Effects of Intermittent Hypoxia on Pulmonary Vascular and Systemic Diseases



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
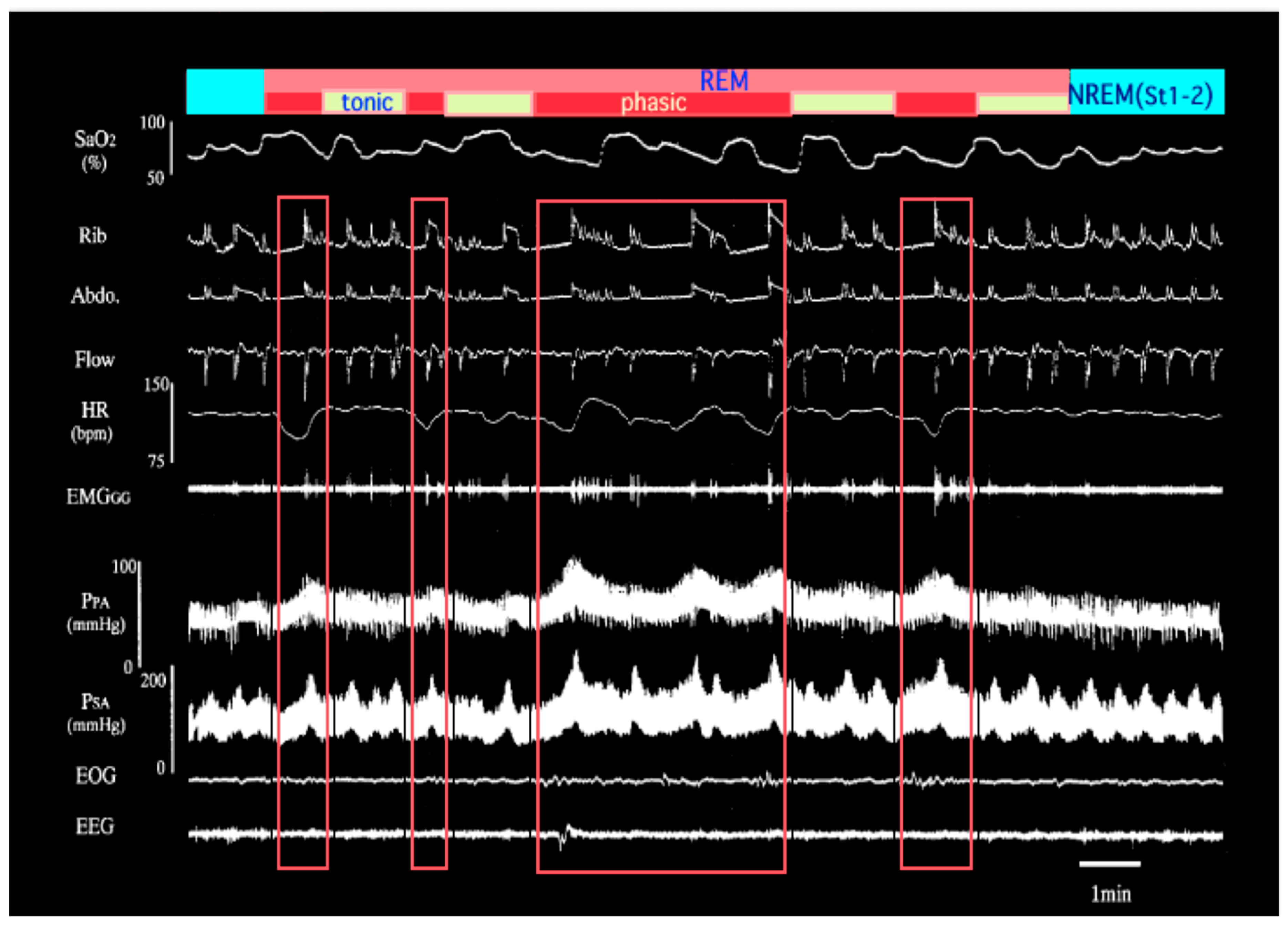
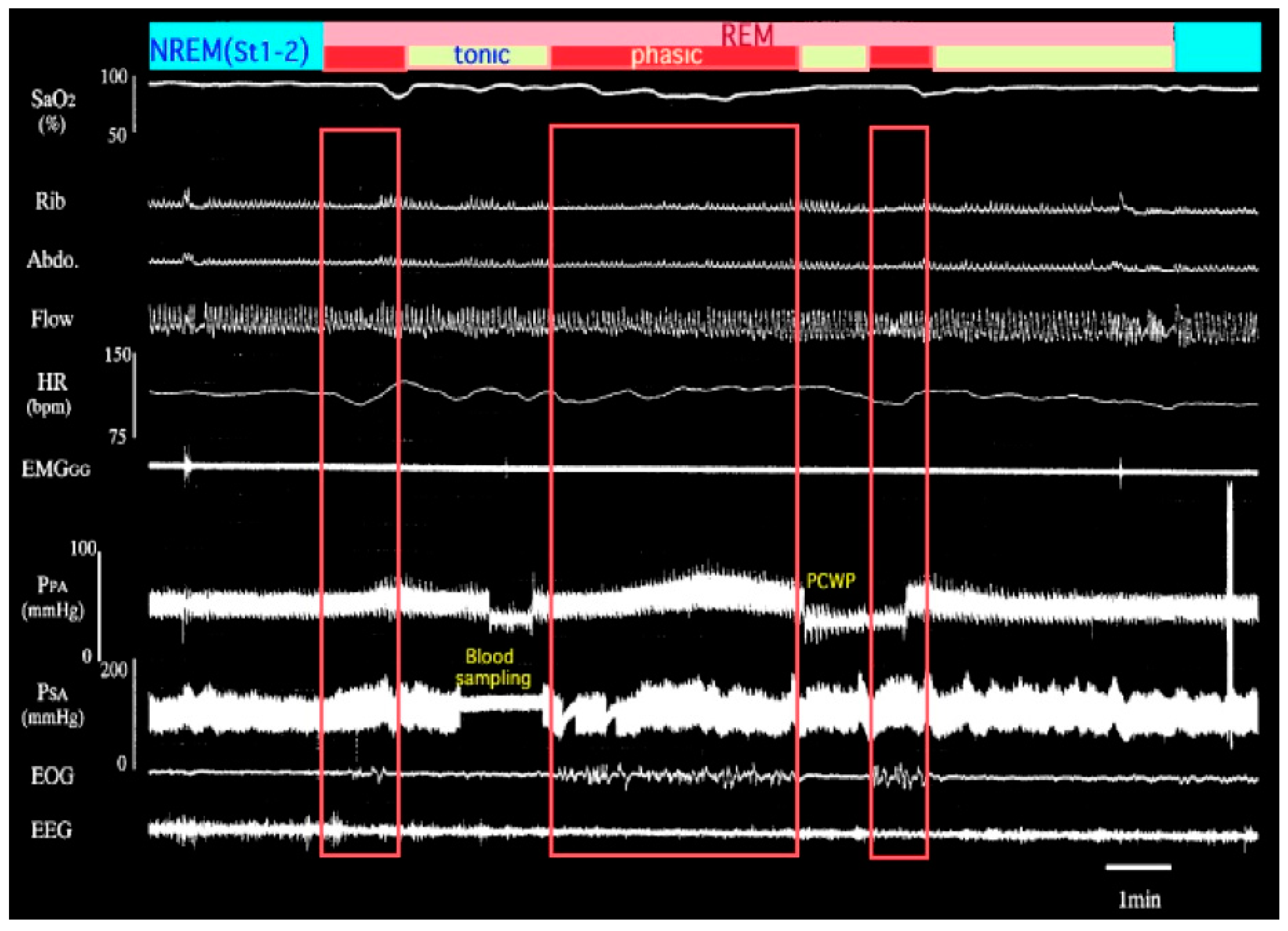
2. Effects of Intermittent Hypoxia on Pulmonary Vascular Diseases
3. Effects of Intermittent Hypoxia on Systemic Diseases
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANGPTL | Angiopoietin-like protein |
| BMI | Body mass index |
| CART | Cocaine- and amphetamine-regulated transcript |
| CB | Carotid body |
| CCL2 | C-C motif chemokine ligand 2 |
| CNS | Central nervous system |
| CPAP | Continuous positive airway pressure |
| ENS | Enteric nervous system |
| GI | Gastrointestinal |
| GIS | Glucose-induced insulin secretion |
| HPV | Hypoxic pulmonary vasoconstriction |
| IH | Intermittent hypoxia |
| IMT | Intima-media thickness |
| IL-6 | Interleukin-6 |
| Mφ | Μacrophages |
| NREM | Non-REM |
| OHS | Obesity hypoventilation syndrome |
| OSA | Obstructive sleep apnea |
| AP | Pulmonary arterial pressure |
| PH | Pulmonary hypertension |
| POMC | Proopiomelanocortin |
| REM | Rapid eye movement |
| SNP | Single-nucleotide polymorphism |
| SDB | Sleep disordered breathing |
| SpO2 | Arterial oxygen saturation |
| TNF-α | Tumor necrosis factor-α |
References
- Senaratna, C.V.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Campbell, B.E.; Matheson, M.C.; Hamilton, G.S.; Dharmage, S.C. Prevalence of obstructive sleep apnea in the general population: A systematic review. Sleep Med. Rev. 2017, 34, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Nakano, H.; Maekawa, J.; Okamoto, Y.; Ohnishi, Y.; Suzuki, T.; Kimura, H. Oxidative stress in obstructive sleep apnea. Chest 2005, 127, 1674–1679. [Google Scholar] [CrossRef]
- Yamauchi, M.; Tamaki, S.; Tomoda, K.; Yoshikawa, M.; Fukuoka, A.; Makinodan, K.; Koyama, N.; Suzuki, T.; Kimura, H. Evidence for activation of nuclear factor kappaB in obstructive sleep apnea. Sleep Breath. 2006, 10, 189–193. [Google Scholar] [CrossRef]
- Kimura, H.; Niijima, M.; Edo, H.; Honda, Y.; Kuriyama, T. Differences in the response of genioglossal muscle activity to sustained hypoxia between healthy subjects and patients with obstructive sleep apnea. Respiration 1994, 61, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Kunitomo, F.; Kimura, H.; Tatsumi, K.; Okita, S.; Tojima, H.; Kuriyama, T.; Honda, Y. Abnormal breathing during sleep and chemical control of breathing during wakefulness in patients with sleep apnea syndrome. Am. Rev. Respir. Dis. 1989, 139, 164–169. [Google Scholar] [CrossRef]
