The Effects of Vitamin E from Elaeis guineensis (Oil Palm) in a Rat Model of Bone Loss Due to Metabolic Syndrome

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Metabolic Syndrome Parameters
2.3. Dual-Energy X-ray Absorptiometry (DXA)
2.4. Bone Processing and Histomorphometry
2.5. Bone Remodeling Markers and Hormones
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [PubMed]
- Lee, K. Metabolic Syndrome and Osteoporosis in Relation to Muscle Mass. Calcif. Tissue Int. 2015, 97, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.K.; Choi, H.J. The relationship between low bone mass and metabolic syndrome in Korean women. Osteoporos. Int. 2010, 21, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Choe, J.W.; Kim, H.K.; Bae, S.J.; Kim, B.J.; Lee, S.H.; Koh, J.M.; Han, K.O.; Park, H.M.; Kim, G.S. Negative association between metabolic syndrome and bone mineral density in Koreans, especially in men. Calcif. Tissue Int. 2010, 86, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Jamil, N.A.; Ima-Nirwana, S. Osteoporosis is associated with metabolic syndrome induced by high-carbohydrate high-fat diet in a rat model. Biomed. Pharmacother. 2017, 98, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Effects of metabolic syndrome on bone mineral density, histomorphometry and remodelling markers in male rats. PLoS ONE 2018, 13, e0192416. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Shafie, S.R.; Prasadam, I.; Crawford, R.; Panchal, S.K.; Brown, L.; Xiao, Y. Saturated fatty acids induce development of both metabolic syndrome and osteoarthritis in rats. Sci. Rep. 2017, 7, 46457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, A.R.; Panchal, S.K.; Friis, T.; Sekar, S.; Crawford, R.; Brown, L.; Xiao, Y.; Prasadam, I. Obesity-associated metabolic syndrome spontaneously induces infiltration of pro-inflammatory macrophage in synovium and promotes osteoarthritis. PLoS ONE 2017, 12, e0183693. [Google Scholar] [CrossRef] [PubMed]
- Hukshorn, C.J.; Saris, W.H. Leptin and energy expenditure. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Takahashi, N.; Hileman, S.M.; Patel, H.R.; Berg, A.H.; Pajvani, U.B.; Scherer, P.E.; Ahima, R.S. Adiponectin acts in the brain to decrease body weight. Nat. Med. 2004, 10, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, I.; Fujino, T.; Douchi, T. The leptin receptor in human osteoblasts and the direct effect of leptin on bone metabolism. Gynecol. Endocrinol. 2004, 19, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Chen, C.Y.; Chuang, T.Y.; Lin, Y.; Liu, H.Y.; Mersmann, H.J.; Wu, S.C.; Ding, S.T. Adiponectin receptor 1 regulates bone formation and osteoblast differentiation by GSK-3beta/beta-catenin signaling in mice. Bone 2014, 64, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Vianna, A.G.D.; Sanches, C.P.; Barreto, F.C. Review article: Effects of type 2 diabetes therapies on bone metabolism. Diabetol. Metab. Syndr. 