Modeling the Heterogeneity of Dengue Transmission in a City

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
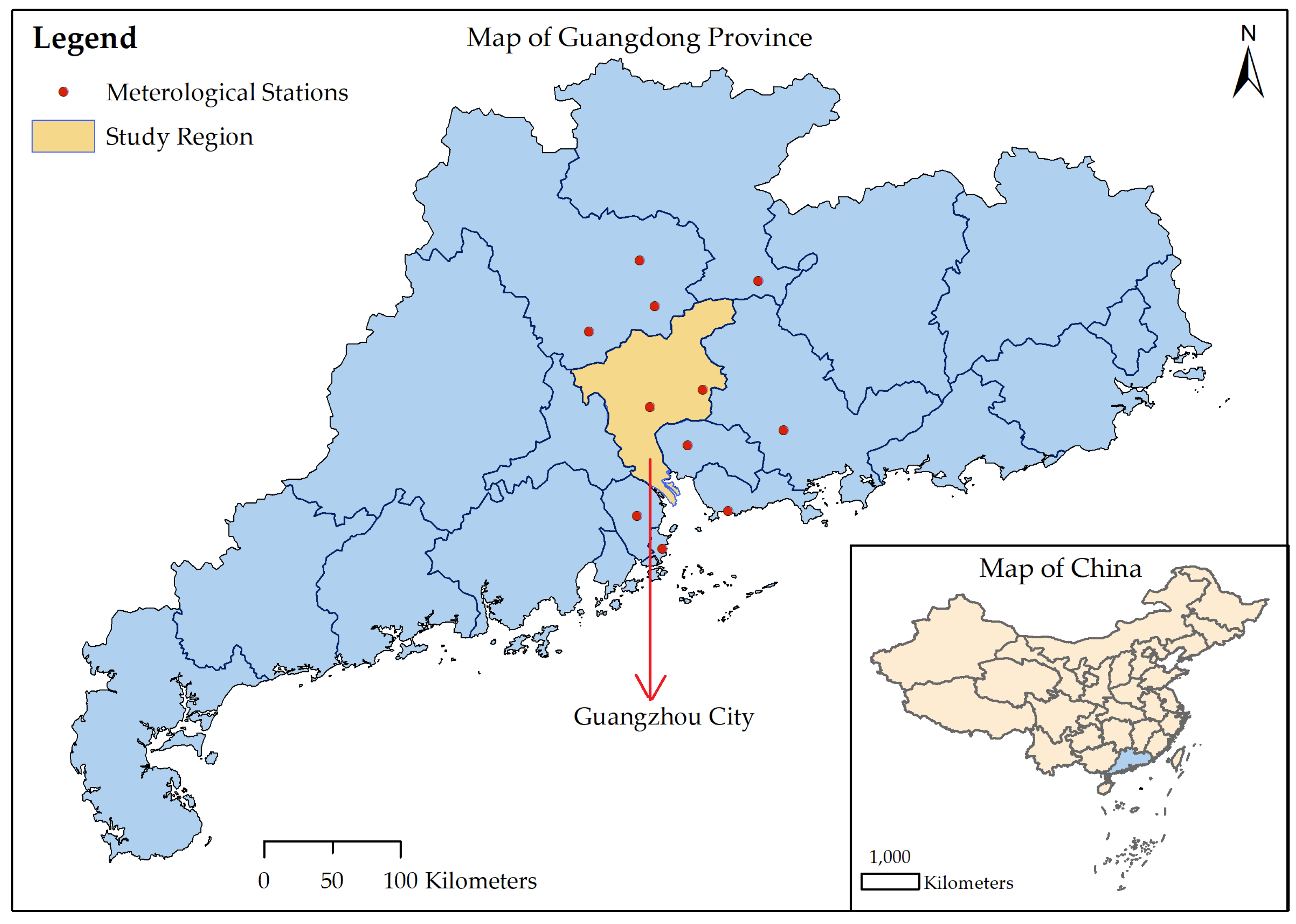
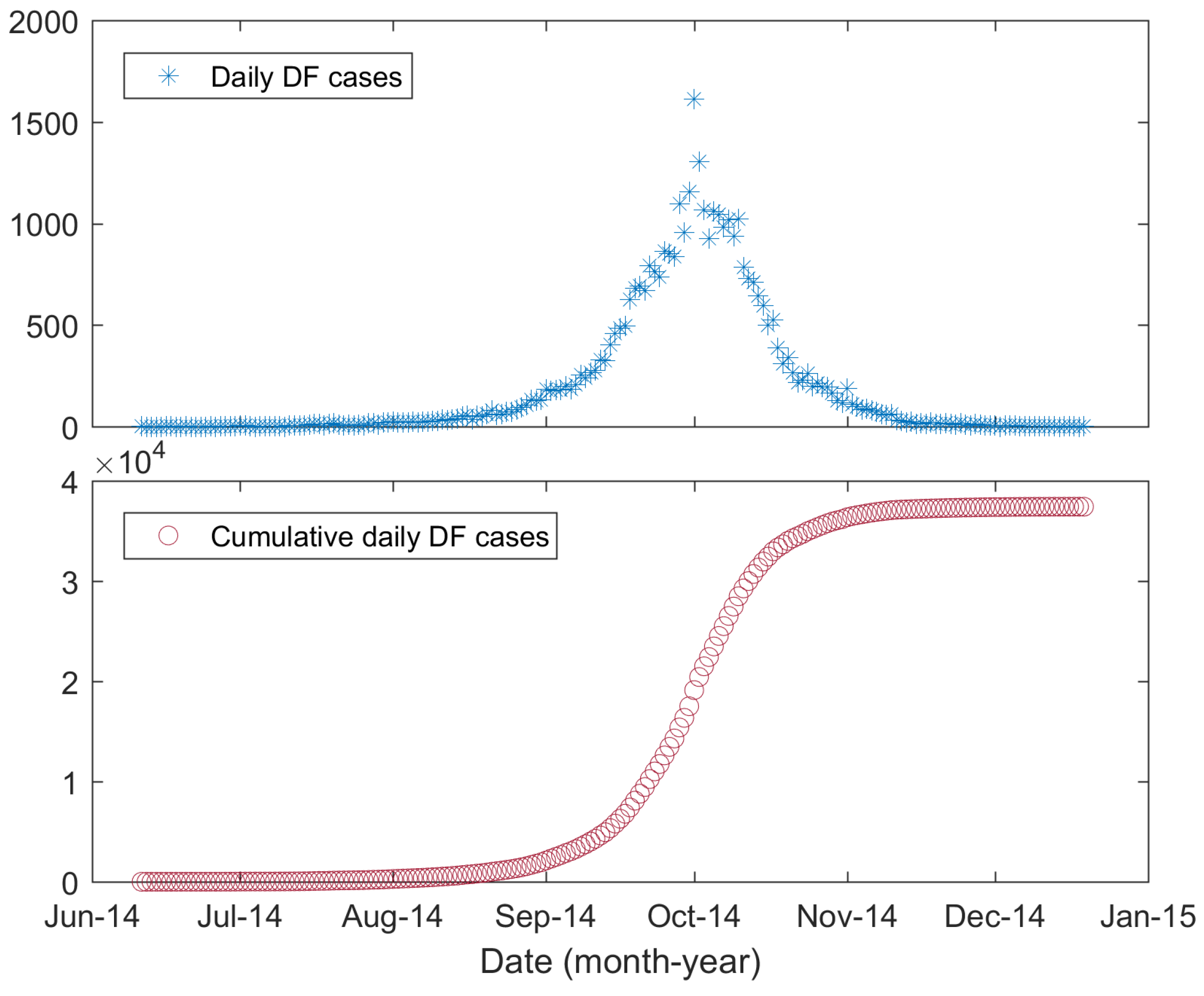
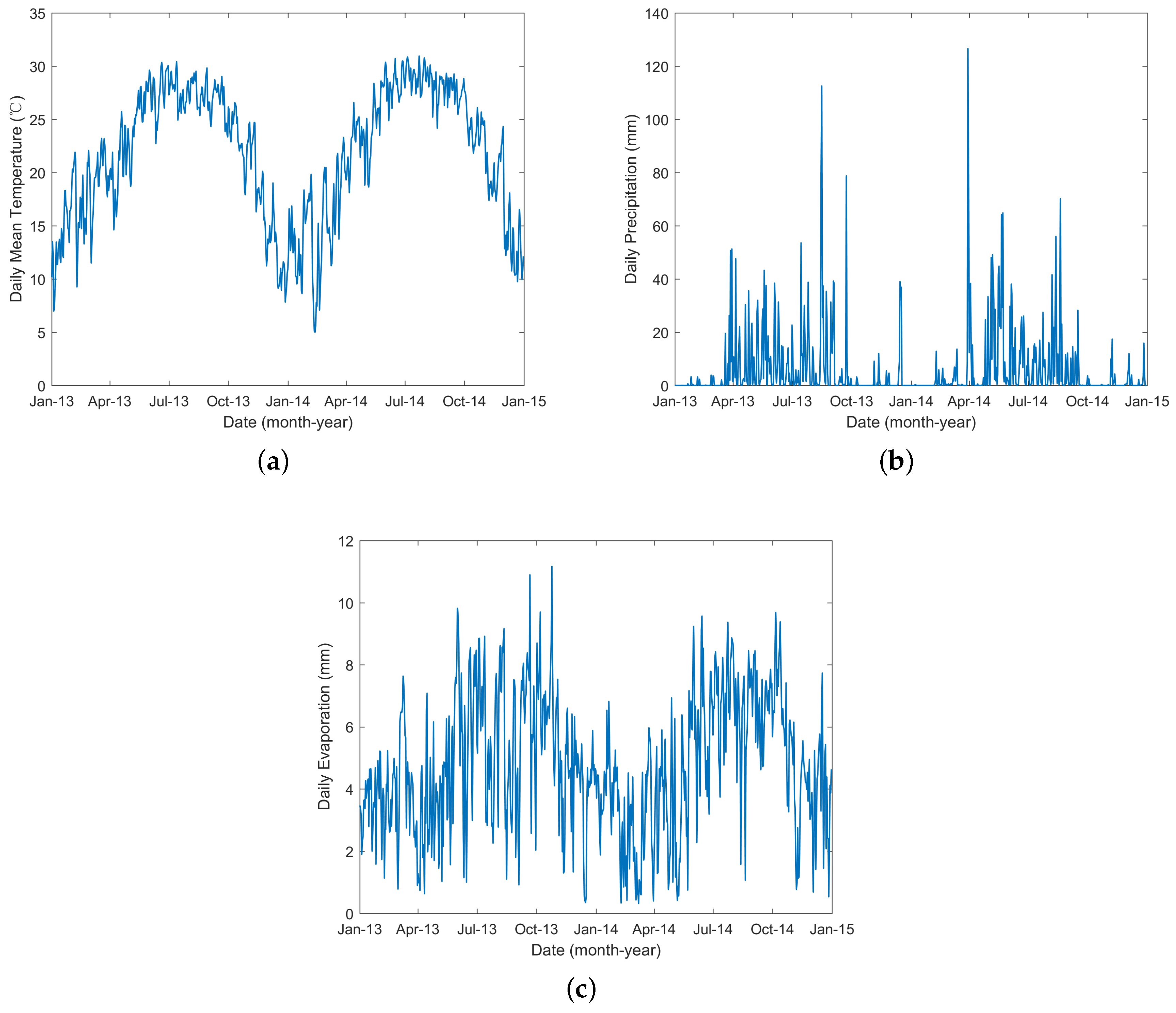
2.1. Data
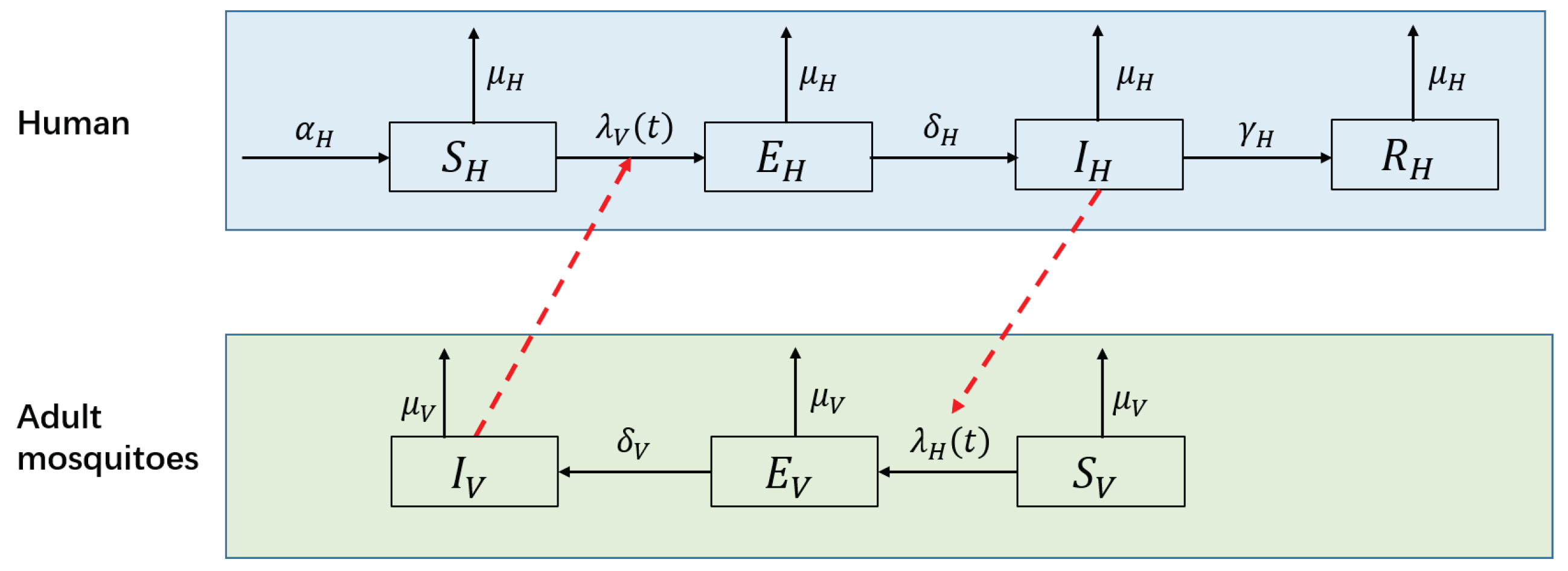
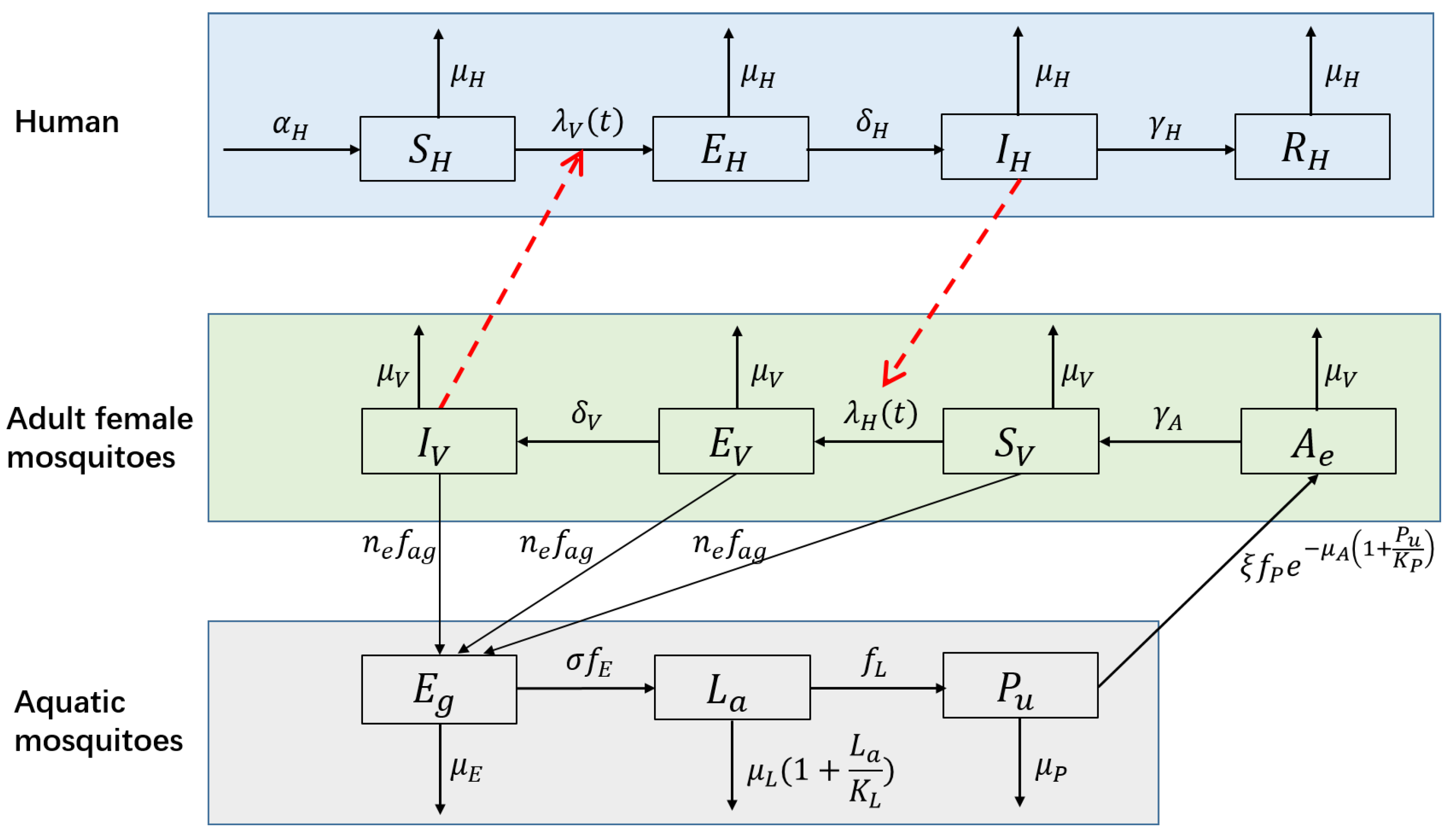
2.2. The NBD SEIR-SEI Model
2.3. Extended Model
2.4. Model Calculation
3. Results
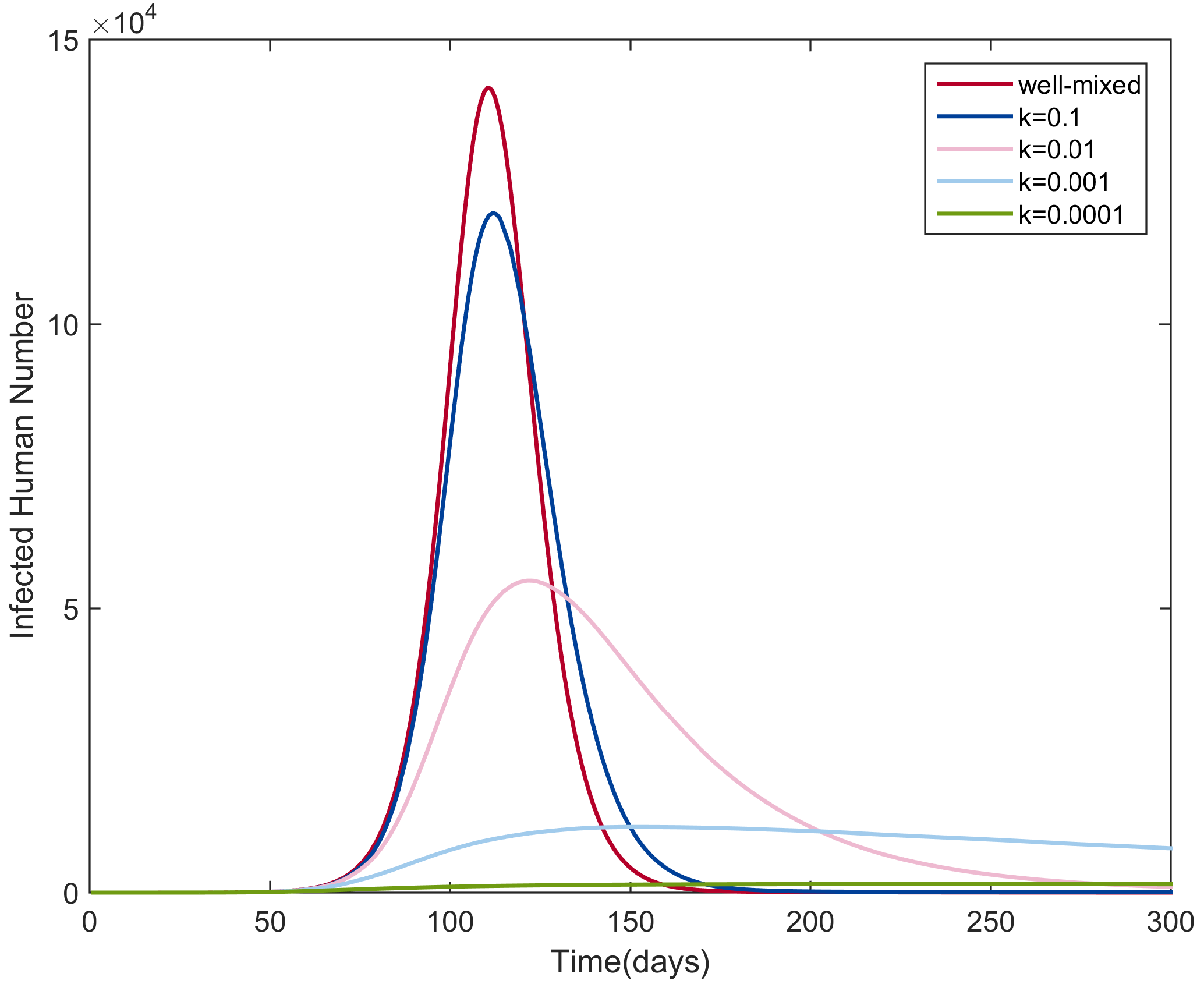
3.1. Transmission Dynamics with Different Heterogeneity Levels
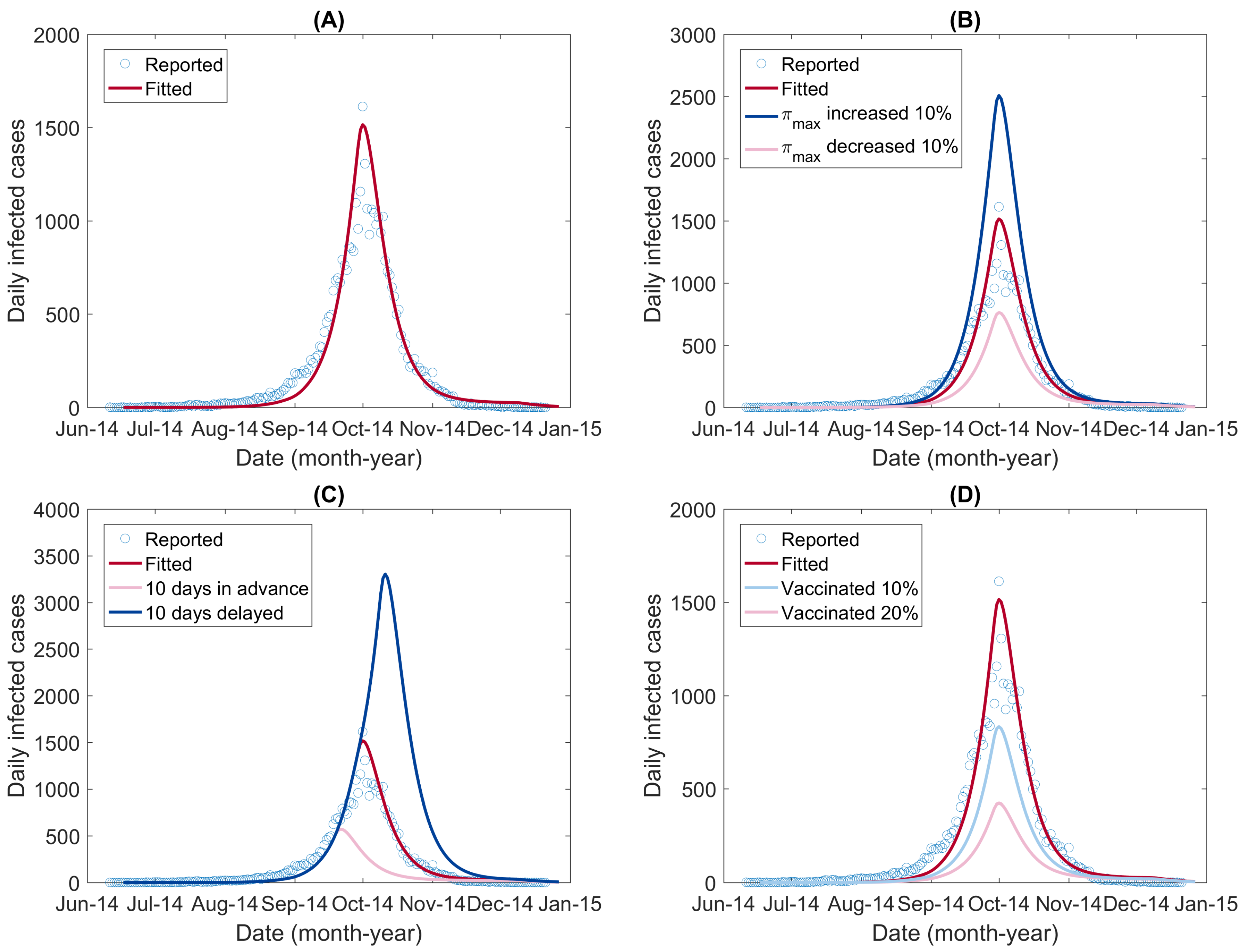
3.2. Model Calibration Results
3.3. Simulation of Dengue Transmission in Different Scenarios
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| DF | dengue fever |
| DENV | dengue virus |
| NBD | negative binomial distribution |
| I:S | inapparent-to-symptomatic ratio |
Appendix A. The Basic Reproductive Number
Appendix B. The Parameters Dependent on Temperature and Precipitation
Appendix B.1. Eggs per Gonotrophic Cycle
Appendix B.2. Reciprocal of the Duration for Gonotrophic Cycle
Appendix B.3. Water Level
Appendix B.4. Spillover Effect
Appendix B.5. Egg Development Rate
Appendix B.6. Larva Development Rate
Appendix B.7. Mortality Rate for Larva
Appendix B.8. Development Rate of Pupae to Emerging Adults
Appendix B.9. Mortality Rate for Pupae
Appendix B.10. Death Rate for Adults
Appendix B.11. Extrinsic Incubation Period
Appendix B.12. Biting Rate
References
- World Health Organization. Global Strategy for Dengue Prevention and Control: 2012–2020; WHO Press: Geneva, Switzerland, 2012. [Google Scholar]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Dengue and Severe Dengue—Fact sheet (Updated April 2017). 2017. Available online: http://www.who.int/mediacentre/factsheets/fs117/en/ (accessed on 20 March 2018).
