosica Populations
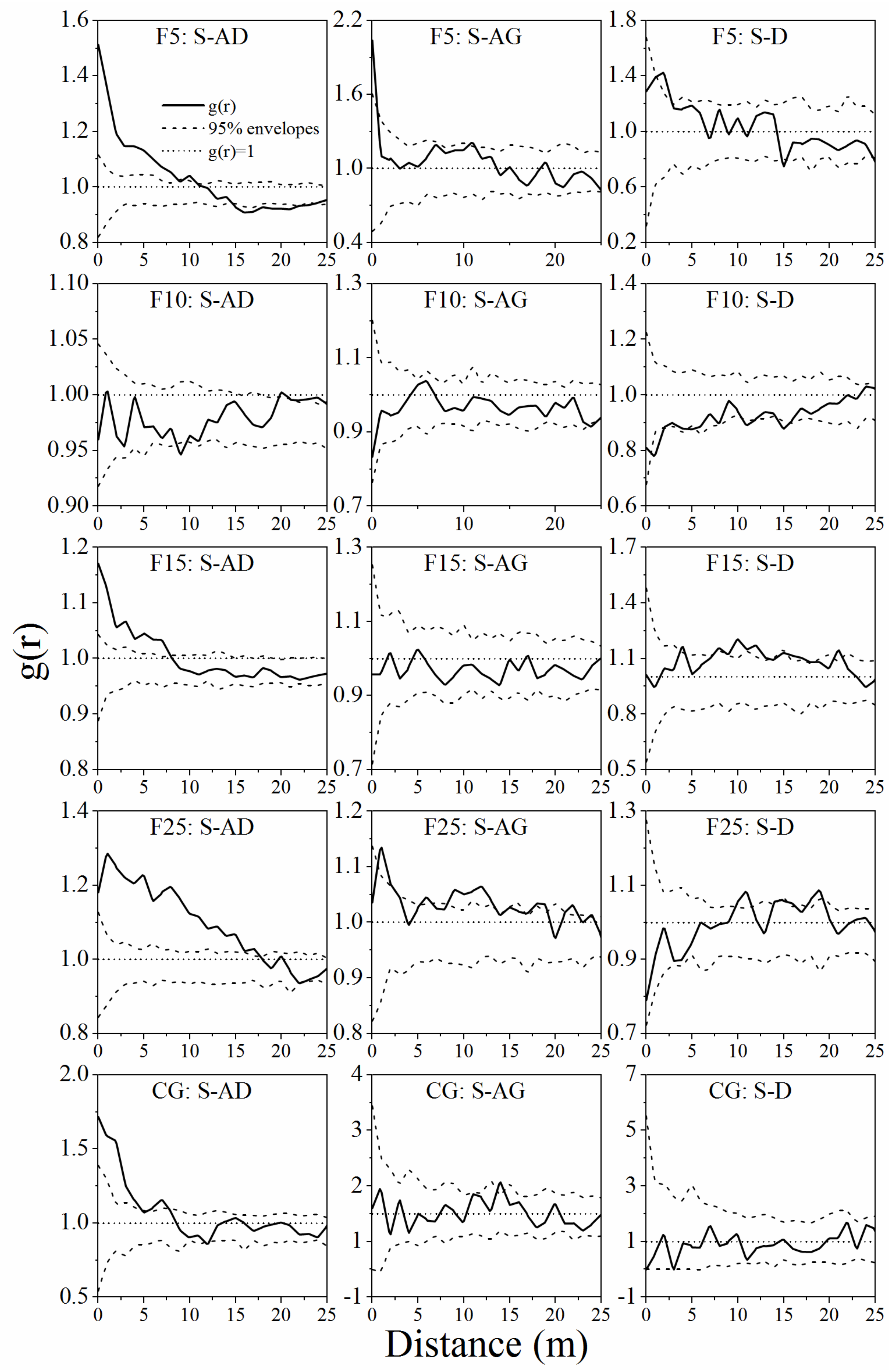
3.2. Spatial Autocorrelation of A. ordosica Populations
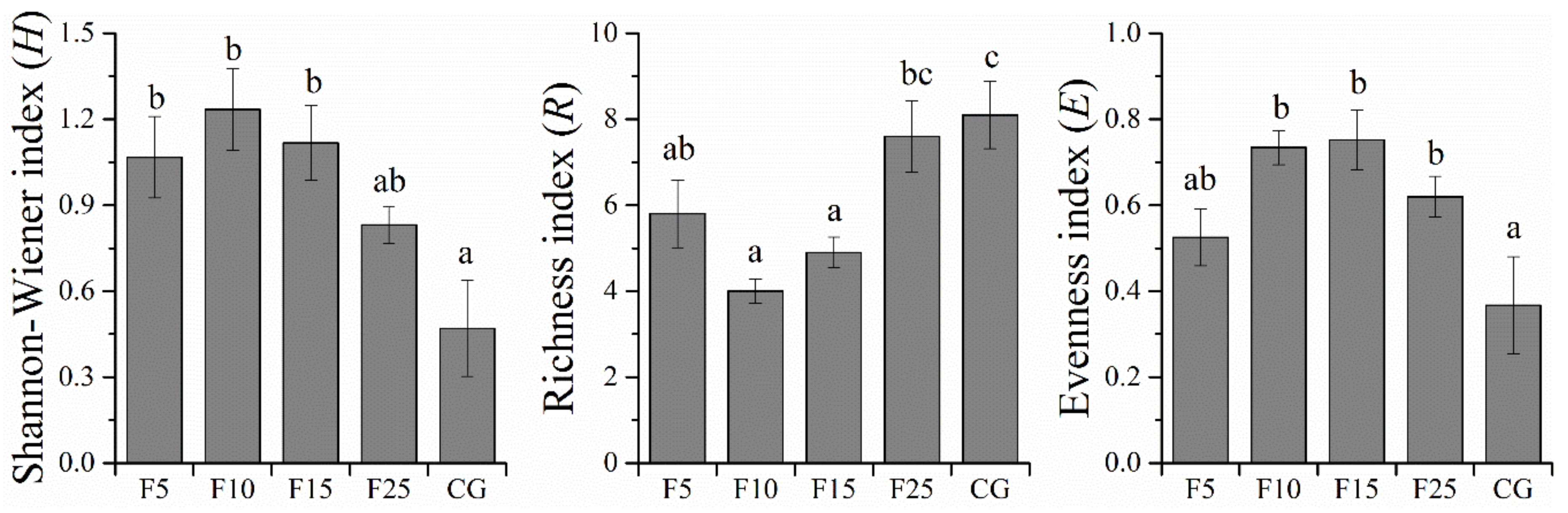
3.3. Species Diversity of Community
4. Discussion
4.1. Population Structure of A. ordosica Population
4.2. Spatial Pattern of A. ordosica Population
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Mara, F.P. The role of grasslands in food security and climate change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Henzell, E.F. Perspectives in Grassland Ecology. Agro-Ecosystems 1981, 7, 169–170. [Google Scholar] [CrossRef]
- Zheng, Y.; Xie, Z.; Jiang, L.; Wu, Y.; Shimizu, H. Model simulation and comparison of the ecological characteristics of three degraded grassland types in China. Belg. J. Bot. 2005, 138, 109–118. [Google Scholar]
- Wang, Z.; Deng, X.; Song, W.; Li, Z.; Chen, J. What is the main cause of grassland degradation? A case study of grassland ecosystem service in the middle-south Inner Mongolia. Catena 2017, 150, 100–107. [Google Scholar] [CrossRef]
- Sivakumar, M.V.K. Interactions between climate and desertification. Agric. For. Meteorol. 2007, 142, 143–155. [Google Scholar] [CrossRef]
- John, R.; Chen, J.; Ou-Yang, Z.T.; Becker, R.; Samanta, A.; Batkhishig, O. Vegetation response to extreme climate events on the Mongolian Plateau from 2000 to 2010. Environ. Res. Lett. 2013, 8, 035033. [Google Scholar] [CrossRef]
- Gang, C.; Zhou, W.; Chen, Y.; Wang, Z.; Sun, Z.; Li, J.; Odeh, I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Qi, J.; Chen, J.; Wan, S.; Ai, L. Understanding the coupled natural and human systems in Dryland East Asia. Environ. Res. Lett. 2012, 7, 015202. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.; Gong, Y.; Wang, Z.; Huang, D. Single irrigation can achieve relatively high production and water use efficiency of Siberian wildrye grass in the semiarid agropastoral ecotone of North China. Agron. J. 2009, 101, 996–1002. [Google Scholar] [CrossRef]
- Andrade, B.O.; Koch, C.; Boldrini, I.I.; Vélezmartin, E.; Hasenack, H.; Hermann, J.M.; Overbeck, G.E. Grassland degradation and restoration: A conceptual framework of stages and thresholds illustrated by southern Brazilian grasslands. Nat. Conserv. 2015, 13, 95–104. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Li, J.; Gang, C.; Zhang, Y.; Zhang, Y.; Qi, J. Comparative assessment of grassland degradation dynamics in response to climate variation and human activities in China, Mongolia, Pakistan and Uzbekistan from 2000 to 2013. J. Arid Environ. 2016, 135, 164–172. [Google Scholar] [CrossRef]
- Jiao, F.; Wen, Z.M.; An, S.S. Changes in soil properties across a chronosequence of vegetation restoration on the Loess Plateau of China. Catena 2011, 86, 110–116. [Google Scholar] [CrossRef]
- Handlová, V.; Münzbergová, Z. Seed banks of managed and degraded grasslands in the Krkonoše Mts., Czech Republic. Folia Geobotanica 2006, 41, 275–288. [Google Scholar] [CrossRef]
- Henning, J.A.G.; Kellner, K. Degradation of a soil (aridosol) and vegetation in the semi-arid grasslands of southern Africa. Bot. Bull. Acad. Sin. 1994, 35, 195–199. [Google Scholar]
