Mercury Contamination in Riverine Sediments and Fish Associated with Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
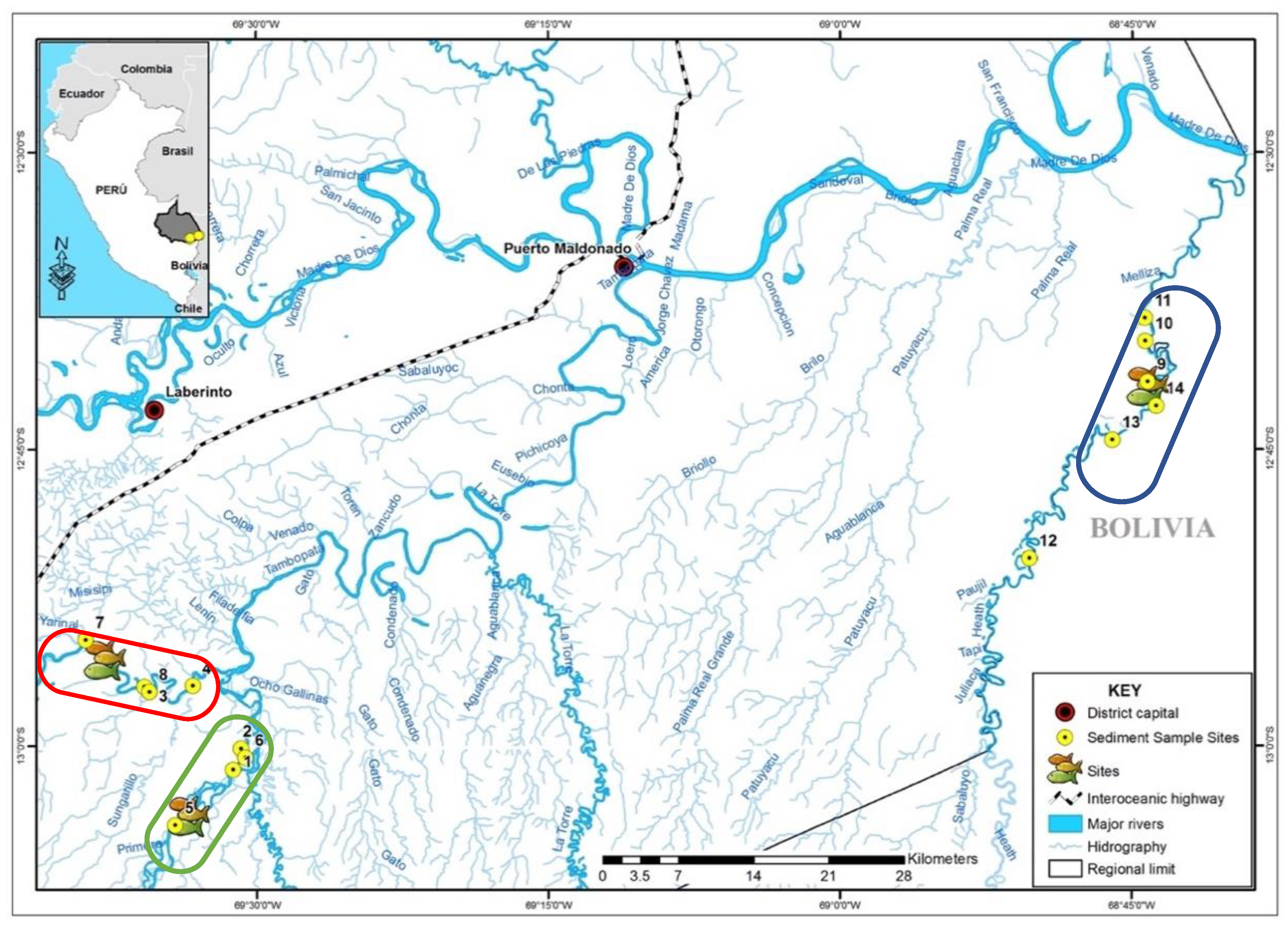
2.1. Site Description
2.2. Sampling and Sample Processing Procedures
2.3. Chemical Analyses
2.4. Statistical Analyses
3. Results
3.1. Field Measurements and Physical Observations
3.2. Total Hg and % C in Sediment
3.3. Hg in Fish
3.3.1. Fish Length
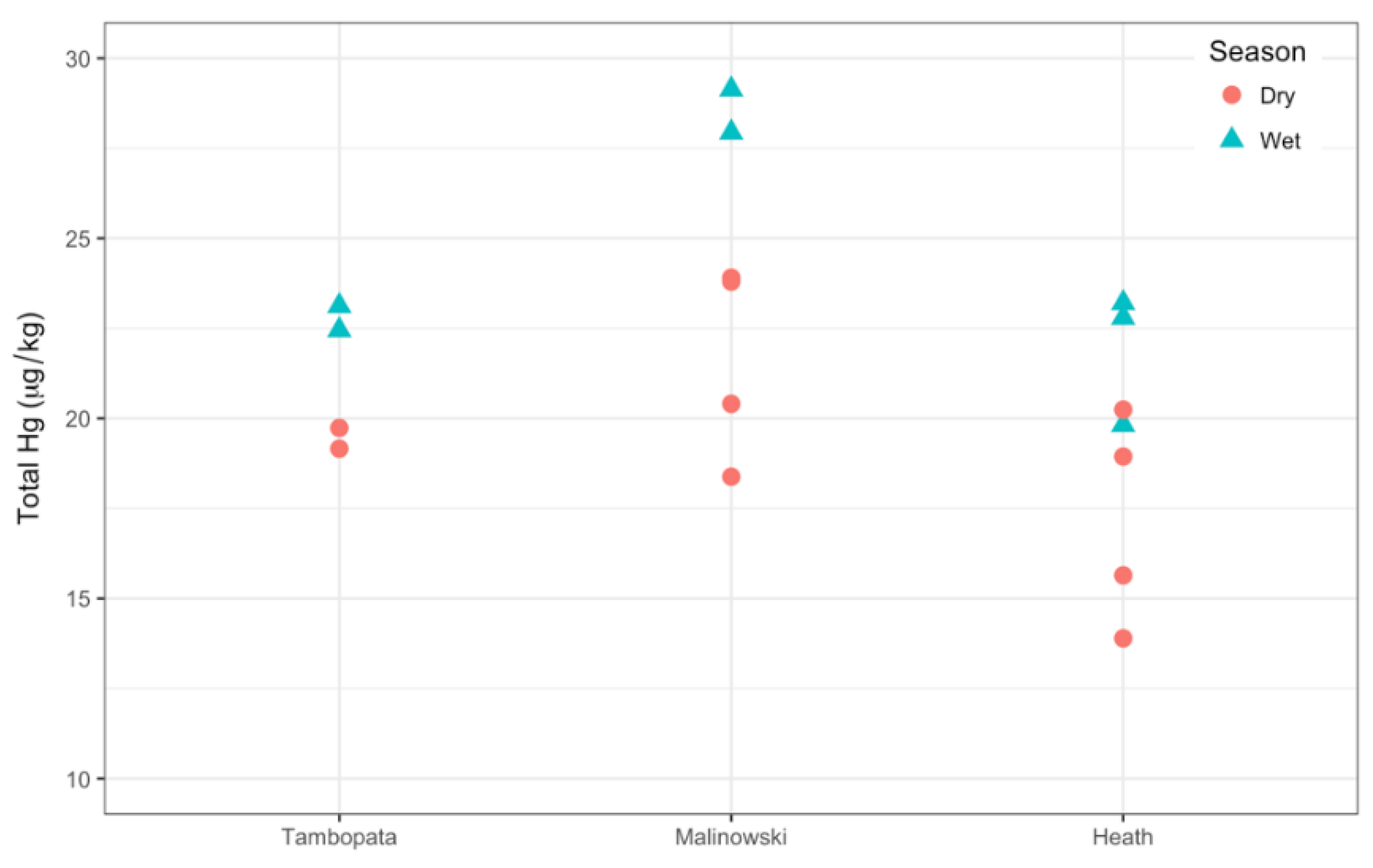
3.3.2. Wet vs. Dry Season
3.3.3. Fish Isotopic Values (δ15N and δ13C)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Exposure to Mercury: A Major Public Health Concern; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a global pollutant: Sources, pathways, and effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef] [PubMed]
- Sundseth, K.; Pacyna, J.M.; Pacyna, E.G.; Pirrone, N.; Thorne, R.J. Global Sources and Pathways of Mercury in the Context of Human Health. Int. J. Environ. Res. Public Health 2017, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Telmer, K.H.; Veiga, M.M. World emissions of mercury from artisanal and small scale gold mining. In Mercury Fate and Transport in the Global Atmosphere; Mason, R., Pirrone, N., Eds.; Springer: Boston, MA, USA, 2009; pp. 131–172. ISBN 978-0-387-93958-2. [Google Scholar]
- United Nations Environment Programme (UNEP). Global Mercury Assessment 2013: Sources, Emissions, Releases, and Environmental Transport; UNEP: Geneva, Switzerland, 2013.