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, D.J. Dengue, Urbanization and Globalization: The Unholy Trinity of the 21(st) Century. Trop. Med. Health 2011, 39, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.E.; Quam, M.B.; Wilder-Smith, A. Epidemiology of dengue: Past, present and future prospects. Clin. Epidemiol. 2013, 5, 299–309. [Google Scholar] [PubMed]
- Johansson, M.A.; Hombach, J.; Cummings, D.A.T. Models of the impact of dengue vaccines: A review of current research and potential approaches. Vaccine 2011, 29, 5860–5868. [Google Scholar] [CrossRef] [PubMed]
- Andraud, M.; Hens, N.; Marais, C.; Beutels, P. Dynamic Epidemiological Models for Dengue Transmission: A Systematic Review of Structural Approaches. PLoS ONE 2012, 7, e49085. [Google Scholar] [CrossRef] [PubMed]
- Reiner, R.C., Jr.; Perkins, T.A.; Barker, C.M.; Niu, T.; Fernando Chaves, L.; Ellis, A.M.; George, D.B.; Le Menach, A.; Pulliam, J.R.C.; Bisanzio, D.; et al. A systematic review of mathematical models of mosquito-borne pathogen transmission: 1970-2010. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, M.A.; Kazura, J.W.; Hollingsworth, T.D.; Reimer, L.J. Understanding heterogeneities in mosquito-bite exposure and infection distributions for the elimination of lymphatic filariasis. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed]
- Perkins, T.A.; Scott, T.W.; Le Menach, A.; Smith, D.L. Heterogeneity, mixing, and the spatial scales of mosquito-borne pathogen transmission. PLoS Comput. Biol. 2013, 9, e1003327. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, S.T.; Forshey, B.M.; Morrison, A.C.; Paz-Soldan, V.A.; Vazquez-Prokopec, G.M.; Astete, H.; Reiner, R.C.J.; Vilcarromero, S.; Elder, J.P.; Halsey, E.S.; et al. House-to-house human movement drives dengue virus transmission. Proc. Natl. Acad. Sci. USA 2013, 110, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Fleisher, A.; Ruiz-Moreno, D.; Vermeylen, F.; Wa, C.V.; Poulson, R.L.; Edman, J.D.; Clark, J.M.; Jones, J.W.; Kitthawee, S.; et al. Heterogeneous Feeding Patterns of the Dengue