- Wu, B.; Yang, H. Spatial patterns and natural recruitment of native shrubs in a semi-arid sandy land. PLoS ONE 2013, 8, e58331. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ci, L.J. Landscape change and desertification development in the Mu Us Sandland, Northern China. J. Arid Environ. 2002, 50, 429–444. [Google Scholar] [CrossRef]
- Wang, X.; Chen, F.H.; Dong, Z.; Xia, D. Evolution of the southern Mu Us Desert in north China over the past 50 years: An analysis using proxies of human activity and climate parameters. Land Degrad. Dev. 2005, 16, 351–366. [Google Scholar] [CrossRef]
- He, Y.H.; Zhao, H.L.; Liu, X.P.; Zhao, X.Y.; LI, Y.Q. Soil physical and chemical characteristics of sandy meadow in natural restoration process. J. Soil Water Conserv. 2008, 22, 159–161. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Xiao, L.; Huang, Y. How Fencing Affects the Soil Quality and Plant Biomass in the Grassland of the Loess Plateau. Int. J. Environ. Res. Public Health 2017, 14, 1117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhao, X.; Tang, Y.; Gu, S.; Zhou, L. Alpine grassland degradation and its control in the source region of the Yangtze and Yellow Rivers, China. Grassl. Sci. 2005, 51, 191–203. [Google Scholar] [CrossRef]
- Batoyun, T.; Shinoda, M.; Cheng, Y.; Purevdorj, Y. Effects of grazing and precipitation variability on vegetation dynamics in a Mongolian dry steppe. J. Plant Ecol. 2016, 9, 508–519. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Deng, L.; Zhang, X.B.; Shangguan, Z.P. Effects of grazing exclusion on plant community and soil physicochemical properties in a desert steppe on the Loess Plateau, China. Ecol. Eng. 2016, 90, 372–381. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.; Wang, J.; Li, X.; Xu, C.; Shi, S. Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Deng, L.; Zhang, Z.; Shangguan, Z. Long-term fencing effects on plant diversity and soil properties in China. Soil Tillage Res. 2014, 137, 7–15. [Google Scholar] [CrossRef]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Shang, Z.H.; Ma, Y.S.; Long, R.J.; Thirgood, S. Effect of fencing, artificial seeding and abandonment on vegetation composition and dynamics of ‘black soil land’ in the headwaters of the Yangtze and the Yellow Rivers of the Qinghai-Tibetan Plateau. Land Degrad. Dev. 2008, 19, 554–563. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Wan, Y.; Jiangcun, W.Z.; Qin, X.B.; Wang, B.S. Significant achievements in protection and restoration of alpine grassland ecosystem in Northern Tibet, China. Restor. Ecol. 2009, 17, 320–323. [Google Scholar] [CrossRef]
- Gómezaparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Armas, C. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Bezemer, T.; Lawson, C.S.; Hedlund, K.; Edwards, A.R.; Brook, A.J.; Igual, J.M.; Van Der Putten, W.H. Plant species and functional group effects on abiotic and microbial soil properties and plant–soil feedback responses in two grasslands. J. Ecol. 2006, 94, 893–904. [Google Scholar] [CrossRef]