- Kimura, H.; Tatsumi, K.; Kunitomo, F.; Okita, S.; Tojima, H.; Kouchiyama, S.; Masuyama, S.; Shinozaki, T.; Honda, Y.; Kuriyama, T. Progesterone therapy for sleep apnea syndrome evaluated by occlusion pressure responses to exogenous loading. Am. Rev. Respir. Dis. 1989, 139, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Borel, J.-C.; Melo-Silva, C.A.; Gakwaya, S.; Rousseau, E.; Series, F. Diaphragm and genioglossus corticomotor excitability in patients with obstructive sleep apnea and control subjects. J. Sleep Res. 2016, 25, 23–60. [Google Scholar] [CrossRef]
- Sivam, S.; Yee, B.; Wong, K.; Wang, D.; Grunstein, R.; Piper, A. Obesity hyperventilation syndrome: Early detection to nocturnal-only hypercapnia in an obese population. J. Clin. Sleep Med. 2018, 14, 1477–1484. [Google Scholar] [CrossRef]
- Held, M.; Walthelm, J.; Baron, S.; Roth, C.; Jany, B. Functional impact of pulmonary hypertension due to hypoventilation and changes under noninvasive ventilation. Eur. Respir. J. 2014, 43, 156–165. [Google Scholar] [CrossRef]
- Mokhlesi, B.; Tulaimat, A. Recent advances in obesity hypoventilation syndrome. Chest 2007, 132, 1322–1336. [Google Scholar] [CrossRef]
- Shirai, M.; Matsukawa, K.; Nishiura, N.; Kawaguchi, A.T.; Ninomiya, I. Changes in efferent pulmonary sympathetic nerve activity during systemic hypoxia in anesthetized cats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1995, 269, R1404–R1409. [Google Scholar] [CrossRef]
- Shirai, M.; Tsuchimochi, H.; Nagai, H.; Gray, E.; Pearson, J.T.; Sonobe, T.; Yoshimoto, M.; Inagaki, T.; Fujii, Y.; Umetani, K.; et al. Pulmonary vascular tone is dependent on the central modulation of sympathetic nerve activity following chronic intermittent hypoxia. Basic Res. Cardiol. 2014, 109, 432. [Google Scholar] [CrossRef]
- Costa-Silva, J.H.; Zoccal, D.B.; Machado, B.H. Chronic intermittent hypoxia alters glutamatergic control of sympathetic and respiratory activities in the commissural NTS of rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R785–R793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niijima, M.; Kimura, H.; Edo, H.; Shinozaki, T.; Kang, J.; Masuyama, S.; Tatsumi, K.; Kuriyama, T. Manifestation of pulmonary hypertension during REM sleep in obstructive sleep apnea syndrome. Am. J. Respir. Crit. Care Med. 1999, 159, 1766–1772. [Google Scholar] [CrossRef]
- Huggett, R.J.; Burns, J.; Mackintosh, A.F.; Mary, D.A.S.G. Sympathetic neural activation in nondiabetic metabolic syndrome and its further augmentation by hypertension. Hypertension 2004, 44, 847–852. [Google Scholar] [CrossRef]
- Chen, D.L.; Brown, R.; Liess, C.; Poljak, A.; Xu, A.; Zhang, J.; Trenell, M.; Jenkins, A.; Chisholm, D.; Samocha-Bonet, D.; et al. Muscle sympathetic nerve activity is associated with liver insulin sensitivity in obese non-diabetic men. Front. Physiol. 2017, 8, 101. [Google Scholar] [CrossRef]
