Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical
2.2. Animals and Treatments
2.3. Total RNA Extraction and Quantitative Real-Time PCR
2.4. Western-Blot Analysis
2.5. Statistical Analysis
3. Results
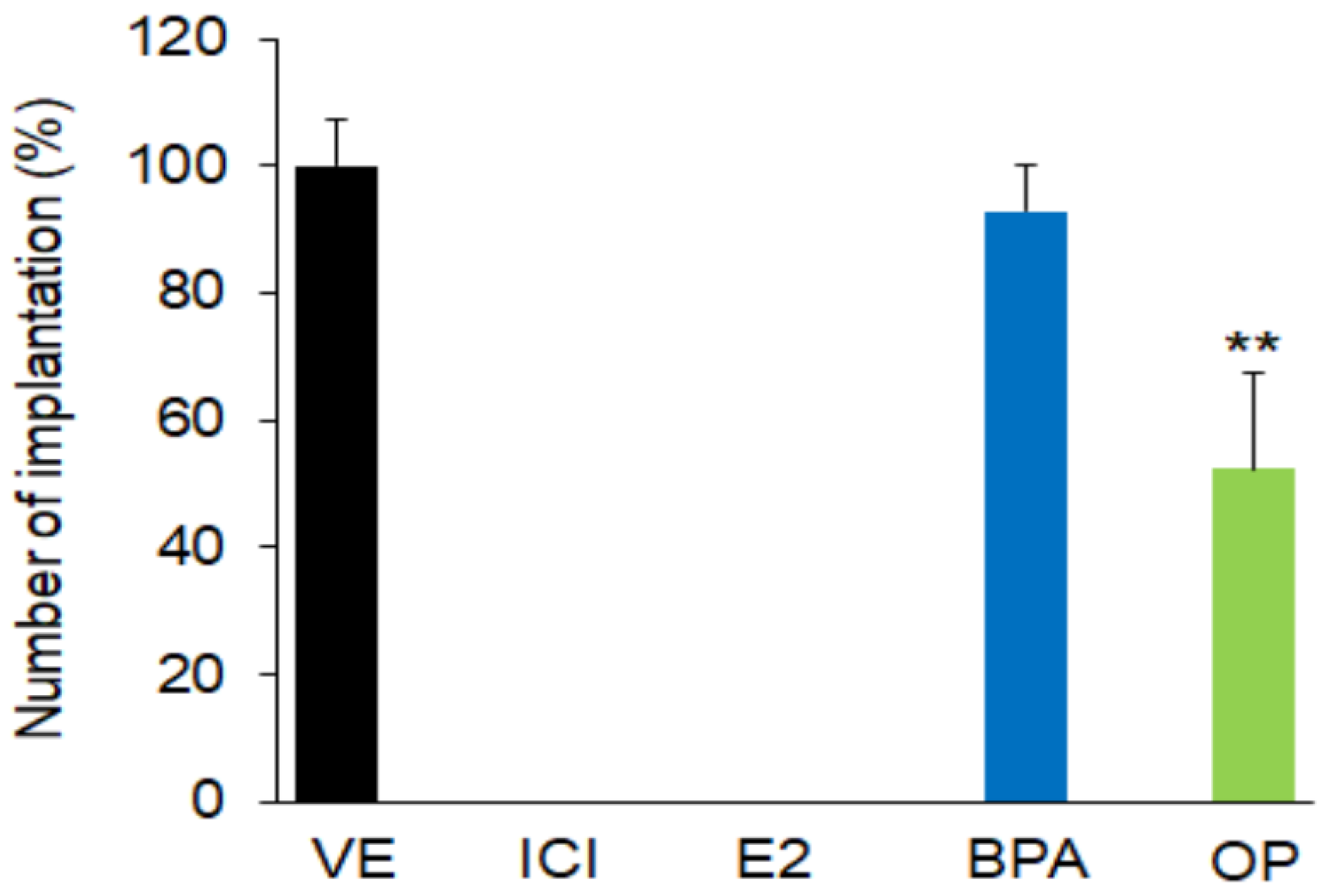
3.1. Effect of EDs on Blastocyst Implantation
3.2. Effects of BPA and OP on TRPV5, TRPV6, PMCA1, and NCX1 Expressions in Maternal Uterus and Implantation Sites
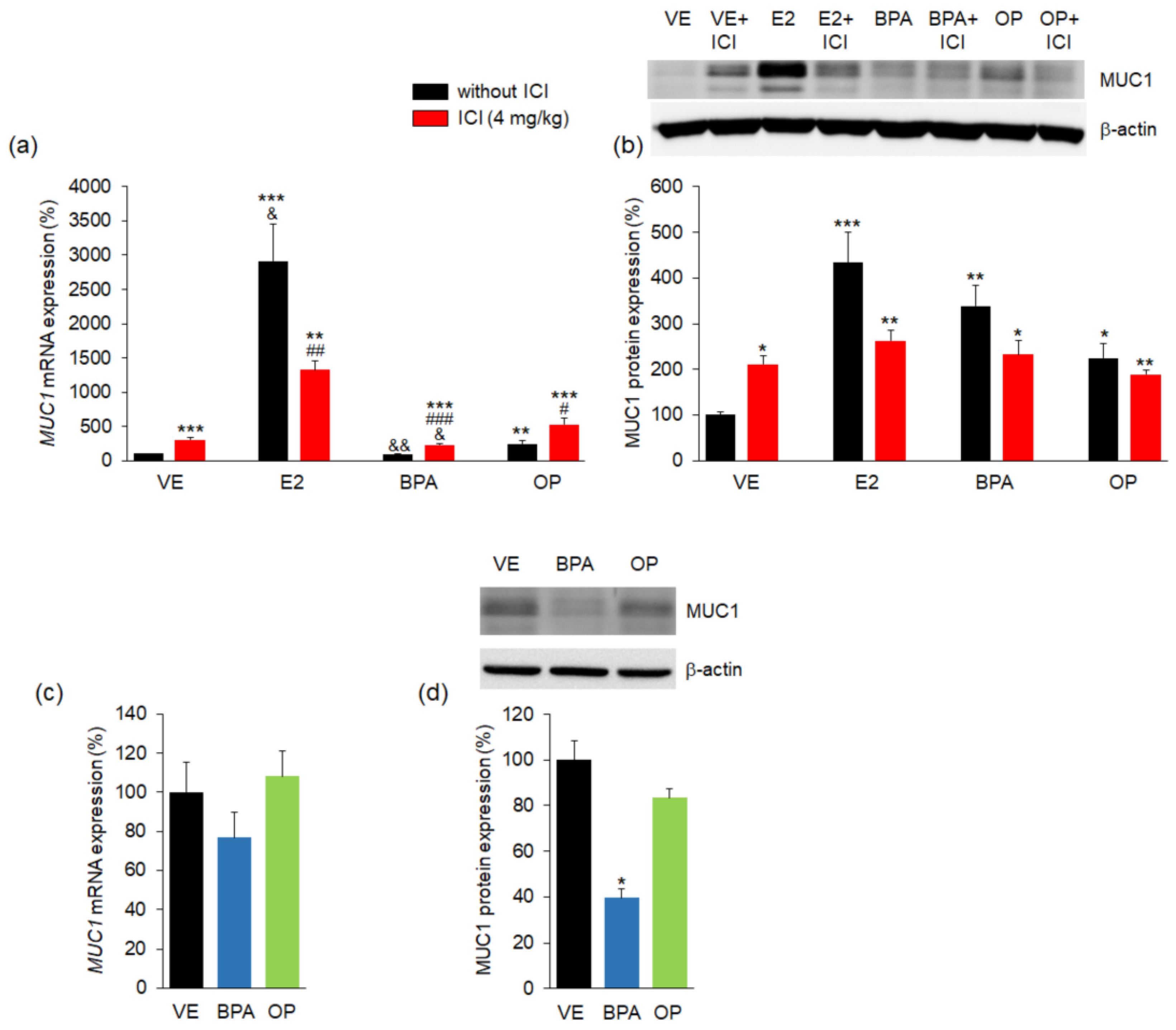
3.3. Effect of BPA and OP on MUC1 Expression in Maternal Uterus and Implantation Sites
3.4. Effects of BPA and OP on HOXA-10 and LIF Expression in Maternal Uterus and Implantation Sites
3.5. Effects of BPA and OP on Estrogen and Progesterone Receptor in Maternal Uterus and Implantation Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Bisphenol A | (BPA; 4,4′-(propane-2,2-diyl)diphenol) |
| OP | 4-tert-Octylphenol |
| E2 | 17β-Estradiol |
| P4 | Progesterone |
| ICI | ICI 182,780 |
| EDs | Endocrine disrupting chemicals |
| TRPV5 and TRPV6 | Transient receptor potential cation channel subfamily V (TRPV) member 5 and 6 |
| PMCA1 | Plasma membrane Ca2+-ATPase 1 |
| NCX1 | Na+/Ca2+ exchanger 1 |
| LIF | Leukemia inhibitory factor |
| HOXA-10 | homebox gene member 10 |
| MUC1 | Mucin 1 |
| ERα and ERβ | Estrogen receptor alpha and beta |
| PR- and PR-B | Progesterone receptor and progesterone receptor B |
| LOAEL | Lowest-observed-adverse-effect level |
References
- Dey, S.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev. 2004, 25, 341–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Paiva, P.; Menkhorst, E.; Salamonsen, L.; Dimitriadis, E. Leukemia inhibitory factor and interleukin-11: Critical regulators in the establishment of pregnancy. Cytokine Growth Factor Rev. 2009, 20, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Surveyor, G.A.; Gendler, S.J.; Pemberton, L.; Das, S.; Chakraborty, I.; Julian, J.; Pimental, R.; Wegner, C.; Dey, S.; Carson, D. Expression and steroid hormonal control of Muc-1 in the mouse uterus. Endocrinology 1995, 136, 3639–3647. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; Aplin, J.D.; Caballero-Campo, P.; O’Connor, J.E.; Martín, J.C.; Remohí, J.; Pellicer, A.; Simón, C. Human endometrial mucin MUC1 is up-regulated by progesterone and down-regulated in vitro by the human blastocyst. Biol. Reprod. 2001, 64, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.S.; Arici, A.; Olive, D.; Igarashi, P. HOXA10 is expressed in response to sex steroids at the time of implantation in the human endometrium. J. Clin. Investig. 1998, 101, 1379. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, M.; Lee, S.K. Role of endometrial immune cells in implantation. Clin. Exp. Reprod. Med. 2011, 38, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Marquardt, M.A.; Lech, J.J. Estrogenic effects of nonylphenol on pS2, ER and MUC1 gene expression in human breast cancer cells-MCF-7. Chem.-Biol. Interact. 1997, 104, 55–64. [Google Scholar] [CrossRef]