Vector, Aedes aegypti, on Individual Human Hosts in Rural Thailand. PLoS Negl. Trop. Dis. 2014, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Dye, C.; Etard, J.F.; Smith, T.; Charlwood, J.D.; Garnett, G.P.; Hagan, P.; Hii, J.L.K.; Ndhlovu, P.D.; Quinnell, R.J.; et al. Heterogeneities in the transmission of infectious agents: Implications for the design of control programs. Proc. Natl. Acad. Sci. USA 1997, 94, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Cosner, C.; Beier, J.C.; Cantrell, R.S.; Impoinvil, D.; Kapitanski, L.; Potts, M.D.; Troyo, A.; Ruan, S. The effects of human movement on the persistence of vector-borne diseases. J. Theor. Biol. 2009, 258, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Nevai, A.L.; Soewono, E. A model for the spatial transmission of dengue with daily movement between villages and a city. Math. Med. Biol. 2014, 31, 150–178. [Google Scholar] [CrossRef] [PubMed]
- Minnick, S.L.; Morrison, A.C.; Scott, T.W. An individual based model for heterogeneous dengue transmission incorporating both age-dependent biting and spatial heterogeneity. Am. J. Trop. Med. Hyg. 2006, 75, 109. [Google Scholar]
- Chao, D.L.; Longini, I.M., Jr.; Halloran, M.E. The Effects of Vector Movement and Distribution in a Mathematical Model of Dengue Transmission. PLoS ONE 2013, 8, e76044. [Google Scholar] [CrossRef] [PubMed]
- Karl, S.; Halder, N.; Kelso, J.K.; Ritchie, S.A.; Milne, G.J. A spatial simulation model for dengue virus infection in urban areas. BMC Infect. Dis. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- May, R.M. Host-Parasitoid Systems in Patchy Environments—Phenomenological Model. J. Anim. Ecol. 1978, 47, 833–844. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Hassell, M.P. Discrete and Continuous Insect Populations in Tropical Environments. J. Anim. Ecol. 1989, 58, 153–174. [Google Scholar] [CrossRef]