- Zuo, X.; Zhao, X.; Zhao, H.; Zhang, T.; Guo, Y.; Li, Y.; Huang, Y. Spatial heterogeneity of soil properties and vegetation–soil relationships following vegetation restoration of mobile dunes in Horqin Sandy Land, Northern China. Plant Soil 2009, 318, 153–167. [Google Scholar] [CrossRef]
- Forup, M.L.; Henson, K.S.E.; Craze, P.G.; Memmott, J. The restoration of ecological interactions: Plant–pollinator networks on ancient and restored heathlands. J. Appl. Ecol. 2008, 45, 742–752. [Google Scholar] [CrossRef]
- Tielbörger, K.; Kadmon, R. Temporal environmental variation tips the balance between facilitation and interference in desert plants. Ecology 2000, 81, 1544–1553. [Google Scholar] [CrossRef]
- An, S.; Zheng, F.; Zhang, F.; Van Pelt, S.; Hamer, U.; Makeschin, F. Soil quality degradation processes along a deforestation chronosequence in the Ziwuling area, China. Catena 2008, 75, 248–256. [Google Scholar] [CrossRef]
- Guo, Y.X.; Kang, B.; Li, G.; Wang, D.X.; Yang, G.H.; Wang, D.W. Species composition and point pattern analysis of standing trees in secondary Betula albosinensis forest in Xiaolongshan of west Qinling Mountains. J. Appl. Ecol. 2011, 22, 2574–2580. [Google Scholar] [CrossRef]
- Druckenbrod, D.L.; Shugart, H.H.; Davies, I. Spatial pattern and process in forest stands within the Virginia piedmont. J. Veg. Sci. 2005, 16, 37–48. [Google Scholar] [CrossRef]
- Mwavu, E.N.; Witkowski, E.T.F. Population structure and regeneration of multiple-use tree species in a semi-deciduous African tropical rainforest: Implications for primate conservation. For. Ecol. Manag. 2009, 258, 840–849. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, L.; Zhang, Z. Numeric Dynamics of Artemisia ordosica Population under Different Habitats in Southeastern Edge of the Tengger Desert. J. Desert Res. 2017, 5, 893–901. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Lee, H.S. Spatial Patterns in the Distribution of Tropical Tree Species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Guo, Y.; Ren, C.; Zhao, F.; Feng, Y.; Han, X.; Yang, G. Population structure and spatial pattern of main tree species in secondary Betula platyphylla forest in Ziwuling Mountains, China. Sci. Rep. 2014, 4, 6873. [Google Scholar] [CrossRef] [PubMed]
- Li, X.R. Study on shrub community diversity of Ordos Plateau, Inner Mongolia, Northern China. J. Arid Environ. 2001, 47, 271–279. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Shi, Z. The formation of the patterns of desert shrub communities on the Western Ordos Plateau, China: The roles of seed dispersal and sand burial. PLoS ONE 2013, 8, e69970. [Google Scholar] [CrossRef] [PubMed]
- King, E.G.; Stanton, M.L. Facilitative effects of Aloe shrubs on grass establishment, growth, and reproduction in degraded Kenyan rangelands: Implications for restoration. Restor. Ecol. 2008, 16, 464–474. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Cui, J.; Zhao, X.; Zhang, T. Nutrient resorption in Caragana microphylla along a chronosequence of plantations: Implications for desertified land restoration in North China. Ecol. Eng. 2013, 53, 299–305. [Google Scholar] [CrossRef]