2017, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, J.; Farr, O.M.; Mantzoros, C.S. The role of leptin in regulating bone metabolism. Metabolism 2015, 64, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducy, P.; Amling, M.; Takeda, S.; Priemel, M.; Schilling, A.F.; Beil, F.T.; Shen, J.; Vinson, C.; Rueger, J.M.; Karsenty, G. Leptin inhibits bone formation through a hypothalamic relay: A central control of bone mass. Cell 2000, 100, 197–207. [Google Scholar] [CrossRef]
- Iwase, S.; Nishimura, N.; Mano, T. Osteoporosis in Spaceflight. In Topics in Osteoporosis; Flores, M.V., Ed.; InTech: Rijeka, Croatia, 2013; Chapter 12. [Google Scholar] [Green Version]
- Lubkowska, A.; Dobek, A.; Mieszkowski, J.; Garczynski, W.; Chlubek, D. Adiponectin as a Biomarker of Osteoporosis in Postmenopausal Women: Controversies. Dis. Mark. 2014, 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Ferron, M. The contribution of bone to whole-organism physiology. Nature 2012, 481, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J. A Comprehensive Review on Metabolic Syndrome. Cardiol. Res. Pract. 2014, 2014, 21. [Google Scholar] [CrossRef] [PubMed]
- Kling, J.M.; Clarke, B.L.; Sandhu, N.P. Osteoporosis Prevention, Screening, and Treatment: A Review. J. Women’s Health 2014, 23, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.Y.; Poudyal, H.; Ward, L.C.; Brown, L. Tocotrienols reverse cardiovascular, metabolic and liver changes in high carbohydrate, high fat diet-fed rats. Nutrients 2012, 4, 1527–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory gamma- and delta-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2015, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Kang, I.; Fang, X.; Wang, W.; Lee, M.A.; Hollins, R.R.; Marshall, M.R.; Chung, S. Gamma-tocotrienol attenuates high-fat diet-induced obesity and insulin resistance by inhibiting adipose inflammation and M1 macrophage recruitment. Int. J. Obes. 2015, 39, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Rashid Khan, M.; Siddiqui, W.A. Comparative hypoglycemic and nephroprotective effects of tocotrienol rich fraction (TRF) from palm oil and rice bran oil against hyperglycemia induced nephropathy in type 1 diabetic rats. Chem. Biol. Interact. 2010, 188, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Newaz, M.A.; Yousefipour, Z.; Nawal, N.; Adeeb, N. Nitric oxide synthase activity in blood vessels of spontaneously hypertensive rats: Antioxidant protection by gamma-tocotrienol. J. Physiol. Pharmacol. 2003, 54, 319–327. [Google Scholar] [PubMed]