- Briggs, C.J.; Godfray, H.C.J. The dynamics of insect-pathogen interactions in stage-structured populations. Am. Nat. 1995, 145, 855–887. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; Schreiber, S.J.; Kopp, P.E.; Getz, W.M. Superspreading and the effect of individual variation on disease emergence. Nature 2005, 438, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Kucharski, A.J.; Althaus, C.L. The role of superspreading in Middle East respiratory syndrome coronavirus (MERS-CoV) transmission. Euro Surveill. 2015, 20, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Padmanabha, H.; Durham, D.; Correa, F.; Diuk-Wasser, M.; Galvani, A. The interactive roles of Aedes aegypti super-production and human density in dengue transmission. PLoS Negl. Trop. Dis. 2012, 6, e1799. [Google Scholar] [CrossRef] [PubMed]
- Barlow, N.D. Non-linear transmission and simple models for bovine tuberculosis. J. Anim. Ecol. 2000, 69, 703–713. [Google Scholar] [CrossRef]
- Kong, L.; Wang, J.; Han, W.; Cao, Z. Modeling Heterogeneity in Direct Infectious Disease Transmission in a Compartmental Model. Int. J. Environ. Res. Public Health 2016, 13, 253. [Google Scholar] [CrossRef] [PubMed]
- Hoch, T.; Fourichon, C.; Viet, A.F.; Seegers, H. Influence of the transmission function on a simulated pathogen spread within a population. Epidemiol. Infect. 2007, 136, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Huang, Z.; Zhou, H.; Anders, K.L.; Perkins, T.A.; Yin, W.; Li, Y.; Mu, D.; Chen, Q.; Zhang, Z.; et al. The changing epidemiology of dengue in China, 1990-2014: A descriptive analysis of 25 years of nationwide surveillance data. BMC Med. 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wu, D.; Zhou, H.; Zhang, H.; Guan, D.; He, X.; Cai, S.; Ke, C.; Lin, J. The epidemiological characteristics and genetic diversity of dengue virus during the third largest historical outbreak of dengue in Guangdong, China, in 2014. J. Infect. 