- Battault, S.; Meziat, C.; Nascimento, A.; Braud, L.; Gayrard, S.; Legros, C.; De Nardi, F.; Drai, J.; Cazorla, O.; Thireau, J.; et al. Vascular endothelial function masks increased sympathetic vasopressor activity in rats with metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H497–H507. [Google Scholar] [CrossRef] [PubMed]
- West, S.D.; Nicoll, D.J.; Stradling, J.R. Prevalance of obstructive sleep apnoea in men with type 2 diabetes. Thorax 2006, 61, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Punjabi, N.M.; Shahar, E.; Redline, S.; Gottlieb, D.J.; Givelber, R.; Rresnick, H.E. Sleep-disordered breathing, glucose intolerance, and insulin resistance: The Sleep Heart Health Study. Am. J. Epidemiol. 2004, 160, 521–530. [Google Scholar] [CrossRef]
- Ip, M.S.; Lam, B.; Ng, M.M.; Lam, W.K.; Tsang, K.W.; Lam, K.S. Obstructive sleep apnea is independently associated with insulin resistance. Am. J. Respir. Crit. Care Med. 2002, 165, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Jehan, S.; Myers, A.K.; Zizi, F.; Pandi-Perumal, S.R.; Jean-Louis, G.; McFarlane, S.I. Obesity, obstructive sleep apnea and type 2 diabetes mellitus: Epidemiology and pathophysiologic insights. Sleep Med. Disord. 2018, 2, 52–58. [Google Scholar]
- Seetho, I.W.; Wilding, J.P. Sleep-disordered breathing, type 2 diabetes and the metabolic syndrome. Chron. Respir. Dis. 2014, 11, 257–275. [Google Scholar] [CrossRef]
- Bonsignore, M.R.; Esquinas, C.; Barceló, A.; Sanchez-de-la-Torre, M.; Paternó, A.; Duran-Cantolla, J.; Marín, J.M.; Barbé, F. Metabolic syndrome, insulin resistance and sleepiness in real-life obstructive sleep apnoea. Eur. Respir. J. 2012, 39, 1136–1143. [Google Scholar] [CrossRef]
- Ribeiro, M.J.; Sacramento, J.F.; Gonzalez, C.; Guarino, M.P.; Monteiro, E.C.; Conde, S.V. Carotid body denervation prevents the development of insulin resistance and hypertension induced by hypercaloric diets. Diabetes 2013, 62, 2905–2916. [Google Scholar] [CrossRef]
- Shahar, E.; Whitney, C.W.; Redline, S.; Lee, E.T.; Newman, A.B.; Nieto, F.J.; O’Connor, G.T.; Boland, L.L.; Schwartz, J.E.; Samet, J.M. Sleep-disordered breathing and cardiovascular disease: Cross-sectional results of the Sleep Heart Health Study. Am. J. Respir. Crit. Care Med. 2001, 163, 19–25. [Google Scholar] [CrossRef]
- Muraki, I.; Tanigawa, T.; Yamagishi, K.; Sakurai, S.; Ohira, T.; Imano, H.; Kiyama, M.; Kitamura, A.; Sato, S.; Shimamoto, T.; et al. Nocturnal intermittent hypoxia and metabolic syndrome; the effect of being overweight: The CIRCS study. J. Atheroscler. Thromb. 2010, 17, 369–377. [Google Scholar] [CrossRef]