- Yang, X.; Baskin, C.C.; Baskin, J.M.; Liu, G.; Huang, Z. Seed mucilage improves seedling emergence of a sand desert shrub. PLoS ONE 2012, 7, e34597. [Google Scholar] [CrossRef] [PubMed]
- Padilla, F.M.; Pugnaire, F.I. The role of nurse plants in the restoration of degraded environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- Li, S.L.; Werger, M.J.A.; Zuidema, P.A.; Yu, F.H.; Dong, M. Seedlings of the semi-shrub Artemisia ordosica are resistant to moderate wind denudation and sand burial in Mu Us sandland, China. Trees 2010, 24, 515–521. [Google Scholar] [CrossRef]
- Li, S.L.; Zuidema, P.A.; Yu, F.H.; Werger, M.J.; Dong, M. Effects of denudation and burial on growth and reproduction of Artemisia ordosica in Mu Us sandland. Ecol. Res. 2010, 25, 655–661. [Google Scholar] [CrossRef]
- She, W.; Zhang, Y.; Qin, S.; Wu, B.; Liu, Z.; Liu, J.; Zhang, W. Habitat effect on allometry of a xeric shrub (Artemisia ordosica Krasch) in the Mu Us Desert of northern China. Forests 2015, 6, 4529–4539. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, J.; Li, Z.; Wu, B.; Zhang, Z.; Wang, Y. Comparative study on spatial patterns of the Artemisia ordosica population in the Mu Us sandy land. Acta Ecol. Sin. 2008, 28, 1901–1910. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Fan, D.; Qin, S.; Jia, X.; Wu, B.; Zhu, L. Effects of sand dune stabilization on the spatial pattern of Artemisia ordosica population in Mu Us Desert, Northwest China. PLoS ONE 2015, 10, e0129728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Wu, B.; Lei, Y.K.; Li, X.M.; He, J.; Gao, D.X. Analysis of Artemisia ordosica plant morphology and structure characteristics in Mu Us Sandland. J. Southwest For. Univ. 2011, 31, 6–9. [Google Scholar] [CrossRef]
- Xue, P.; He, X. Physiological water strategy of Artemisia ordosica around soil threshold of drought. Front. Biol. China 2008, 3, 32–39. [Google Scholar] [CrossRef]
- Zhang, J.H.; Han, H.Y.; Lei, Y.K.; Yang, W.B.; Li, Y.H.; Yang, D.F.; Zhao, X.B. Correlations between distribution characteristics of Artemisia ordosica root system and soil moisture under different fixation stage of sand dunes. J. Southwest For. Univ. 2012, 32, 1–5. [Google Scholar] [CrossRef]
- Zhao, X.L.; He, X.D.; Xue, P.P.; Zhang, N.; Wu, W.; Li, R.; Zhao, H. Effects of soil stoichiometry of the CaCO3/available phosphorus ratio on plant density in Artemisia ordosica communities. Chin. Sci. Bull. 2012, 57, 492–499. [Google Scholar] [CrossRef]
- She, W.; Bai, Y.; Zhang, Y.; Qin, S.; Liu, Z.; Wu, B. Plasticity in Meristem Allocation as an Adaptive Strategy of a Desert Shrub under Contrasting Environments. Front. Plant Sci. 2017, 8, 1933. [Google Scholar] [CrossRef]
- Kobayashi, T.; Liao, R.T.; Li, S.Q. Ecophysiological behavior of Artemisia ordosica on the process of sand dune fixation. Ecol. Res. 1995, 10, 339–349. [Google Scholar] [CrossRef]
- Wei, B.; Ding, G.D.; Wu, B.; Zhang, Y.Q. Study on Windbreak Effect of Artemisia ordosica Community over Different Vegetation Coverage. Arid Zone Res. 2012, 29, 213–217. [Google Scholar] [CrossRef]