2016, 72, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Jing, Q.; Spear, R.C.; Marshall, J.M.; Yang, Z.; Gong, P. Climate and the Timing of Imported Cases as Determinants of the Dengue Outbreak in Guangzhou, 2014: Evidence from a Mathematical Model. PLoS Negl. Trop. Dis. 2016, 10, e0004417. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Luo, Y.; Chen, Y.; Lamers, M.M.; Zhou, Q.; Yang, X.H.; Sanyal, S.; Mok, C.K.; Liu, Z.M. Clinical and epidemiological features of the 2014 large-scale dengue outbreak in Guangzhou city, China. BMC Infect. Dis. 2016, 16, 102. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Jiang, L.Y.; Xiao, X.C.; Di, B.; Jing, Q.L.; Wang, S.Y.; Tang, J.L.; Wang, M.; Tang, X.P.; Yang, Z.C. The dengue preface to endemic in mainland China: The historical largest outbreak by Aedes albopictus in Guangzhou, 2014. Infect. Dis. Poverty 2017, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Li, X.; Xiao, X.; Xu, Y.; Huang, M.; Yang, Z. Identification of Aedes albopictus larval index thresholds in the transmission of dengue in Guangzhou, China. J. Vector Ecol. 2015, 40, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Yiguan, W.; Xin, L.; Chengling, L.; Su, T.; Jianchao, J.; Yuhong, G.; Dongsheng, R.; Zhicong, Y.; Qiyong, L.; Fengxia, M. A Survey of Insecticide Resistance in Aedes albopictus (Diptera: Culicidae) During a 2014 Dengue Fever Outbreak in Guangzhou, China. J. Econ. Entomol. 2017, 110, 239–244. [Google Scholar] [PubMed]
- Samanta, S.; Pal, D.K.; Lohar, D.; Pal, B. Interpolation of climate variables and temperature modeling. Theretical Appl. Climatol. 2012, 107, 35–45. [Google Scholar] [CrossRef]
- Newton, E.; Reiter, P. A model of the transmission of dengue fever with an evaluation of the impact of ultra-low volume (ULV) insecticide applications on dengue epidemics. Am. J. Trop. Med. Hyg. 1992, 47, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Erickson, R.A.; Presley, S.M.; Allen, L.J.S.; Long, K.R.; Cox, S.B. A dengue model with a dynamic Aedes albopictus vector population. Ecol. Model. 2010, 221, 2899–2908. [Google Scholar] [CrossRef]