- Hoyos, C.M.; Melehan, K.L.; Liu, P.Y.; Grunstein, R.R.; Phillips, C.L. Does obstructive sleep apnea cause endothelial dysfunction? A critical review of the literature. Sleep Med. Rev. 2015, 20, 15–26. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Boß, M.; Zezina, E.; Weigert, A.; Dehne, N.; Fleming, I.; Brüne, B.; Namgaladze, D. Hypoxia potentiates palmitate-induced pro-inflammatory activation of primary human macrophages. J. Biol. Chem. 2016, 291, 413–424. [Google Scholar] [CrossRef]
- Akhtar, S.; Hartmann, P.; Karshovska, E.; Rinderknecht, F.-A.; Subramanian, P.; Gremse, F.; Grommes, J.; Jacobs, M.; Kiessling, F.; Weber, C.; et al. Endothelial hypoxia-inducible factor-1α promotes atherosclerosis and monocyte recruitment by upregulating microRNA-19a. Hypertension 2015, 66, 1220–1226. [Google Scholar] [CrossRef]
- Yamauchi, M.; Kimura, H. Oxidative stress in obstructive sleep apnea: Putative pathways to the cardiovascular complications. Antioxid. Redox Signal. 2008, 10, 755–768. [Google Scholar] [CrossRef]
- Suzuki, T.; Nakano, H.; Maekawa, J.; Okamoto, Y.; Ohnishi, Y.; Yamauchi, M.; Kimura, H. Obstructive sleep apnea and carotid-artery intima-media thickness. Sleep 2004, 27, 129–133. [Google Scholar] [CrossRef]
- Tamaki, S.; Yamauchi, M.; Fukuoka, A.; Makinodan, K.; Koyama, N.; Tomoda, K.; Yoshikawa, M.; Kimura, H. Nocturnal hypoxic stress activates invasive ability of monocytes in patients with obstructive sleep apnoea syndrome. Respirology 2009, 14, 689–694. [Google Scholar] [CrossRef]
- Hardy, R.; Wills, A.K.; Wong, A.; Elks, C.E.; Wareham, N.J.; Loos, R.J.F.; Kuh, D.; Ong, K.K. Life course variations in the associations between FTO and MC4R gene variants and body size. Hum. Mol. Genet. 2010, 19, 545–552. [Google Scholar] [CrossRef]
- Muraki, I.; Tanigawa, T.; Yamagishi, K.; Sakurai, S.; Ohira, T.; Imano, H.; Kitamura, A.; Kiyama, M.; Sato, S.; Shimamoto, T.; et al. Nocturnal intermittent hypoxia and the development of type 2 diabetes: The Circulatory Risk in Communities Study (CIRCS). Diabetologia 2010, 53, 481–488. [Google Scholar] [CrossRef]
- Nagayoshi, M.; Punjabi, N.M.; Selvin, E.; Pankow, J.S.; Shahar, E.; Iso, H.; Folsom, A.R.; Lutsey, P.L. Obstructive sleep apnea and incident type 2 diabetes. Sleep Med. 2016, 25, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Appleton, S.L.; Vakulin, A.; McEvoy, R.D.; Wittert, G.A.; Martin, S.A.; Grant, J.F.; Taylor, A.W.; Antic, N.A.; Catcheside, P.G.; Adams, R.J. Nocturnal hypoxemia and severe obstructive sleep apnea are associated with incident type 2 diabetes in a population cohort of men. J. Clin. Sleep Med. 2015, 11, 609–614. [Google Scholar] [CrossRef]
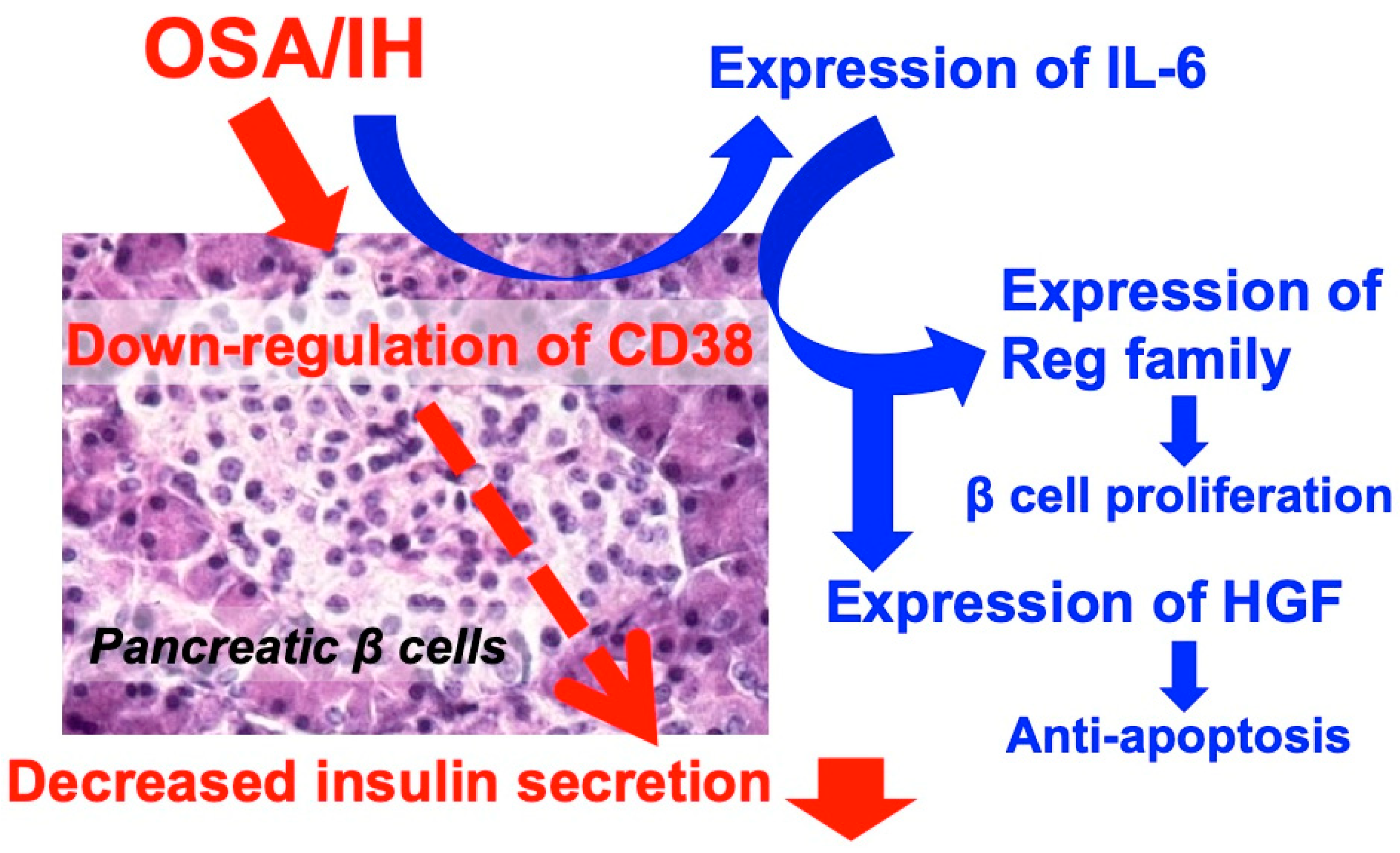