- Driscoll, C.T.; Han, Y.-J.; Chen, C.Y.; Evers, D.C.; Lambert, K.F.; Holsen, T.M.; Kamman, N.C.; Munson, R.K. Mercury Contamination in Forest and Freshwater Ecosystems in the Northeastern United States. BioScience 2007, 57, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Mergler, D.; Anderson, H.A.; Chan, L.H.M.; Mahaffey, K.R.; Murray, M.; Sakamoto, M.; Stern, A.H. Methylmercury Exposure and Health Effects in Humans: A Worldwide Concern. Ambio 2007, 36, 3–11. [Google Scholar] [CrossRef]
- Eagles-Smith, C.A.; Silbergeld, E.K.; Basu, N.; Bustamante, P.; Diaz-Barriga, F.; Hopkins, W.A.; Kidd, K.A.; Nyland, J.F. Modulators of mercury risk to wildlife and humans in the context of rapid global change. Ambio 2018, 47, 170–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malm, O.; Branches, F.J.P.; Akagi, H.; Castro, M.B.; Pfeiffer, W.C.; Harada, M.; Bastos, W.R.; Kato, H. Mercury and methylmercury in fish and human hair from the Tapajós river basin, Brazil. Sci. Total Environ. 1995, 175, 141–150. [Google Scholar] [CrossRef]
- Bastos, W.R.; Gomes, J.P.; Oliveira, R.C.; Almeida, R.; Nascimento, E.L.; Bernardi, J.V.; de Lacerda, L.D.; da Silveira, E.G.; Pfeiffer, W.C. Mercury in the environment and riverside population in the Madeira River Basin, Amazon, Brazil. Sci. Total Environ. 2006, 368, 344–351. [Google Scholar] [CrossRef] [PubMed]
- United States Geological Survey (USGS). Mineral Commodity Summaries 2016; USGS: Reston, VA, USA, 2016; p. 202. [Google Scholar]
- World Atlas. Available online: https://www.worldatlas.com/articles/top-14-gold-producers-in-the-world.html (accessed on 28 February 2018).
- Investing News Network. Available online: https://investingnews.com/daily/resource-investing/precious-metals-investing/gold-investing/top-gold-producing-countries/ (accessed on 28 February 2018).