- Li, S.L.; Yu, F.H.; Werger, M.J.A.; Dong, M.; Zuidema, P.A. Habitat-specific demography across dune fixation stages in a semi-arid sandland: Understanding the expansion, stabilization and decline of a dominant shrub. J. Ecol. 2011, 99, 610–620. [Google Scholar] [CrossRef]
- Ma, J.; Li, J. A comparative study on microstructure features of seed from Artemisia plants in desert of China. J. Desert Res. 2002, 22, 586–590. [Google Scholar]
- Martinezmeza, E.; Whitford, W.G. Stemflow, throughfall and channelization of stemflow by roots in three Chihuahuan desert shrubs. J. Arid Environ. 1996, 32, 271–287. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Haase, P.; Puigdefábregas, J.; Cueto, M.; Clark, S.C.; Incoll, L.D. Facilitation and succession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 1996, 76, 455–464. [Google Scholar] [CrossRef]
- Zhang, Z.S.; Zhao, Y.; Li, X.R.; Huang, L.; Tan, H.J. Gross rainfall amount and maximum rainfall intensity in 60-minute influence on interception loss of shrubs: A 10-year observation in the Tengger Desert. Sci. Rep. 2016, 6, 26030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Hu, R.; Pan, Y.X. Stemflow volume per unit rainfall as a good variable to determine the relationship between stemflow amount and morphological metrics of shrubs. J. Arid Environ. 2017, 141, 1–6. [Google Scholar] [CrossRef]
- Wang, J.H.; Jin, H.J.; Ma, Q.L.; Zhang, D.K.; Liu, Y.J.; Xu, B.Y. Structure and Distribution Pattern of Artermisia Ordosica Population in Arid Region. J. Desert Res. 2010, 30, 534–538. [Google Scholar]
- Zhao, Y.; Zhang, Z.; Chen, Y.; Xu, B.X.; Chen, D. Effect of Degradation Stage of Artemisia ordosica on Morphology of Nebkhas. J. Desert Res. 2015, 35, 1136–1140. [Google Scholar] [CrossRef]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Spatial Analysis and Modelling of Spatial Point Patterns. Technometrics 2008, 47, 516–517. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, X.; Li, L.; Shi, Z.; Yang, X. Spatial Association of Shrubs and Their Interrelation to Burrowing Site Preference of Subterranean Rodents on Dune Slope in the Otindag Sandy Land, China. Sustainability 2017, 9, 1729. [Google Scholar] [CrossRef]
- Wiegand, T.; A Moloney, K. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, B.; Zhang, K.; Zhou, J.; Ahmad, B. The Spatial Pattern and Interactions of Woody Plants on the Temperate Savanna of Inner Mongolia, China: The Effects of Alternating Seasonal Grazing-Mowing Regimes. PLoS ONE 2015, 10, e0133277. [Google Scholar] [CrossRef] [PubMed]
- Getzin, S.; Dean, C.; He, F.; A Trofymow, J.; Wiegand, K.; Wiegand, T. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography 2006, 29, 671–682. [Google Scholar] [CrossRef]
- Abeli, T.; Cauzzi, P.; Rossi, G.; Adorni, M.; Vagge, I.; Parolo, G.; Orsenigo, S. Restoring population structure and dynamics in translocated species: Learning from wild populations. Plant Ecol. 2016, 217, 183–192. [Google Scholar] [CrossRef]