- Ota, H.; Tamaki, S.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Morioka, T.; Takasawa, S.; Kimura, H. Attenuation of glucose-induced insulin secretion by intermittent hypoxia via down-regulation of CD38. Life Sci. 2012, 30, 206–211. [Google Scholar] [CrossRef]
- Okamoto, H.; Takasawa, S. Recent advances in the Okamoto model: The CD38-cyclic ADP-ribose signal system and the regenerating gene protein (Reg)-Reg receptor system in β-cells. Diabetes 2002, 51, S462–S473. [Google Scholar] [CrossRef]
- Okamoto, H.; Takasawa, S.; Yamamoto, Y. From insulin synthesis to secretion: Alternative splicing of type 2 ryanodine receptor gene is essential for insulin secretion in pancreatic β cells. Int. J. Biochem. Cell Biol. 2017, 91, 176–183. [Google Scholar] [CrossRef]
- Takasawa, S. Regenerating gene (REG) product and its potential clinical usage. Expert Opin. Ther. Targets 2016, 20, 541–550. [Google Scholar] [CrossRef]
- Nakagawa, K.; Takasawa, S.; Nata, K.; Yamauchi, A.; Itaya-Hironaka, A.; Ota, H.; Yoshimoto, K.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Takeda, M.; et al. Prevention of Reg I-induced β-cell apoptosis by IL-6/dexamethasone through activation of HGF gene regulation. Biochim. Biophys. Acta 2013, 1833, 2988–2995. [Google Scholar] [CrossRef]
- Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Fujimura, T.; Tsujinaka, H.; Yoshimoto, K.; Nakagawara, K.; Tamaki, S.; et al. Pancreatic β cell proliferation by intermittent hypoxia via up-regulation of Reg family genes and HGF gene. Life Sci. 2013, 93, 664–672. [Google Scholar] [CrossRef]
- Ikehata, F.; Satoh, J.; Nata, K.; Tohgo, A.; Nakazawa, T.; Kato, I.; Kobayashi, S.; Akiyama, T.; Takasawa, S.; Toyota, T.; et al. Autoantibodies against CD38 (ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase) that impair glucose-induced insulin secretion in insulin-dependent diabetes patients. J. Clin. Investig. 1998, 102, 395–401. [Google Scholar] [CrossRef]
- Yagui, K.; Shimada, F.; Miura, M.; Hashimoto, N.; Suzuki, Y.; Tokuyama, Y.; Nata, K.; Tohgo, A.; Ikehata, F.; Takasawa, S.; et al. A missense mutation in CD38 gene, a novel factor for insulin secretion: Association with Type II diabetes mellitus in Japanese subjects and evidence of abnormal function when expressed in vitro. Diabetologia 1998, 41, 1024–1028. [Google Scholar] [CrossRef]
- Pupilli, C.; Giannini, S.; Marchetti, P.; Lupi, R.; Antonelli, A.; Malavasi, F.; Takasawa, S.; Okamoto, H.; Ferrannini, E. Autoantibodies to CD38 (ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase) in Caucasian patients with diabetes: Effects on insulin release from human islets. Diabetes 1999, 48, 2309–2315. [Google Scholar] [CrossRef]