- Brack, A.; Ipenza, C.; Alvarez, J.; Sotero, V. Minería Aurífera en Madre de Dios y Contaminación con Mercurio—Una Bomba de Tiempo, 1st ed.; Editorial Súper Gráfica, E.I.R.L., Ed.; Ministerio del Ambiente: Lima, Peru, 2011; ISBN 978-612-45818-7-8. [Google Scholar]
- Asner, G.P.; Llactayo, W.; Tupayachi, R.; Luna, E.R. Elevated rates of gold mining in the Amazon revealed through high-resolution monitoring. Proc. Natl. Acad. Sci. USA 2013, 110, 18454–18459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asner, G.P.; Tupayachi, R. Accelerated losses of protected forests from gold mining in the Peruvian Amazon. Environ. Res. Lett. 2017, 12, 094004. [Google Scholar] [CrossRef]
- Pachas, V.H. Historia de una Incertidumbre: Hábitat, Conflicto y Poder en la Minería Artesanal de oro en el Perú; Conservación y Desarrollo; Earth First SAC: Lima, Perú, 2013; p. 292. [Google Scholar]
- Valencia, L. Políticas de Pequeña Minería y Deforestación: El Caso de Madre de Dios; SPDA: Lima, Peru, 2016. [Google Scholar]
- Ashe, K. Elevated mercury concentrations in humans of Madre de Dios, Peru. PLoS ONE 2012, 7, e33305. [Google Scholar] [CrossRef] [PubMed]
- Gerson, J.R.; Driscoll, C.T.; Hsu-Kim, H.; Berhhardt, E.S. Senegalese artisanal gold mining leads to elevated total mercury and methylmercury concentrations in soils, sediments, and rivers. Elementa Sci. Anthr. 2018, 6. [Google Scholar] [CrossRef]
- Alpers, C.N.; Hunerlach, M.P.; May, J.T.; Hothem, R.L.; Taylor, H.E.; Antweiler, R.C.; De Wild, J.F.; Lawler, D.A. Geochemical characterization of water, sediment, and biota affected by mercury contamination and acidic drainage from historical gold mining, Greenhorn Creek, Nevada County, California, 1999–2001. In U.S. Geological Survey Scientific Investigations Report 2004-5251; U.S. Department of Interior and the U.S. Geological Survey: Reston, VA, USA, 2005; p. 278. [Google Scholar]
- Tollefson, L.; Cordle, F. Methylmercury in Fish: A Review of Residue Levels, Fish Consumption and Regulatory Action in the United States. Environ. Health Perspect. 1986, 68, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Bloom, N.S. On the Chemical Form of Mercury in Edible Fish and Marine Invertabrate Tissue. Can. J. Fish. Aquat. Sci. 1992, 49, 1010–1017. [Google Scholar] [CrossRef]
- United States Geological Survey (USGS). Mercury Contamination of Aquatic Ecosystems; USGS: Reston, VA, USA, 1995. Available online: https://pubs.usgs.gov/fs/1995/fs216-95/pdf/fs21695.pdf (accessed on 25 July 2018).
- Tello, S.; Bayley, P.B. La pesquería comercial de Loreto con énfasis en el análisis de la relación entre captura y esfuerzo pesquero de la flota comercial de Iquitos, cuenca del Amazonas (Perú). Folia Amaz. 2001, 12, 123–139. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (USEPA). Mercury Study Report to Congress, Volume III: Fate and Transport of Mercury Fate in the Environment; USEPA: Washington, DC, USA, 1997.
- Diringer, S.E.; Feingold, B.J.; Ortiz, E.J.; Gallis, J.A.; Araujo-Flores, J.M.; Berky, A.; Pan, W.K.; Hsu-Kim, H. River transport of mercury from artisanal and small-scale gold mining and risks for dietary mercury exposure in Madre de Dios, Peru. Environ. Sci. Process Impacts 2015, 17, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Centro de Atención Médica Especializada y Preventiva (CAMEP). Mercury in Madre de Dios—Mercury Concentrations in Fish and Humans in Puerto Maldonado; CAMEP: Vista Alegre, Panama, 2013; Available online: https://dge.carnegiescience.edu/research/CAMEP/CAMEP%20Research%20Brief%20-%20Puerto%20Maldonado%20English%20-%20FINAL.pdf (accessed on 26 April 2018).
- Luis Fernández, V.G. Niveles del Mercurio en Peces de Madre de Dios. Available online: http://www.minam.gob.pe/mineriailegal/wp-content/uploads/sites/43/2013/10/Carnegie-mercurio-Madre-de-Dios.pdf (accessed on 26 April 2018).