- Mori, A.; Takeda, H. Effects of undisturbed canopy structure on population structure and species coexistence in an old-growth subalpine forest in central Japan. For. Ecol. Manag. 2004, 200, 89–100. [Google Scholar] [CrossRef]
- Vidal, S. Plant biodiversity and vegetation structure in traditional cocoa forest gardens in southern Cameroon under different management. Biodivers. Conserv. 2008, 17, 1821–1835. [Google Scholar] [CrossRef]
- Wadt, L.H.O.; Kainer, K.A.; Gomes-Silva, D.A.P. Population structure and nut yield of a Bertholletia excelsa stand in Southwestern Amazonia. For. Ecol. Manag. 2005, 211, 371–384. [Google Scholar] [CrossRef]
- Hagiwara, Y.; Kachi, N.; Suzuki, J. Combined effects between temporal heterogeneity of water supply, nutrient level, and population density on biomass of four broadly distributed herbaceous species. J. Plant Res. 2012, 125, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Kraaij, T.; Ward, D. Effects of rain, nitrogen, fire and grazing on tree recruitment and early survival in bush-encroached savanna, South Africa. Plant Ecol. 2006, 186, 235–246. [Google Scholar] [CrossRef]
- Gómez, D.A.; Aranibar, J.N.; Tabeni, S.; Villagra, P.E.; Garibotti, I.A.; Atencio, A. Biological soil crust recovery after long-term grazing exclusion in the Monte Desert (Argentina). Changes in coverage, spatial distribution, and soil nitrogen. Acta Oecol. 2012, 38, 33–40. [Google Scholar] [CrossRef]
- Read, C.F.; Duncan, D.H.; Vesk, P.A.; Elith, J. Surprisingly fast recovery of biological soil crusts following livestock removal in southern Australia. J. Veg. Sci. 2011, 22, 905–916. [Google Scholar] [CrossRef]
- Oba, G.; Vetaas, O.R.; Stenseth, N.C. Relationships between biomass and plant species richness in arid-zone grazing lands. J. Appl. Ecol. 2001, 38, 836–845. [Google Scholar] [CrossRef]
- King, D.A.; Maindonald, J.H. Tree architecture in relation to leaf dimensions and tree stature in temperate and tropical rain forests. J. Ecol. 1999, 87, 1012–1024. [Google Scholar] [CrossRef]
- Kitajima, K.; Mulkey, S.S.; Wright, S.J. Variation in crown light utilization characteristics among tropical canopy trees. Ann. Bot. 2004, 95, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Grellier, S.; Barot, S.; Janeau, J.L.; Ward, D. Grass competition is more important than seed ingestion by livestock for Acacia recruitment in South Africa. Plant Ecol. 2012, 213, 899–908. [Google Scholar] [CrossRef]
- Kimball, S.; Schiffman, P.M. Differing effects of cattle grazing on native and alien plants. Conserv. Biol. 2003, 17, 1681–1693. [Google Scholar] [CrossRef]
- Lin, Y.C.; Chang, L.W.; Yang, K.C.; Wang, H.H.; Sun, I.F. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia 2011, 165, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Coop, J.D.; Massatti, R.T.; Schoettle, A.W. Subalpine vegetation pattern three decades after stand-replacing fire: Effects of landscape context and topography on plant community composition, tree regeneration, and diversity. J. Veg. Sci. 2010, 21, 472. [Google Scholar] [CrossRef]