- Otero, M.; Barmak, D.H.; Dorso, C.O.; Solari, H.G.; Natiello, M.A. Modeling dengue outbreaks. Math. Biosci. 2011, 232, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Oki, M.; Sunahara, T.; Hashizume, M.; Yamamoto, T. Optimal Timing of Insecticide Fogging to Minimize Dengue Cases: Modeling Dengue Transmission among Various Seasonalities and Transmission Intensities. PLoS Negl. Trop. Dis. 2011, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.; Johansson, M.A. The Incubation Periods of Dengue Viruses. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A.; et al. Dengue Viremia Titer, Antibody Response Pattern, and Virus Serotype Correlate with Disease Severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Blumberg, S.; Lloyd-Smith, J.O. Inference of R0 and Transmission Heterogeneity from the Size Distribution of Stuttering Chains. PLoS Comput. Biol. 2013, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.W.; Comrie, A.C.; Ernst, K. Climate and dengue transmission: Evidence and implications. Environm. Health Perspect. 2013, 121, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Macoris, M.L.G.; Galvani, K.C.; Andrighetti, M.T.M.; Wanderley, D.M.V. Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiol. Infect. 2009, 137, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; L’Ambert, G.; Lacour, G.; Benoît, R.; Demarchi, M.; Cros, M.; Cailly, P.; Aubry-Kientz, M.; Balenghien, T.; Ezanno, P. A Rainfall- and Temperature-Driven Abundance Model for Aedes albopictus Populations. Int. J. Environ. Res. Public Health 2013, 10, 1698–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Dengue Control: Environmental Management. Available online: http://www.who.int/denguecontrol/control_strategies/environmental_management/en/ (accessed on 20 March 2018).