- Qureshi, A.A.; Peterson, D.M.; Hasler-Rapacz, J.O.; Rapacz, J. Novel tocotrienols of rice bran suppress cholesterogenesis in hereditary hypercholesterolemic swine. J. Nutr. 2001, 131, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Chopra, K. Attenuation of diabetic nephropathy by tocotrienol: Involvement of NFkB signaling pathway. Life Sci. 2009, 84, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Bishnoi, M.; Tiwari, V.; Chopra, K. Suppression of NF-kappabeta signaling pathway by tocotrienol can prevent diabetes associated cognitive deficits. Pharmacol. Biochem. Behav. 2009, 92, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Soelaiman, I.N.; Ming, W.; Abu Bakar, R.; Hashnan, N.A.; Mohd Ali, H.; Mohamed, N.; Muhammad, N.; Shuid, A.N. Palm tocotrienol supplementation enhanced bone formation in oestrogen-deficient rats. Int. J. Endocrinol. 2012, 2012, 532862. [Google Scholar] [CrossRef] [PubMed]
- Ima-Nirwana, S.; Kiftiah, A.; Zainal, A.; Norazlina, M.; Gapor, M.; Khalid, B. Palm vitamin E prevents osteoporosis in orchidectomized growing male rats. Nat. Prod. Sci. 2000, 6, 155–160. [Google Scholar]
- Ahmad, N.S.; Khalid, B.A.; Luke, D.A.; Ima Nirwana, S. Tocotrienol offers better protection than tocopherol from free radical-induced damage of rat bone. Clin. Exp. Pharmacol. Physiol. 2005, 32, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Hermizi, H.; Faizah, O.; Ima-Nirwana, S.; Ahmad Nazrun, S.; Norazlina, M. Beneficial effects of tocotrienol and tocopherol on bone histomorphometric parameters in sprague-dawley male rats after nicotine cessation. Calcif. Tissue Int. 2009, 84, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The effects of palm tocotrienol on metabolic syndrome and bone loss in male rats induced by high-carbohydrate high-fat diet. J. Funct. Foods. 2018, 44, 246–254. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Exploring the potential of tocotrienol from Bixa. orellana. as a single agent targeting metabolic syndrome and bone loss. Bone 2018, 116, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The effects of a modified high-carbohydrate high-fat diet on metabolic syndrome parameters in male rats. Exp. Clin. Endocrinol. Diabetes 2018, 126, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.; Brown, L. Comparison of purple carrot juice and beta-carotene in a high-carbohydrate, high-fat diet-fed rat model of the metabolic syndrome. Br. J. Nutr. 2010, 104, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Lu, X.; Sun, M.; Li, K.; Shen, L.; Wu, T. Protective effects of L-arabinose in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. Food Nutr. Res. 2015, 59, 28886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Vitamin E as a Potential Interventional Treatment for Metabolic Syndrome: Evidence from Animal and Human Studies. Front. Pharmacol. 