- Lin, Y.; Hong, M.; Han, G.; Zhao, M.; Bai, Y.; Chang, S.X. Grazing intensity affected spatial patterns of vegetation and soil fertility in a desert steppe. Agric. Ecosyst. Environ. 2010, 138, 282–292. [Google Scholar] [CrossRef]
- Alados, C.L.; ElAich, A.; Papanastasis, V.P.; Ozbek, H.; Navarro, T.; Freitas, H.; Cabezudo, B. Change in plant spatial patterns and diversity along the successional gradient of Mediterranean grazing ecosystems. Ecol. Model. 2004, 180, 523–535. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species associations in a heterogeneous Sri Lankan dipterocarp forest. Am. Nat. 2007, 170, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Gutterman, Y. Regeneration of Plants in Arid Ecosystems Resulting from Patch Disturbance; Springer Science & Business Media: Berlin, Germany, 2001. [Google Scholar]
- Adler, P.; Raff, D.; Lauenroth, W. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia 2001, 128, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Kawamura, K. Grassland degradation in China: Methods of monitoring, management and restoration. Grassl. Sci. 2007, 53, 1–17. [Google Scholar] [CrossRef]
- Nedessa, B.; Ali, J.; Nyborg, I. Exploring ecological and socio-economic issues for the improvement of area enclosure management: A case study from Ethiopia. Drylands Coord. Group Rep. 2005, 12, 95–107. [Google Scholar]
- Zuo, X.; Zhao, H.; Zhao, X.; Zhang, T.; Guo, Y.; Wang, S.; Drake, S. Spatial pattern and heterogeneity of soil properties in sand dunes under grazing and restoration in Horqin Sandy Land, Northern China. Soil Tillage Res. 2008, 99, 202–212. [Google Scholar] [CrossRef]
- He, F.; Duncan, R.P. Density-dependent effects on tree survival in an old-growth Douglas fir forest. J. Ecol. 2000, 88, 676–688. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Coomes, D.A.; Rees, M.; Turnbull, L. Identifying aggregation and association in fully mapped spatial data. Ecology 1999, 80, 554–565. [Google Scholar] [CrossRef]
- Defalco, L.A.; Esque, T.C.; Kane, J.M.; Nicklas, M.B. Seed banks in a degraded desert shrubland: Influence of soil surface condition and harvester ant activity on seed abundance. J. Arid Environ. 2009, 73, 885–893. [Google Scholar] [CrossRef]
- Giladi, I.; Segoli, M.; Ungar, E.D. Shrubs and herbaceous seed flow in a semi-arid landscape: Dual functioning of shrubs as trap and barrier. J. Ecol. 2013, 101, 97–106. [Google Scholar] [CrossRef]
- Aguiar, M.R.; Soriano, A.; Sala, O.E. Competition and facilitation in the recruitment of seedlings in Patagonian steppe. Funct. Ecol. 1992, 66–70. [Google Scholar] [CrossRef]
- Hao, H.M.; Lu, R.; Liu, Y.; Fang, N.F.; Wu, G.L.; Shi, Z.H. Effects of shrub patch size succession on plant diversity and soil water content in the water-wind erosion crisscross region on the Loess Plateau. Catena 2016, 144, 177–183. [Google Scholar] [CrossRef]
- Brantley, S.T.; Young, D.R. Contribution of sunflecks is minimal in expanding shrub thickets compared to temperate forest. Ecology 2009, 90, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Aarssen, L.W.; Schamp, B.S.; Wight, S. Big plants—Do they affect neighbourhood species richness and composition in herbaceous vegetation? Acta Oecol. 2014, 55, 36–42. [Google Scholar] [CrossRef]