- Du, H.; Taylor, H.S. The role of Hox genes in female reproductive tract development, adult function, and fertility. Cold Spring Harbor Perspect. Med. 2016, 6, a023002. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L. Leukemia inhibitory factor and human embryo implantation. Ann. N. Y. Acad. Sci. 2004, 1034, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Hambartsoumian, E. Endometrial leukemia inhibitory factor (LIF) as a possible cause of unexplained infertility and multiple failures of implantation. Am. J. Reprod. Immunol. 1998, 39, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, Z.; Shokrzadeh, N.; Saidijam, M.; Sanoee, M.F. Semi-quantitative analysis of HOXA11, leukemia inhibitory factor and basic transcriptional element binding protein 1 mRNA expression in the mid-secretory endometrium of patients with endometriosis. Iran. Biomed. J. 2011, 15, 66. [Google Scholar] [PubMed]
- Cheng, T.-C.; Huang, C.-C.; Chen, C.-I.; Liu, C.-H.; Hsieh, Y.-S.; Huang, C.-Y.; Lee, M.-S.; Liu, J.-Y. Leukemia inhibitory factor antisense oligonucleotide inhibits the development of murine embryos at preimplantation stages. Biol. Reprod. 2004, 70, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, X.; Saravelos, S.H.; Liu, Y.; Huang, J.; Zhang, J.; Li, T.C. HOXA-10 and E-cadherin expression in the endometrium of women with recurrent implantation failure and recurrent miscarriage. Fertil. Steril. 2017, 107, 136–143. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, M.M.; Mani, S.K.; Julian, J.; Carson, D.D. Reduction of mucin-1 expression during the receptive phase in the rat uterus. Biol. Reprod. 1998, 58, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.A.; Bazer, F.W.; Burghardt, R.C. Spatial and temporal analyses of integrin and Muc-1 expression in porcine uterine epithelium and trophectoderm in vivo. Biol. Reprod. 1996, 55, 1098–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Fukuda, K.; Ishida, H.; Miyoshi, S.; Koura, T.; Kodama, H.; Nakazawa, H.K.; Ogawa, S. Leukemia inhibitory factor, a potent cardiac hypertrophic cytokine, enhances L-type Ca2+ current and [Ca2+] i transient in cardiomyocytes. J. Mol. Cell. Cardiol. 1999, 31, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Guang, W.; Kim, K.C.; Lillehoj, E.P. MUC1 mucin interacts with calcium-modulating cyclophilin ligand. Int. J. Biochem. Cell Biol. 2009, 41, 1354–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Padh, H.; Nivsarkar, M. Role of the calcium channel in blastocyst implantation: A novel contraceptive target. J. Basic Clin. Physiol. Pharmacol. 2009, 20, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar] [CrossRef] [PubMed]