- Shervani, N.J.; Takasawa, S.; Uchigata, Y.; Akiyama, T.; Nakagawa, K.; Noguchi, N.; Takada, H.; Takahashi, I.; Yamauchi, A.; Ikeda, T.; et al. Autoantibodies to REG. a beta-cell regeneration factor, in diabetic patients. Eur. J. Clin. Investig. 2004, 34, 752–758. [Google Scholar] [CrossRef]
- Antonelli, A.; Fallahi, P.; Nesti, C.; Pupilli, C.; Marchetti, P.; Takasawa, S.; Okamoto, H.; Ferrannini, E. Anti-CD38 autoimmunity in patients with chronic autoimmune thyroiditis or Graves’ disease. Clin. Exp. Immunol. 2001, 126, 426–431. [Google Scholar] [CrossRef]
- Pavón, E.J.; Zumaquero, E.; Rosal-Vela, A.; Khoo, K.M.; Cerezo-Wallis, D.; García-Rodríguez, S.; Carrascal, M.; Abian, J.; Graeff, R.; Callejas-Rubio, J.L.; et al. Increased CD38 expression in T cells and circulating anti-CD38 IgG autoantibodies differentially correlate with distinct cytokine profiles and disease activity in systemic lupus erythematosus patients. Cytokine 2013, 62, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, K.; Fujimoto, T.; Itaya-Hironaka, A.; Miyaoka, T.; Sakuramoto-Tsuchida, S.; Yamauchi, A.; Takeda, M.; Kasai, T.; Nakagawara, K.; Nonomura, A.; et al. Involvement of autoimmunity to REG, a regenerating factor, in patients with primary Sjögren’s syndrome. Clin. Exp. Immunol. 2013, 174, 1–9. [Google Scholar] [CrossRef]
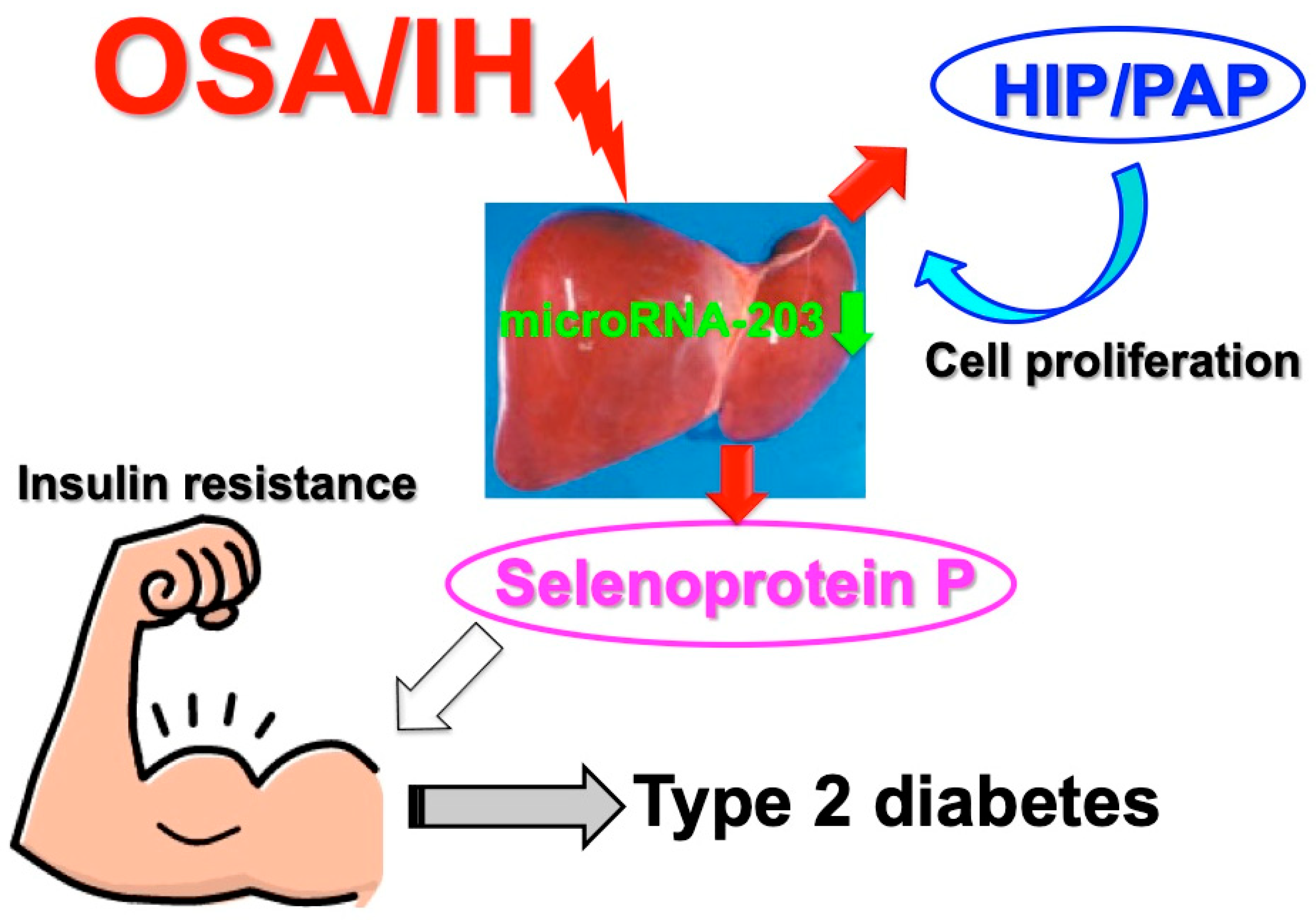
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef]
- Misu, H.; Takamura, T.; Takayama, H.; Hayashi, H.; Matsuzawa-Nagata, N.; Kurita, S.; Ishikura, K.; Ando, H.; Takeshita, Y.; Ota, T.; et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010, 12, 483–495. [Google Scholar] [CrossRef]
- Lieu, H.-T.; Batteux, F.; Simon, M.-T.; Cortes, A.; Nicco, C.; Zavala, F.; Pauloin, A.; Tralhao, J.G.; Soubrane, O.; Weill, B.; et al. HIP/PAP accelerates liver regeneration and protects against acetoaminophen injury in mice. Hepatology 2005, 42, 618–626. [Google Scholar] [CrossRef]
- Muraki, I.; Wada, H.; Tanigawa, T. Sleep apnea and type 2 diabetes. J. Diabetes Investig. 2018, 9, 991–997. [Google Scholar] [CrossRef]
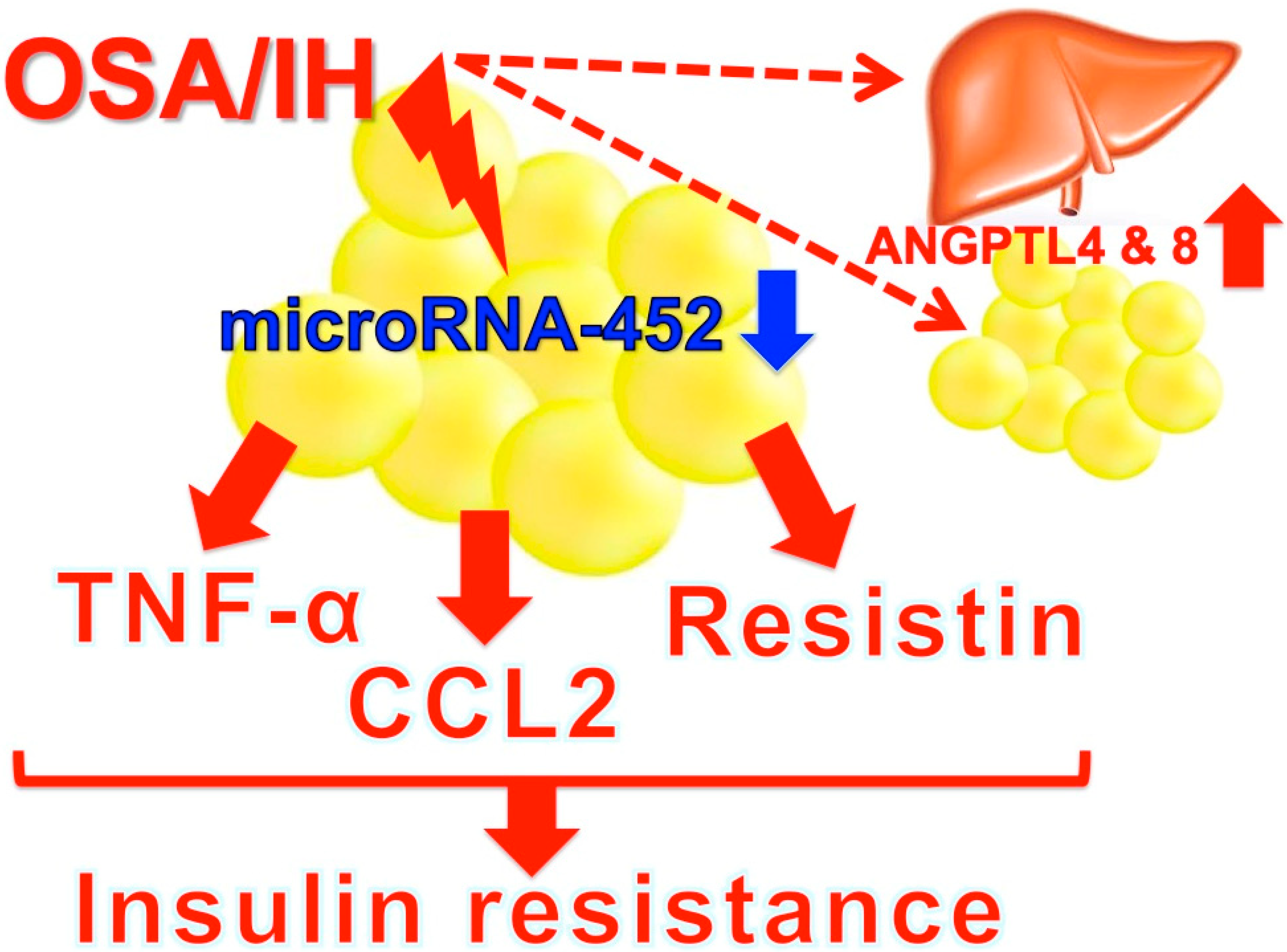
- Al-Terki, A.; Abu-Farha, M.; AlKhairi, I.; Cherian, P.T.; Sriraman, D.; Shyamsundar, A.; Ali, S.; Almulla, F.; Tuomilehto, J.; Abubaker, J.A. Increased level of angiopoietin like proteins 4 and 8 in people with sleep apnea. Front. Endocrinol. 2018, 9, 651. [Google Scholar] [CrossRef]
- Uchiyama, T.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Shobatake, R.; Ota, H.; Takeda, M.; Ohbayashi, C.; Takasawa, S. Intermittent hypoxia up-regulates CCL2, RETN, and TNFα mRNAs in adipocytes via down-regulation of mR-452. Int. J. Mol. Sci. 2019, 20, 1960. [Google Scholar] [CrossRef]