2017, 8, 444. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The Relationship between Metabolic Syndrome and Osteoporosis: A Review. Nutrients 2016, 8, 347. [Google Scholar] [CrossRef] [PubMed]
- Norazlina, M.; Ima-Nirwana, S.; Gapor, M.T.; Khalid, B.A. Palm vitamin E is comparable to alpha-tocopherol in maintaining bone mineral density in ovariectomised female rats. Exp. Clin. Endocrinol. Diabetes 2000, 108, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, H.M.; New, S.A.; Golden, M.H.; Campbell, M.K.; Reid, D.M. Nutritional associations with bone loss during the menopausal transition: Evidence of a beneficial effect of calcium, alcohol, and fruit and vegetable nutrients and of a detrimental effect of fatty acids. Am. J. Clin. Nutr. 2004, 79, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Maggio, D.; Barabani, M.; Pierandrei, M.; Polidori, M.C.; Catani, M.; Mecocci, P.; Senin, U.; Pacifici, R.; Cherubini, A. Marked decrease in plasma antioxidants in aged osteoporotic women: Results of a cross-sectional study. J. Clin. Endocrinol. Metab. 2003, 88, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.L.; Cauley, J.A.; Pettinger, M.; Jackson, R.; Lacroix, A.; Leboff, M.S.; Lewis, C.E.; Nevitt, M.C.; Simon, J.A.; Stone, K.L.; et al. Lack of a relation between vitamin and mineral antioxidants and bone mineral density: Results from the Women’s Health Initiative. Am. J. Clin. Nutr. 2005, 82, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Ima-Nirwana, S. The effects of alpha-tocopherol on bone: A double-edged sword? Nutrients 2014, 6, 1424–1441. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Martinez, G.; Soriguer, F.J.; Gonzalez-Romero, S.; Tinahones, F.; Moreno, F.; de Adana, S.R.; Garriga, M.J.; Esteva, I.; Garcia-Arnes, J.; Gomez-Zumaquero, J.M.; et al. Serum leptin and habitual fatty acid dietary intake in patients with type 1 diabetes mellitus. Eur. J. Endocrinol. 2000, 142, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handjieva-Darlenska, T.; Boyadjieva, N. The effect of high-fat diet on plasma ghrelin and leptin levels in rats. J. Physiol. Biochem. 2009, 65, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Ludgero-Correia, A.; Aguila, M.B.; Mandarim-de-Lacerda, C.A.; Faria, T.S. Effects of high-fat diet on plasma lipids, adiposity, and inflammatory markers in ovariectomized C57BL/6 mice. Nutrition 2012, 28, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Chia, L.L.; Jantan, I.; Chua, K.H.; Lam, K.W.; Rullah, K.; Aluwi, M.F.M. Effects of Tocotrienols on Insulin Secretion-Associated Genes Expression of Rat Pancreatic Islets in a Dynamic Culture. Front. Pharmacol. 2016, 7, 291. [Google Scholar] [CrossRef] [PubMed]