- Roach, K.A.; Jacobsen, N.F.; Fiorello, C.V.; Stronza, A.; Winemiller, K.O. Gold Mining and Mercury Bioaccumulation in a Floodplain Lake and Main Channel of the Tambopata River, Perú. J. Environ. Prot. 2013, 4, 51–60. [Google Scholar] [CrossRef]
- Langeland, A.L.; Hardin, R.D.; Neitzel, R.L. Mercury Levels in Human Hair and Farmed Fish near Artisanal and Small-Scale Gold Mining Communities in the Madre de Dios River Basin, Peru. Int. J. Environ. Res. Public Health 2017, 14, 302. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, L.; Ortiz, E.J.; Feingold, B.; Berky, A.; Diringer, S.; Morales, A.M.; Jurado, E.R.; Hsu-Kim, H.; Pan, W. Spatial, Temporal, and Dietary Variables Associated with Elevated Mercury Exposure in Peruvian Riverine Communities Upstream and Downstream of Artisanal and Small-Scale Gold Mining. Int. J. Environ. Res. Public Health 2017, 14, 1582. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Brush, M.; Rydberg, J.; Gamboa, N.; Storch, I.; Biester, H. Is mercury from small-scale gold mining prevalent in the southeastern Peruvian Amazon? Environ. Pollut. 2016, 218, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Myers, N. Threatened Biotas: “Hot Spots” in Tropical Forests. Environmentalist 1988, 8, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M.; Dinerstein, E. The Global 200: A Representation Approach to Conserving the Earth’s Most Biologically Valuable Ecoregions. Conserv. Biol. 1998, 12, 502–515. [Google Scholar] [CrossRef]
- Jensen, K.E.; Naik, N.N.; O’Neal, C.; Salmon-Mulanovich, G.; Riley-Powell, A.R.; Lee, G.O.; Hartinger, S.M.; Bausch, D.G.; Paz-Soldan, V.A. Small scale migration along the interoceanic highway in Madre de Dios, Peru: An exploration of community perceptions and dynamics due to migration. BMC Int. Health Hum. Rights 2018, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.E.; Kullander, S.O.; Ferraris, C.J.J. Check List of the Freshwater Fishes of South and Central America; EDIPUCRS: Porto Alegre, Brasil, 2003; p. 742. ISBN 85-7430-361-5. [Google Scholar]
- Van der Sleen, P. Albert, J.S. Field Guide to the Fishes of the Amazon, Orinoco, & Guianas; Princeton University Press: Princeton, NJ, USA, 2018; ISBN 978-0-691-17074-9. [Google Scholar]
- Baker, R.F.; Blanchfield, P.J.; Paterson, M.J.; Flett, R.J.; Wesson, L. Evaluation of Nonlethal Methods for the Analysis of Mercury in Fish Tissue. Am. Fish. Soc. 2004, 133, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Peterson, S.A.; Van Sickle, J.; Hughes, R.M.; Schacher, J.A.; Echols, S.F. A biopsy procedure for determining filet and predicting whole-fish mercury concentration. Arch. Environ. Contam. Toxicol. 2005, 48, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Ackerson, J.R.; McKee, M.J.; Schmitt, C.J.; Brumbaugh, W.G. Implementation of a non-lethal biopsy punch monitoring program for mercury in smallmouth bass, Micropterus dolomieu Lacepede, from the Eleven Point River, Missouri. Bull. Environ. Contam. Toxicol. 2014, 92, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.J.; Stevens, T.F.; Lee, S.Y. Assessing the suitability of a non-lethal biopsy punch for sampling fish muscle tissue. Fish Physiol. Biochem. 2016, 42, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency (USEPA). Method 7473 (SW-846): Mercury in Solids and Solutions by Thermal Decomposition, Amalgamation, and Atomic Absorption Spectrophotometry; USEPA: Washington, DC, USA, 1998.