- Santella, L.; Lim, D.; Moccia, F. Calcium and fertilization: The beginning of life. Trends Biochem. Sci. 2004, 29, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-B.; Brown, E.M.; Hediger, M.A. Apical entry channels in calcium-transporting epithelia. Physiology 2003, 18, 158–163. [Google Scholar] [CrossRef]
- Van Abel, M.; Hoenderop, J.G.; Bindels, R.J. The epithelial calcium channels TRPV5 and TRPV6: Regulation and implications for disease. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2005, 371, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-A.; Yang, H.; Hwang, I.; Jung, E.-M.; Choi, K.-C.; Jeung, E.-B. Expression patterns and potential action of the calcium transport genes TRPV5, TRPV6, NCX1 and PMCA1B in the canine duodenum, kidney and uterus. In Vivo 2011, 25, 773–780. [Google Scholar] [PubMed]
- Lee, B.-M.; Lee, G.-S.; Jung, E.-M.; Choi, K.-C.; Jeung, E.-B. Uterine and placental expression of TRPV6 gene is regulated via progesterone receptor-or estrogen receptor-mediated pathways during pregnancy in rodents. Reprod. Biol. Endocrinol. 2009, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Sprekeler, N.; Kowalewski, M.P.; Boos, A. TRPV6 and Calbindin-D9k-expression and localization in the bovine uterus and placenta during pregnancy. Reprod. Biol. Endocrinol. 2012, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Choi, K.C.; Hyun, S.H.; Jeung, E.B. Coexpression and estrogen-mediated regulation of TRPV6 and PMCA1 in the human endometrium during the menstrual cycle. Mol. Reprod. Dev. 2011, 78, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Chappell, V.; Fenton, S.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R. EDC-2: The endocrine society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, T.J.; Carlson, A.; Schwartz, J.M.; Giudice, L.C. Proceedings of the summit on environmental challenges to reproductive health and fertility: Executive summary. Fertil. Steril. 2008, 89, e1–e20. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Bai, M.-Z.; Huang, X.-F.; Zhang, Y.; Liu, J.; Hu, M.-H.; Zheng, W.-Q.; Jin, F. Preimplantation exposure to bisphenol A and triclosan may lead to implantation failure in humans. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.G.; Shaw, J. Impact of acute bisphenol-A exposure upon intrauterine implantation of fertilized ova and urinary levels of progesterone and 17β-estradiol. Reprod. Toxicol. 2008, 26, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Harazono, A.; Ema, M. Effects of 4-tert-octylphenol on initiation and maintenance of pregnancy following oral administration during early pregnancy in rats. Toxicol. Lett. 2001, 119, 79–84. [Google Scholar] [CrossRef]