- Romero-Corral, A.; Caples, S.M.; Lopez-Jimenez, F.; Somers, V.K. Interactions between obesity and obstructive sleep apnea: Implications for treatment. Chest 2010, 137, 711–719. [Google Scholar] [CrossRef]
- Mesarwi, O.A.; Sharma, E.V.; Jun, J.C.; Polotsky, V.Y. Metabolic dysfunction in obstructive sleep apnea: A critical examination of underlying mechanisms. Sleep Biol. Rhythms 2015, 13, 2–17. [Google Scholar] [CrossRef]
- Li, K.K.; Kushida, C.; Powell, N.B.; Riley, R.W.; Guilleminault, C. Obstructive sleep apnea syndrome: A comparison between Far-East Asian and white men. Laryngoscope 2000, 110, 1689–1693. [Google Scholar] [CrossRef]
- Shobatake, R.; Takasawa, K.; Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Makino, M.; Sugie, K.; Takasawa, S.; et al. Up-regulation of POMC and CART mRNAs by intermittent hypoxia via GATA transcription factors in human neuronal cells. Int. J. Biochem. Cell Biol. 2018, 95, 100–107. [Google Scholar] [CrossRef]
- Konturek, S.J.; Konturek, J.W.; Pawlik, T.; Brzozowki, T. Brain-gut axis and the role in the control of food intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar]
- Shobatake, R.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Takahashi, N.; Ueno, S.; Sugie, K.; et al. Intermittent hypoxia up-regulates gene expressions of peptide YY (PYY), glucagon-like peptide-1 (GLP-1), and neurotensin (NTS) in enteroendocrine cells. Int. J. Mol. Sci. 2019, 20, 1849. [Google Scholar] [CrossRef]
- WHO. Attaining the Nine Global Noncommunicable Diseases Targets; A Shared Responsicility; Global Status Report on Noncommunicable Diseases; World Health Organization: Geneva, Switzerland, 2014; Volume 176. [Google Scholar]
- Esposito, K.; Chiodini, P.; Colao, A.; Lenzi, A.; Giugliano, D. Metabolic syndrome and risk of cancer: A systematic review and meta-analysis. Diabetes Care 2012, 35, 2402–2411. [Google Scholar] [CrossRef]
- Calle, E.E.; Rodriguez, C.; Walker-Thurmond, K.; Thun, M.J. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med. 2003, 348, 1625–1638. [Google Scholar] [CrossRef]
- Samson, S.L.; Garber, A.J. Metabolic syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23. [Google Scholar] [CrossRef]
- Iiyori, N.; Alonso, L.C.; Li, J.; Sanders, M.H.; Garcia-Ocana, A.; O’Doherty, R.M.; Polotsky, V.Y.; O’Donnell, C.P. Intermittent hypoxia causes insulin resistance in lean mice independent of autonomic activity. Am. J. Respir. Crit. Care Med. 2007, 175, 851–857. [Google Scholar] [CrossRef]
- Mackenzie, R.; Maxwell, N.; Castle, P.; Brickley, G.; Watt, P. Acute hypoxia and exercise improve insulin sensitivity (S(I) (2*)) in individuals with type 2 diabetes. Diabetes Metab. Res. Rev. 2011, 27, 94–101. [Google Scholar] [CrossRef]