- Zimmerman, C.F.; Keefe, C.W.; Bashe, J. Method 440.0 Determination of Carbon and Nitrogen in Sediments and Particulatesof Estuarine/Coastal Waters Using Elemental Analysis; EPA/600/R-15/009; U.S. Environmental Protection Agency: Washington, DC, USA, 1997.
- Stewart, A.R.; Saiki, M.K.; Kuwabara, J.S.; Alpers, C.N.; Marvin-DiPasquale, M.; Krabbenhoft, D.P. Influence of plankton mercury dynamics and trophic pathways on mercury concentrations of top predator fish of a mining-impacted reservoir. Can. J. Fish. Aquat. Sci. 2008, 65, 2351–2366. [Google Scholar] [CrossRef] [Green Version]
- Post, D.M. Using Stable Isotopes to Estimate Trophic Position: Models, Methods, and Assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Carscallen, W.M.A.; Vandenberg, K.; Lawson, J.M.; Martinez, N.D.; Romanuk, T.N. Estimating trophic position in marine and estuarine food webs. Ecosphere 2012, 3. [Google Scholar] [CrossRef]
- Vander Zanden, J.; Rasmussen, J. Variation in d15N and d13C trophic fractionation: Implications for aquatic food web studies. Limnol. Oceanogr. 2001, 46, 2061–2066. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between d15N and animal age. Geochim. Cosmochim. Acta 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W. H. Freeman and Company: New York, NY, USA, 2011; ISBN 0-7167-8604-4. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- United States Environmental Protection Agency. Water Quality Criterion for the Protection of Human Health: Methylmercury Final; EPA: Washington, DC, USA, 2001.
- Malm, O.; Pfeiffer, W.C.; Souza, C.M.M.; Reuther, R. Mercury pollution due to gold mining in the Madeira River Basin, Brazil. Ambio 1990, 19, 11–15. [Google Scholar]
- Lacerda, L.D.; Marins, R.V.; Souza, C.M.M.; Rodrigues, S.; Pfeiffer, W.C.; Bastos, W.R. Mercury dispersal in water, sediments and aquatic biota of a gold mining tailings drainage in Pocone’, Brazil. Water Air Soil Pollut. 1991, 55, 283–294. [Google Scholar] [CrossRef]
- Malm, O.; Castro, M.B.; Bastos, W.R.; Branches, F.J.P.; Guimarães, J.R.D.; Zuffo, C.E.; Pfeiffer, W.C. An assessment of Hg pollution in different goldmining areas, Amazon Brazil. Sci. Total Environ. 1995, 175, 127–140. [Google Scholar] [CrossRef]
- Lechler, P.J.; Miller, J.R.; Lacerda, L.D.; Vinson, D.; Bonzongo, J.-C.; Lyons, W.B.; Warwick, J.J. Elevated mercury concentrations in soils, sediments, water, and fish of the Madeira River basin, Brazilian Amazon: A function of natural enrichments? Sci. Total Environ. 2000, 260, 87–96. [Google Scholar] [CrossRef]
- Eagles-Smith, C.A.; Ackerman, J.T.; Willacker, J.J.; Tate, M.T.; Lutz, M.A.; Fleck, J.A.; Stewart, A.R.; Wiener, J.G.; Evers, D.C.; Lepak, J.M.; et al. Spatial and temporal patterns of mercury concentrations in freshwater fish across the Western United States and Canada. Sci. Total Environ. 2016, 568, 1171–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eagles-Smith, C.A.; Wiener, J.G.; Eckley, C.S.; Willacker, J.J.; Evers, D.C.; Marvin-DiPasquale, M.; Obrist, D.; Fleck, J.A.; Aiken, G.R.; Lepak, J.M.; et al. Mercury in western North America: A synthesis of environmental contamination, fluxes, bioaccumulation, and risk to fish and wildlife. Sci. Total Environ. 2016, 568, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Wiener, J.G.; Krabbenhoft, D.P.; Heinz, G.H.; Scheuhammer, A.M. Ecotoxicology of Mercury. In Handbook of Ecotoxicology, 2nd ed.; Hoffman, D.J., Rattner, B.A., Burton, G.A., Jr., Cairns, J., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 409–463. ISBN 1-56670-546-0. [Google Scholar]
- Davis, J.A.; Ross, J.R.; Bezalel, S.; Sim, L.; Bonnema, A.; Ichikawa, G.; Heim, W.A.; Schiff, K.; Eagles-Smith, C.A.; Ackerman, J.T. Hg concentrations in fish from coastal waters of California and Western North America. Sci. Total Environ. 2016, 568, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.C.; De Souza, J.; Dorea, J.G.; Jardim, W.F.; Fadini, P.S. Mercury Biomagnification in a Tropical Black Water, Rio Negro, Brazil. Arch. Environ. Contam. Toxicol. 2003, 45, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.; Hidalgo, M.; Trevejo, G.; Correa, E.; Cortijo, A.M.; Meza, V.; Espino, J. Lista Anotada de los Peces de Aguas Continentales del Perú: Estado Actual del Conocimiento, Distribución, Usos y Aspectos de Conservación; Ministerio del Ambiente: Lima, Peru, 2012; p. 56. ISBN 978-612-46053-2-1.