- Kim, S.; An, B.-S.; Yang, H.; Jeung, E.-B. Effects of octylphenol and bisphenol A on the expression of calcium transport genes in the mouse duodenum and kidney during pregnancy. Toxicology 2013, 303, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Ahn, C.; Kang, H.Y.; Hong, E.-J.; Hyun, S.-H.; Choi, K.-C.; Jeung, E.-B. Effects of octylphenol and bisphenol A on the metal cation transporter channels of mouse placentas. Int. J. Environ. Res. Public Health 2016, 13, 965. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Davila, J.; Bagchi, M.K.; Bagchi, I.C. Chronic exposure to bisphenol a impairs progesterone receptor-mediated signaling in the uterus during early pregnancy. Recept. Clin. Investig. 2016, 3, e1369. [Google Scholar]
- Varayoud, J.; Ramos, J.G.; Bosquiazzo, V.L.; Lower, M.; Muñoz-de-Toro, M.; Luque, E.H. Neonatal exposure to bisphenol A alters rat uterine implantation-associated gene expression and reduces the number of implantation sites. Endocrinology 2011, 152, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Sugihara, A.; Uchida, K.; Sato, T.; Ohta, Y.; Katsu, Y.; Watanabe, H.; Iguchi, T. Developmental effects of perinatal exposure to bisphenol-A and diethylstilbestrol on reproductive organs in female mice. Reprod. Toxicol. 2002, 16, 107–116. [Google Scholar] [CrossRef]
- Berger, R.G.; Foster, W.G. Bisphenol-A exposure during the period of blastocyst implantation alters uterine morphology and perturbs measures of estrogen and progesterone receptor expression in mice. Reprod. Toxicol. 2010, 30, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Diao, H.; Smith, M.A.; Song, X.; Ye, X. Preimplantation exposure to bisphenol A (BPA) affects embryo transport, preimplantation embryo development, and uterine receptivity in mice. Reprod. Toxicol. 2011, 32, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Yadav, A.; Pakrasi, P. Expression of ER-α and ER-β during peri-implantation period in uterus is essential for implantation and decidualization in golden hamster. Life Sci. 2017, 170, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hild-Petito, S.; Fazleabas, A.T.; Julian, J.; Carson, D.D. Mucin (Muc-1) expression is differentially regulated in uterine luminal and glandular epithelia of the baboon (Papio anubis). Biol. Reprod. 1996, 54, 939–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, W.; Lee, C.; Yeung, W.; Giesy, J.P.; Wong, M.; Zhang, X.; Hecker, M.; Wong, C.K. Effect of perinatal and postnatal bisphenol A exposure to the regulatory circuits at the hypothalamus–pituitary–gonadal axis of CD-1 mice. Reprod. Toxicol. 2011, 31, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, J.; Thygesen, P.S.; Kaerlev, L.; Knudsen, L.E.; Bonde, J.P. Potential exposure to endocrine disrupting chemicals and selected adverse pregnancy outcomes: A follow-up study of pregnant women referred for occupational counselling. J. Occup. Med. Toxicol. 2017, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.G.; Hancock, T. Influence of oral and subcutaneous bisphenol-A on intrauterine implantation of fertilized ova in inseminated female mice. Reprod. Toxicol. 