- Elefteriou, F.; Takeda, S.; Ebihara, K.; Magre, J.; Patano, N.; Ae Kim, C.; Ogawa, Y.; Liu, X.; Ware, S.M.; Craigen, W.J.; et al. Serum leptin level is a regulator of bone mass. Proc. Natl. Acad. Sci. USA 2004, 101, 3258–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, R.T.; Kalra, S.P.; Wong, C.P.; Philbrick, K.A.; Lindenmaier, L.B.; Boghossian, S.; Iwaniec, U.T. Peripheral leptin regulates bone formation. J. Bone Miner. Res. 2013, 28, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Furukawa, F.; Nishikawa, A.; Miyauchi, M.; Son, H.Y.; Imazawa, T.; Hirose, M. Oral toxicity of a tocotrienol preparation in rats. Food Chem. Toxicol. 2001, 39, 799–805. [Google Scholar] [CrossRef]


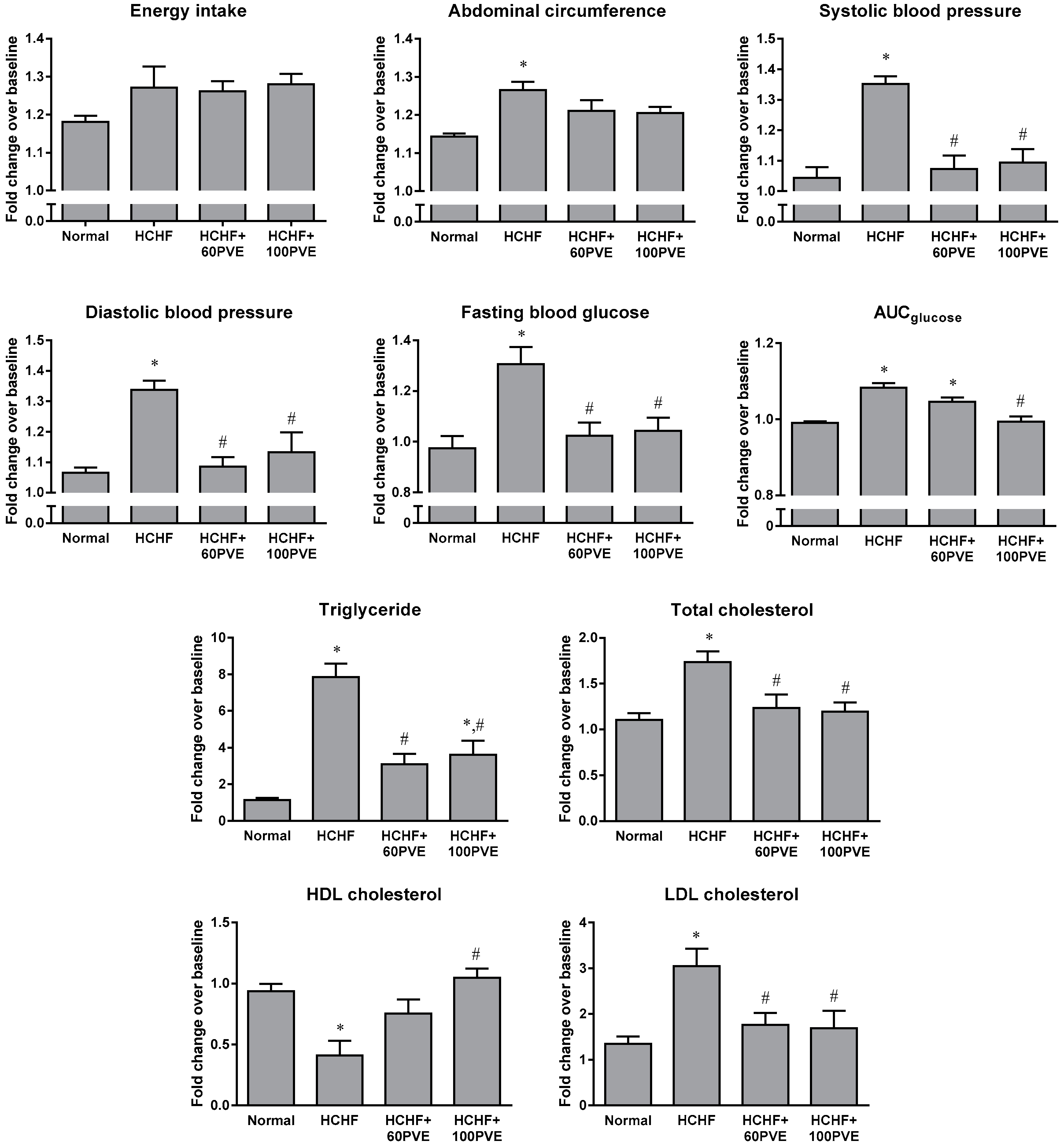{kind=link}
| Baseline | Normal | HCHF | HCHF + 60 PVE | HCHF + 100 PVE | |
|---|---|---|---|---|---|
| Body weight (g) | 225.50 ± 6.22 | 374.67 ± 3.61 a | 372.17 ± 10.84 a | 361.67 ± 13.11 a | 348.67 ± 11.36 a |
| Fat mass (g) | 11.83 ± 2.32 | 26.35 ± 4.11 | 58.57 ± 5.00 a,* | 56.45 ± 3.53 a,* | 61.42 ± 2.91 a,* |
| Liver weight (g) | 9.88 ± 0.36 | 11.81 ± 0.18 | 14.81 ± 0.73 a,* | 12.85 ± 0.51 a | 12.78 ± 0.86 a |
| Baseline | Normal | HCHF | HCHF + 60 PVE | HCHF + 100 PVE | |
|---|---|---|---|---|---|
| Whole body BMD (g/cm2) | 0.129 ± 0.001 | 0.182 ± 0.003 a | 0.182 ± 0.005 a | 0.186 ± 0.005 a | 0.186 ± 0.002 a |
| Left femur BMD (g/cm2) | 0.228 ± 0.002 | 0.304 ± 0.006 a | 0.306 ± 0.007 a | 0.313 ± 0.007 a | 0.299 ± 0.007 a |
| Right femur BMD (g/cm2) | 0.230 ± 0.005 | 0.283 ± 0.007 a | 0.287 ± 0.012 a | 0.295 ± 0.010 a | 0.302 ± 0.008 a |
| Left tibia BMD (g/cm2) | 0.159 ± 0.003 | 0.207 ± 0.003 a | 0.215 ± 0.005 a | 0.219 ± 0.002 a | 0.202 ± 0.009 a |
| Right tibia BMD (g/cm2) | 0.158 ± 0.002 | 0.185 ± 0.009 | 0.190 ± 0.013 | 0.197 ± 0.010 | 0.197 ± 0.012 |
| Baseline | Normal | HCHF | HCHF + 60 PVE | HCHF + 100 PVE | |
|---|---|---|---|---|---|
| Ob.S/BS (%) | 5.22 ± 0.77 | 8.67 ± 0.58 a | 4.28 ± 0.30 * | 8.58 ± 0.88 a,# | 8.38 ± 0.92 a,# |
| Oc.S/BS (%) | 4.08 ± 0.64 | 5.74 ± 0.93 | 4.71 ± 0.53 | 5.25 ± 0.22 | 4.22 ± 0.35 |
| ES/BS (%) | 7.47 ± 0.62 | 6.07 ± 0.73 | 12.04 ± 1.37 a,* | 9.49 ± 0.68* | 8.24 ± 0.31# |
| OS/BS (%) | 5.76 ± 1.01 | 9.75 ± 0.72 a | 4.72 ± 0.47* | 8.26 ± 0.32# | 8.46 ± 0.16 a,# |
| OV/BV (%) | 4.70 ± 0.36 | 7.88 ± 0.86 a | 3.47 ± 0.63* | 4.60 ± 0.31* | 4.54 ± 0.69* |
| sLS/BS (%) | 11.34 ± 1.01 | 11.76 ± 0.79 | 13.76 ± 0.77 | 7.82 ± 1.05*,# | 9.98 ± 1.00 |