- Bastos, W.R.; Dorea, J.G.; Bernardi, J.V.; Lauthartte, L.C.; Mussy, M.H.; Lacerda, L.D.; Malm, O. Mercury in fish of the Madeira river (temporal and spatial assessment), Brazilian Amazon. Environ. Res. 2015, 140, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.M.; Hecky, R.E.; Wandera, S.B. Stable Isotope Analyses of Food Web Structure and Fish Diet in Napoleon and Winam Gulfs, Lake Victoria, East Africa. J. Great Lakes Res. 2003, 29, 243–257. [Google Scholar] [CrossRef]
- Araujo-Lima, C.A.; Forsberg, B.R.; Victoria, R.; Martinelli, L. Energy Sources for Detritivorous Fishes in the Amazon. Science 1986, 234, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Wantzen, K.M.; Machado, F.D.A.; Voss, M.; Boriss, H.; Junk, W.J. Seasonal isotopic shifts in fish of the Pantanal wetland, Brazil. Aquat. Sci. 2002, 64, 239–251. [Google Scholar] [CrossRef]
- Mol, J.; Ouboter, P.E. Downstream Effects of Erosion from Small-Scale Gold Mining on the Instream Habitat and Fish Community of a Small Neotropical Rainforest Stream. Conserv. Biol. 2004, 18, 201–214. [Google Scholar] [CrossRef]
- Aksnes, D.L.; Nejstgaard, J.; Sædberg, E.; Sørnes, T. Optical control of fish and zooplankton populations. Limnol. Oceanogr. 2004, 49, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Wantzen, K.; Mol, J. Soil Erosion from Agriculture and Mining: A Threat to Tropical Stream Ecosystems. Agriculture 2013, 3, 660–683. [Google Scholar] [CrossRef]
- Lujan, N.K.; Roach, K.A.; Jacobsen, D.; Winemiller, K.O.; Vargas, V.M.; Ching, V.R.; Maestre, J.A.; Ladle, R. Aquatic community structure across an Andes-to-Amazon fluvial gradient. J. Biogeogr. 2013, 40, 1715–1728. [Google Scholar] [CrossRef]
- Slotton, D.G.; Ayers, S.M.; Suchanek, T.H.; Weyand, R.D.; Liston, A.M.; MacDonald, C.; Nelson, D.C.; Johnson, B. The Effects of Wetland Restoration on the Production and Bioaccumulation of Methylmercury in the Sacramento-San Joaquin Delta, California: Draft Final Report; CALFED: Davis, CA, USA, 2002; p. 48. Available online: http://loer.tamug.edu/calfed/Report/DraftFinal/UCD_Delta_Report.pdf (accessed on 25 July 2018).