2007, 23, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Borman, E.D.; Foster, W.G.; Greenacre, M.K.; Muir, C.C. Stress lowers the threshold dose at which bisphenol A disrupts blastocyst implantation, in conjunction with decreased uterine closure and e-cadherin. Chem.-Biol. Interact. 2015, 237, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Sugiura-Ogasawara, M.; Ozaki, Y.; Sonta, S.-I.; Makino, T.; Suzumori, K. Exposure to bisphenol A is associated with recurrent miscarriage. Hum. Reprod. 2005, 20, 2325–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.; van der Kemp, A.W.; Hartog, A.; van de Graaf, S.F.; van Os, C.H.; Willems, P.H.; Bindels, R.J. Molecular identification of the apical Ca2+ channel in 1, 25-dihydroxyvitamin D3-responsive epithelia. J. Biol. Chem. 1999, 274, 8375–8378. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-B.; Chen, X.-Z.; Berger, U.V.; Vassilev, P.M.; Tsukaguchi, H.; Brown, E.M.; Hediger, M.A. Molecular cloning and characterization of a channel-like transporter mediating intestinal calcium absorption. J. Biol. Chem. 1999, 274, 22739–22746. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, D.; Tambutté, E.; Kulhanek, E.; Puverel, S.; Scimeca, J.-C.; Allemand, D.; Tambutté, S. Molecular cloning and localization of a PMCA P-type calcium ATPase from the coral Stylophora pistillata. Biochim. Biophys. Acta 2004, 1663, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Linck, B.; Qiu, Z.; He, Z.; Tong, Q.; Hilgemann, D.W.; Philipson, K.D. Functional comparison of the three isoforms of the Na+/Ca2+ exchanger (NCX1, NCX2, NCX3). Am. J. Physiol.-Cell Physiol. 1998, 274, C415–C423. [Google Scholar] [CrossRef]
- Żylińska, L.; Kawecka, I.; Lachowicz, L.; Szemraj, J. The isoform-and location-dependence of the functioning of the plasma membrane calcium pump. Cell. Mol. Biol. Lett. 2002, 7, 1037–1045. [Google Scholar] [PubMed]
- Blaustein, M.P. Sodium/calcium exchange. In Handbook of Cell Signaling, 2nd ed.; Elsevier: New York, NY, USA, 2009; pp. 949–953. [Google Scholar]
- Choi, K.; An, B.; Yang, H.; Jeung, E. Regulation and molecular mechanisms of calcium transport genes: Do they play a role in calcium transport in the uterine endometrium. J. Physiol. Pharmacol. 2011, 62, 499–504. [Google Scholar] [PubMed]
- Belkacemi, L.; Bédard, I.; Simoneau, L.; Lafond, J. Calcium channels, transporters and exchangers in placenta: A review. Cell Calcium 2005, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Orrenius, S.; Nicotera, P. The calcium ion and cell death. J. Neural Transm. Suppl. 1994, 43, 1–11. [Google Scholar] [PubMed]