- Pérez, F.L. Geoecological alteration of surface soils by the Hawaiiansilversword (Argyroxiphium sandwicense DC.) in Haleakala’s crater, Maui. Plant Ecol. 2001, 157, 215–233. [Google Scholar] [CrossRef]
- Shumway, S.W. Facilitative effects of a sand dune shrub on species growing beneath the shrub canopy. Oecologia 2000, 124, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; He, M.; Li, X.; Chen, Y.; Liu, L. Effects of Salsola passerina shrub patches on the microscale heterogeneity of soil in a montane grassland, China. J. Arid Environ. 2008, 72, 150–161. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Number | Coverage (%) | Slope | Altitude | Soil Type | Geographic Coordinates | |
|---|---|---|---|---|---|---|
| Latitude (N) | Longitude (E) | |||||
| F5 | 37 | 1–3° | 1393 m | Sierozem | 37°50′54.4” | 107°23′12.7” |
| F10 | 75 | 1–2° | 1393 m | Sierozem | 37°50′49.9” | 107°23′49.0” |
| F15 | 40 | 1–3° | 1394 m | Sierozem | 37°50′46.3″ | 107°23′48.3″ |
| F26 | 48 | 1–3° | 1396 m | Sierozem | 37°50′46.1″ | 107°24′07.8″ |
| CG | 25 | 2–4° | 1395 m | Sierozem | 37°50′40.5″ | 107°24′31.3″ |
| Plot Number | Population Individuals of Different Groups | Population Coverage (%) | Crown (m) | Height (m) | ||||
|---|---|---|---|---|---|---|---|---|
| Seedling (%) | Adult (%) | Aging (%) | Dead (%) | Total | ||||
| F5 | 279 (19.1) | 1089 (74.7) | 46 (3.2) | 44 (3.0) | 1458 | 19.7 | 0.601 ± 0.262 b | 0.289 ± 0.102 a |
| F10 | 257 (5.5) | 3523 (75.9) | 536 (11.5) | 327 (7.0) | 4643 | 71.7 | 0.661 ± 0.236 c | 0.355 ± 0.115 c |
| F15 | 387 (13.3) | 2201 (75.9) | 229 (7.9) | 83 (2.9) | 2900 | 32.6 | 0.556 ± 0.221 a | 0.341 ± 0.111 b |
| F25 | 216 (7.5) | 1294 (45.0) | 980 (34.1) | 385 (13.4) | 2875 | 36.7 | 0.596 ± 0.226 b | 0.352 ± 0.126 c |
| CG | 53 (6.8) | 678 (87.0) | 40 (5.1) | 8 (1.0) | 779 | 18.3 | 0.814 ± 0.294 d | 0.387 ± 0.138 d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, K. Spatial Pattern and Population Structure of Artemisia ordosica Shrub in a Desert Grassland under Enclosure, Northwest China. Int. J. Environ. Res. Public Health 2018, 15, 946. https://doi.org/10.3390/ijerph15050946
Liu J, Zhang K. Spatial Pattern and Population Structure of Artemisia ordosica Shrub in a Desert Grassland under Enclosure, Northwest China. International Journal of Environmental Research and Public Health. 2018; 15(5):946. https://doi.org/10.3390/ijerph15050946
Chicago/Turabian StyleLiu, Jiankang, and Kebin Zhang. 2018. "Spatial Pattern and Population Structure of Artemisia ordosica Shrub in a Desert Grassland under Enclosure, Northwest China" International Journal of Environmental Research and Public Health 15, no. 5: 946. https://doi.org/10.3390/ijerph15050946
APA StyleLiu, J., & Zhang, K. (2018). Spatial Pattern and Population Structure of Artemisia ordosica Shrub in a Desert Grassland under Enclosure, Northwest China. International Journal of Environmental Research and Public Health, 15(5), 946. https://doi.org/10.3390/ijerph15050946