- Slotton, D.G.; Ayers, S.M.; Suchanek, T.H.; Weyand, R.D.; Liston, A.M. Mercury Bioaccumulation and Trophic Transfer in the Cache Creek Watershed of California, in Relation to Diverse Aqueous Mercury Exposure Conditions. Assessment of Ecological and Human Health Impacts of Mercury in the San Francisco Bay-Delta Watershed. A CALFED Bay-Delta Program Project. 2004, p. 74. Available online: http://loer.tamug.edu/calfed/Report/Final/UCDavis_Cache_Bio_Final.pdf (accessed on 25 July 2018).




{kind=link}
{kind=link}
{kind=link}
| River | Season | Site (M/T) | pH | Temperature (°C) | Dissolved Oxygen (mg/L) | NO3− (mg N/L) | Turbidity (NTU) |
|---|---|---|---|---|---|---|---|
| Tambopata | Wet | M | 7.5 | 26.3 (0.41) | 7.4 (0.04) | 0.29 (0.06) | 581 (362) |
| Wet | T | 7.6 | 27.5 | 7.0 | 0.26 | 215 | |
| Dry | M | 7.4 (0.23) | 25.7 (0.28) | 7.6 (0.17) | 0.05 (0.03) | 42 (31) | |
| Dry | T | 7.3 | 24.6 | 7.4 | 0.03 | 12 | |
| Malinowski | Wet | M | 7.1 (0.42) | 28.1 (0.55) | 6.9 (0.17) | 0.25 (0.14) | 359 (169) |
| Wet | T | 6.3 | 24.8 | 6.7 | 0.12 | 9 | |
| Dry | M | 6.9 (0.02) | 28.9 (1.6) | 6.3 (0.36) | 0.09 | 401 (56) | |
| Dry | T | NS 1 | NS | NS | NS | NS | |
| Heath | Wet | M | 6.5 (0.40) | 26.1 (1.1) | 5.0 (0.33) | 0.28 (0.12) | 305 (188) |
| Wet | T | 6.4 (0.02) | 25.1 (1.3) | 4.9 | 0.34 (0.03) | 79 (18) | |
| Dry | M | 7.2 (0.04) | 27.5 (1.9) | 6.2 (0.84) | 0.23 (0.02) | 644 (221) | |
| Dry | T | 6.6 | 25.9 | 6.2 | 0.06 | 33 |
| River | Season | Mean Hgfish (mg/kg) |
|---|---|---|
| Tambopata | Wet | 0.23 ± 0.23 |
| Dry | 0.22 ± 0.27 | |
| Malinowski | Wet | 0.18 ± 0.15 |
| Dry | 0.28 ± 0.24 | |
| Heath | Wet | 0.20 ± 0.22 |
| Dry | 0.19 ± 0.27 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez, G.; McCord, S.A.; Driscoll, C.T.; Todorova, S.; Wu, S.; Araújo, J.F.; Vega, C.M.; Fernandez, L.E. Mercury Contamination in Riverine Sediments and Fish Associated with Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru. Int. J. Environ. Res. Public Health 2018, 15, 1584. https://doi.org/10.3390/ijerph15081584
Martinez G, McCord SA, Driscoll CT, Todorova S, Wu S, Araújo JF, Vega CM, Fernandez LE. Mercury Contamination in Riverine Sediments and Fish Associated with Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru. International Journal of Environmental Research and Public Health. 2018; 15(8):1584. https://doi.org/10.3390/ijerph15081584
Chicago/Turabian StyleMartinez, Gerardo, Stephen A. McCord, Charles T. Driscoll, Svetoslava Todorova, Steven Wu, Julio F. Araújo, Claudia M. Vega, and Luis E. Fernandez. 2018. "Mercury Contamination in Riverine Sediments and Fish Associated with Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru" International Journal of Environmental Research and Public Health 15, no. 8: 1584. https://doi.org/10.3390/ijerph15081584
APA StyleMartinez, G., McCord, S. A., Driscoll, C. T., Todorova, S., Wu, S., Araújo, J. F., Vega, C. M., & Fernandez, L. E. (2018). Mercury Contamination in Riverine Sediments and Fish Associated with Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru. International Journal of Environmental Research and Public Health, 15(8), 1584. https://doi.org/10.3390/ijerph15081584