- Nie, M.; Bal, M.S.; Yang, Z.; Liu, J.; Rivera, C.; Wenzel, A.; Beck, B.B.; Sakhaee, K.; Marciano, D.K.; Wolf, M.T. Mucin-1 increases renal TRPV5 activity in vitro, and urinary level associates with calcium nephrolithiasis in patients. J. Am. Soc. Nephrol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Satokata, I.; Benson, G.; Maas, R. Sexually dimorphic sterility phenotypes in HoxalO-deficient mice. Nature 1995, 374, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.L.; Kaspar, P.; Brunet, L.J.; Bhatt, H.; Gadi, I.; Köntgen, F.; Abbondanzo, S.J. Blastocyst implantation depends on maternal expression of leukaemia inhibitory factor. Nature 1992, 359, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Margioula-Siarkou, C.; Prapas, Y.; Petousis, S.; Milias, S.; Ravanos, K.; Kalogiannidis, I.; Mavromatidis, G.; Haitoglou, C.; Prapas, N.; Rousso, D. LIF and LIF-R expression in the endometrium of fertile and infertile women: A prospective observational case-control study. Mol. Med. Rep. 2016, 13, 4721–4728. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef] [PubMed]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.A.; Zhang, Y.; Ing, N.H. ICI 182,780 acts as a partial agonist and antagonist of estradiol effects in specific cells of the sheep uterus. J. Steroid Biochem. Mol. Biol. 2001, 77, 281–287. [Google Scholar] [CrossRef]
- Movérare-Skrtic, S.; Börjesson, A.E.; Farman, H.H.; Sjögren, K.; Windahl, S.H.; Lagerquist, M.K.; Andersson, A.; Stubelius, A.; Carlsten, H.; Gustafsson, J.-Å. The estrogen receptor antagonist ICI 182,780 can act both as an agonist and an inverse agonist when estrogen receptor α AF-2 is modified. Proc. Natl. Acad. Sci. USA 2014, 111, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; He, Y.; Shang, D.; Pan, J.; Wang, H.; Chen, H.; Zhu, Z.; Wan, L.; Wang, X. Estrogen and pure antiestrogen fulvestrant (ICI 182 780) augment cell–matrigel adhesion of MCF-7 breast cancer cells through a novel G protein coupled estrogen receptor (GPR30)-to-calpain signaling axis. Toxicol. Appl. Pharmacol. 2014, 275, 176–181. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, D.N.; Jung, E.-M.; Ahn, C.; Lee, J.-H.; Yoo, Y.-M.; Jeung, E.-B. Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse. Int. J. Environ. Res. Public Health 2018, 15, 1614. https://doi.org/10.3390/ijerph15081614
Tran DN, Jung E-M, Ahn C, Lee J-H, Yoo Y-M, Jeung E-B. Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse. International Journal of Environmental Research and Public Health. 2018; 15(8):1614. https://doi.org/10.3390/ijerph15081614
Chicago/Turabian StyleTran, Dinh Nam, Eui-Man Jung, Changhwan Ahn, Jae-Hwan Lee, Yeong-Min Yoo, and Eui-Bae Jeung. 2018. "Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse" International Journal of Environmental Research and Public Health 15, no. 8: 1614. https://doi.org/10.3390/ijerph15081614
APA StyleTran, D. N., Jung, E. -M., Ahn, C., Lee, J. -H., Yoo, Y. -M., & Jeung, E. -B. (2018). Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse. International Journal of Environmental Research and Public Health, 15(8), 1614. https://doi.org/10.3390/ijerph15081614