- Lecoultre, V.; Peterson, C.M.; Covington, J.D.; Ebenezer, P.J.; Frost, E.A.; Schwarz, J.M.; Ravussin, E. Ten nights of moderate hypoxia improves insulin sensitivity in obese humans. Diabetes Care 2013, 36, e197–e198. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.; Duan, Y.; Hu, C.A.; Tang, Y.; Yin, T. Myokines and adipokines: Involvement in the crosstalk between skeletal muscle and adipose tissue. Cytokine Growth Factor Rev. 2017, 33, 73–82. [Google Scholar] [CrossRef]
- Sorajja, D.; Gami, A.S.; Somers, V.K.; Behrenbeck, T.R.; Garcia-Touchard, A.; Lopez-Jimenez, F. Independent association between obstructive sleep apnea and subclinical coronary artery disease. Chest 2008, 133, 927–933. [Google Scholar] [CrossRef]
- Wu, C.-J.; Kao, T.-W.; Chen, Y.-W.; Chen, W.-L. Examining the association between obstructive sleep apnea and cardiometabolic risk factors in the elderly. Sleep Biol. Rhythms 2018, 16, 231–234. [Google Scholar] [CrossRef]
- Kyotani, Y.; Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Zhao, J.; Ozawa, K.; Nagayama, K.; Ito, S.; Takasawa, S.; et al. Intermittent hypoxia induces the proliferation of rat vascular smooth muscle cell with the increases in epidermal growth factor family and erbB2 receptor. Exp. Cell Res. 2013, 319, 3042–3050. [Google Scholar] [CrossRef]
- Kyotani, Y.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Takasawa, S.; Yoshizumi, M. Intermittent hypoxia-induced epiregulin expression by IL-6 production in human coronary artery smooth muscle cells. FEBS Open Bio 2018, 8, 868–876. [Google Scholar] [CrossRef]
- Kyotani, Y.; Takasawa, S.; Yoshizumi, M. Proliferative pathways of vascular smooth muscle cells by intermittent hypoxia. Int. J. Mol. Sci. 2019, 20, 2706. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimura, H.; Ota, H.; Kimura, Y.; Takasawa, S. Effects of Intermittent Hypoxia on Pulmonary Vascular and Systemic Diseases. Int. J. Environ. Res. Public Health 2019, 16, 3101. https://doi.org/10.3390/ijerph16173101
Kimura H, Ota H, Kimura Y, Takasawa S. Effects of Intermittent Hypoxia on Pulmonary Vascular and Systemic Diseases. International Journal of Environmental Research and Public Health. 2019; 16(17):3101. https://doi.org/10.3390/ijerph16173101
Chicago/Turabian StyleKimura, Hiroshi, Hiroyo Ota, Yuya Kimura, and Shin Takasawa. 2019. "Effects of Intermittent Hypoxia on Pulmonary Vascular and Systemic Diseases" International Journal of Environmental Research and Public Health 16, no. 17: 3101. https://doi.org/10.3390/ijerph16173101
APA StyleKimura, H., Ota, H., Kimura, Y., & Takasawa, S. (2019). Effects of Intermittent Hypoxia on Pulmonary Vascular and Systemic Diseases. International Journal of Environmental Research and Public Health, 16(17), 3101. https://doi.org/10.3390/ijerph16173101