- Barrera, R.; Amador, M.; MacKay, A.J. Population Dynamics of Aedes aegypti and Dengue as Influenced by Weather and Human Behavior in San Juan, Puerto Rico. PLoS Negl. Trop. Dis. 2011, 5, e1378. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, N.B.; Thomas, S.M.; Fischer, D.; Beierkuhnlein, C. Extrinsic Incubation Period of Dengue: Knowledge, Backlog, and Applications of Temperature Dependence. PLoS Negl. Trop. Dis. 2013, 7, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Barrera, R.; Amador, M.; Clark, G.G. Ecological factors influencing Aedes aegypti (Diptera: Culicidae) productivity in artificial containers in Salinas, Puerto Rico. J. Med. Entomol. 2006, 43, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Vikram, K.; Nagpal, B.N.; Pande, V.; Srivastava, A.; Gupta, S.K.; Anushrita, V.P.; Singh, H.; Saxena, R.; Tuli, N.R.; et al. Comparison of Ae. aegypti breeding in localities of different socio-economic groups of Delhi, India. Int. J. Mosq. Res. 2015, 2, 83–88. [Google Scholar]
- Ferdousi, F.; Yoshimatsu, S.; Ma, E.; Sohel, N.; Wagatsuma, Y. Identification of Essential Containers for Aedes Larval Breeding to Control Dengue in Dhaka, Bangladesh. Trop. Med. Health 2015, 43, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Thavara, U.; Tawatsin, A.; Chansang, C.; Kong-ngamsuk, W.; Paosriwong, S.; Boon-Long, J.; Rongsriyam, Y.; Komalamisra, N. Larval occurrence, oviposition behavior and biting activity of potential mosquito vectors of dengue on Samui Island, Thailand. J. Vector Ecol. 2001, 26, 172–180. [Google Scholar] [PubMed]
- Becker, N.; Petrić, D.; Boase, C.; Lane, J.; Zgomba, M.; Dahl, C.; Kaiser, A. Biology of Mosquitoes. In Mosquitoes and Their Control; Springer: Boston, MA, USA, 2003; pp. 9–27. [Google Scholar] [CrossRef]
- Cailly, P.; Tran, A.; Balenghien, T.; L’Ambert, G.; Toty, C.; Ezanno, P. A climate-driven abundance model to assess mosquito control strategies. Ecol. Model. 2012, 227, 7–17. [Google Scholar] [CrossRef]
- Juliano, S.A. Population dynamics. J. Am. Mosq. Control Assoc. 2007, 23, 265–275. [Google Scholar] [CrossRef]
- Lucas, E.A., Jr.; Romoser, W.S. The energetic costs of diving in Aedes aegypti and Aedes albopictus pupae. J. Am. Mosq. Control Assoc. 2001, 17, 56–60. [Google Scholar] [PubMed]
- Walsh, R.K.; Facchinelli, L.; Ramsey, J.M.; Bond, J.G.; Gould, F. Assessing the impact of density dependence in field populations of Aedes aegypti. J. Vector Ecol. 2011, 36, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of Chikungunya and Dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef] [PubMed]
- MathWorks. Global Optimization Toolbox. Available online: http://cn.mathworks.com/help/gads/index.html (accessed on 20 March 2018).
- Ugray, Z.; Lasdon, L.; Plummer, J.; Glover, F.; Kelly, J.; Marti, R. Scatter search and local NLP solvers: A multistart framework for global optimization. INFORMS J. Comput. 2007, 19, 328–340. [Google Scholar] [CrossRef]
- MathWorks. goodnessOfFit: Goodness of Fit between Test and Reference Data. Available online: http://cn.mathworks.com/help/ident/ref/goodnessoffit.html (accessed on 20 March 2018).