| dLS/BS (%) | 10.65 ± 0.83 | 10.44 ± 0.86 | 9.13 ± 0.53 | 11.51 ± 0.98 | 8.20 ± 1.23 |
| MS/BS (%) | 17.62 ± 0.89 | 17.11 ± 0.71 | 16.73 ± 0.69 | 14.19 ± 0.61 a | 15.21 ± 0.64 |
| MAR (µm/day) | 1.60 ± 0.07 | 1.65 ± 0.19 | 1.49 ± 0.08 | 1.78 ± 0.14 | 1.42 ± 0.13 |
| BFR (µm3/µm2/day) | 25.45 ± 1.77 | 23.92 ± 1.81 | 21.37 ± 1.41 | 23.53 ± 1.63 | 21.52 ± 2.24 |
| BV/TV (%) | 32.09 ± 2.56 | 29.78 ± 2.30 | 18.98 ± 0.90 a,* | 32.55 ± 1.36# | 30.90 ± 0.63 # |
| Tb.Th (µm) | 108.20 ± 6.95 | 92.68 ± 4.86 | 70.28 ± 13.00 a | 101.44 ± 9.46 | 99.44 ± 6.72 |
| Tb.N (µm−1) | 0.0030 ± 0.0003 | 0.0032 ± 0.0002 | 0.0033 ± 0.0007 | 0.0034 ± 0.0001 | 0.0032 ± 0.0002 |
| Tb.Sp (µm) | 250.47 ± 29.35 | 226.24 ± 17.24 | 313.71 ± 60.97 | 207.64 ± 8.73 | 222.95 ± 13.43 |
| Baseline | Normal | HCHF | HCHF + 60 PVE | HCHF + 100 PVE | |
|---|---|---|---|---|---|
| Osteocalcin (ng/mL) | 773.28 ± 38.01 | 401.94 ± 13.34 a | 305.55 ± 15.65 a | 386.49 ± 25.73 a | 369.19 ± 22.15 a |
| CTX-1 (ng/mL) | 42.47 ± 3.13 | 17.87 ± 0.36 a | 24.48 ± 1.44 a,* | 19.22 ± 0.69 a | 20.85 ± 1.22 a |
| Baseline | Normal | HCHF | HCHF + 60 PVE | HCHF + 100 PVE | |
|---|---|---|---|---|---|
| Leptin (pg/mL) | 530.36 ± 74.14 | 1134.70 ± 153.36 | 3425.55 ± 377.31 a,* | 2050.00 ± 246.92 a,*,# | 530.36 ± 74.14 |
| Adiponectin (ng/mL) | 59.11 ± 1.29 | 58.51 ± 1.76 | 50.87 ± 1.43* | 56.55 ± 2.58 | 59.11 ± 1.29 |
| Insulin (ng/mL) | 0.35 ± 0.04 | 0.38 ± 0.02 | 6.51 ± 0.55 a,* | 7.62 ± 0.50 a,* | 0.35 ± 0.04 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The Effects of Vitamin E from Elaeis guineensis (Oil Palm) in a Rat Model of Bone Loss Due to Metabolic Syndrome. Int. J. Environ. Res. Public Health 2018, 15, 1828. https://doi.org/10.3390/ijerph15091828
Wong SK, Chin K-Y, Suhaimi FH, Ahmad F, Ima-Nirwana S. The Effects of Vitamin E from Elaeis guineensis (Oil Palm) in a Rat Model of Bone Loss Due to Metabolic Syndrome. International Journal of Environmental Research and Public Health. 2018; 15(9):1828. https://doi.org/10.3390/ijerph15091828
Chicago/Turabian StyleWong, Sok Kuan, Kok-Yong Chin, Farihah Hj Suhaimi, Fairus Ahmad, and Soelaiman Ima-Nirwana. 2018. "The Effects of Vitamin E from Elaeis guineensis (Oil Palm) in a Rat Model of Bone Loss Due to Metabolic Syndrome" International Journal of Environmental Research and Public Health 15, no. 9: 1828. https://doi.org/10.3390/ijerph15091828
APA StyleWong, S. K., Chin, K. -Y., Suhaimi, F. H., Ahmad, F., & Ima-Nirwana, S. (2018). The Effects of Vitamin E from Elaeis guineensis (Oil Palm) in a Rat Model of Bone Loss Due to Metabolic Syndrome. International Journal of Environmental Research and Public Health, 15(9), 1828. https://doi.org/10.3390/ijerph15091828