- Allen, L.J.S.; Burgin, A.M. Comparison of deterministic and stochastic SIS and SIR models in discrete time. Math. Biosci. 2000, 163, 1–33. [Google Scholar] [CrossRef]
- Lin, H.; Liu, T.; Song, T.; Lin, L.; Xiao, J.; Lin, J.; He, J.; Zhong, H.; Hu, W.; Deng, A.; et al. Community Involvement in Dengue Outbreak Control: An Integrated Rigorous Intervention Strategy. PLoS Negl. Trop. Dis. 2016, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Favier, C.; Degallier, N.; Rosa-Freitas, M.G.; Boulanger, J.P.; Costa Lima, J.R.; Luitgards-Moura, J.F.; Menkes, C.E.; Mondet, B.; Oliveira, C.; Weimann, E.T.S.; et al. Early determination of the reproductive number for vector-borne diseases: The case of dengue in Brazil. Trop. Med. Int. Health 2006, 11, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Otero, M.; Solari, H.G.; Schweigmann, N. A stochastic population dynamics model for Aedes aegypti: Formulation and application to a city with temperate climate. Bull. Math. Biol. 2006, 68, 1945–1974. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, M.; Shu, B.; Chen, X.Q.; Luo, L.; Wang, J.Y.; Cen, Y.Z.; Anderson, B.D.; Merrill, M.M.; Merrill, H.R.; et al. Evaluation of Inapparent Dengue Infections During an Outbreak in Southern China. PLoS Negl. Trop. Dis. 2015, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue vaccine: WHO position paper, July 2016—Recommendations. Vaccine 2017, 35, 1200–1201. [Google Scholar]
- Favier, C.; Schmit, D.; Muller-Graf, C.D.M.; Cazelles, B.; Degallier, N.; Mondet, B.; Dubois, M.A. Influence of spatial heterogeneity on an emerging infectious disease: The case of dengue epidemics. Proc. R. Soc. B Biol. Sci. 2005, 272, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Pongsumpun, P.; Garcia Lopez, D.; Favier, C.; Torres, L.; Llosa, J.; Dubois, M.A. Dynamics of dengue epidemics in urban contexts. Trop. Med. Int. Health 2008, 13, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue Control: The Mosquito. Available online: http://www.who.int/denguecontrol/mosquito/en/ (accessed on 20 March 2018).
- Reiner, R.C.; Stoddard, S.T.; Scott, T.W. Socially structured human movement shapes dengue transmission despite the diffusive effect of mosquito dispersal. Epidemics 2014, 6, 30–36. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control (New Edition); WHO Press: Geneva, Switzerland, 2009. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Biological Meaning | Range | Value | Source |
|---|---|---|---|---|
| a | Average daily biting rate | 0.3–1 | 1 | [8] |
| Transmission probability from vector to human per bite | 0.1–0.75 | 0.5 | [8] | |
| Transmission probability from human to vector per bite | 0.5–1 | 0.75 | [8] | |
| Human life expectancy (years) | - | 75 | Assumed | |
| Average lifespan of mosquitoes (days) | 4–50 | 21 | [8] | |
| Intrinsic incubation period (IIP, days) | 4–10 | 7 | [41] | |
| Extrinsic incubation period (EIP, days) | 8–12 | 10 | [8] | |
| Infectious period (days) | 1–7 | 4.5 | [42] |
| Parameter | Biological Meaning | Values |
|---|---|---|
| Diapause | 1 in Mar. 15 to Oct. 25; 0 otherwise ([31]) | |
| Eggs per gonotrophic cycle (per female) | Temperature dependent | |
| 1/duration for gonotrophic cycle (per day) | Temperature dependent | |
| Egg development rate | Temperature and precipitation dependent | |
| Larva development rate | Temperature and precipitation dependent | |
| Development rate of pupae to emerging adults | Temperature dependent | |
| Egg mortality rate | 0.05 ([47]) | |
| Mortality for larva | Temperature and density dependent | |
| Mortality for pupa | Temperature dependent | |
| Mortality rate during adult emergence | 0.1 ([47]) | |
| Mortality rate of adult mosquitoes | Temperature dependent | |
| Sex ratio of Aedes albopictus at emergence | 0.5 ([61]) | |
| Development rate of emerging adults (day) | 0.4 ([47]) | |
| Carrying capacity of mosquito larvae population | Precipitation and environment dependent | |
| Carrying capacity of mosquito pupae population | Precipitation and environment dependent | |
| Maximum carrying capacity for immature mosquitoes | Environment and density dependent | |
| Total population of adult female mosquitoes |
| Parameter | Biological Meaning | Optimized Value ± Std |
|---|---|---|
| The date on which an infectious human trigged the outbreak | 17 June 2014 ± 7 days | |
| Heterogeneity level in the 1st phase | ||
| Heterogeneity level in the 2nd phase | ||
| Transmission probability from vector to human per bite | 0.414 | |
| Transmission probability from human to vector per bite | 0.682 | |
| Maximum carrying capacity of immature mosquitoes | ± 14665 | |
| Intrinsic incubation period (IIP, days) | 7.853 ± 0.22 | |
| Infectious period (days) | 4.746 ± 0.135 | |
| Estimated cumulative reported cases | 37,733 |
| Scenario | Reported Total Infections | Change |
|---|---|---|
| Reported | 37,420 | - |
| Fitted | 37,733 | - |
| decreased by 10% | 18,758 | Decreased by 50.29% |
| increased by 10% | 61,684 | Increased by 63.47% |
| Intervention 10 days earlier | 14,831 | Decreased by 60.69% |
| Intervention 10 days delayed | 84,746 | Increased by 124.59% |
| Vaccinating 10% | 21,232 | Decreased by 43.73% |
| Vaccinating 20% | 11,563 | Decreased by 69.36% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, L.; Wang, J.; Li, Z.; Lai, S.; Liu, Q.; Wu, H.; Yang, W. Modeling the Heterogeneity of Dengue Transmission in a City. Int. J. Environ. Res. Public Health 2018, 15, 1128. https://doi.org/10.3390/ijerph15061128
Kong L, Wang J, Li Z, Lai S, Liu Q, Wu H, Yang W. Modeling the Heterogeneity of Dengue Transmission in a City. International Journal of Environmental Research and Public Health. 2018; 15(6):1128. https://doi.org/10.3390/ijerph15061128
Chicago/Turabian StyleKong, Lingcai, Jinfeng Wang, Zhongjie Li, Shengjie Lai, Qiyong Liu, Haixia Wu, and Weizhong Yang. 2018. "Modeling the Heterogeneity of Dengue Transmission in a City" International Journal of Environmental Research and Public Health 15, no. 6: 1128. https://doi.org/10.3390/ijerph15061128
APA StyleKong, L., Wang, J., Li, Z., Lai, S., Liu, Q., Wu, H., & Yang, W. (2018). Modeling the Heterogeneity of Dengue Transmission in a City. International Journal of Environmental Research and Public Health, 15(6), 1128. https://doi.org/10.3390/ijerph15061128