Icariin Treatment Enhanced the Skeletal Response to Exercise in Estrogen-Deficient Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Intervention Protocols
2.2. Bone Density and Morphometry Analyses
2.3. Bone Mesenchymal Stem Cell Culture
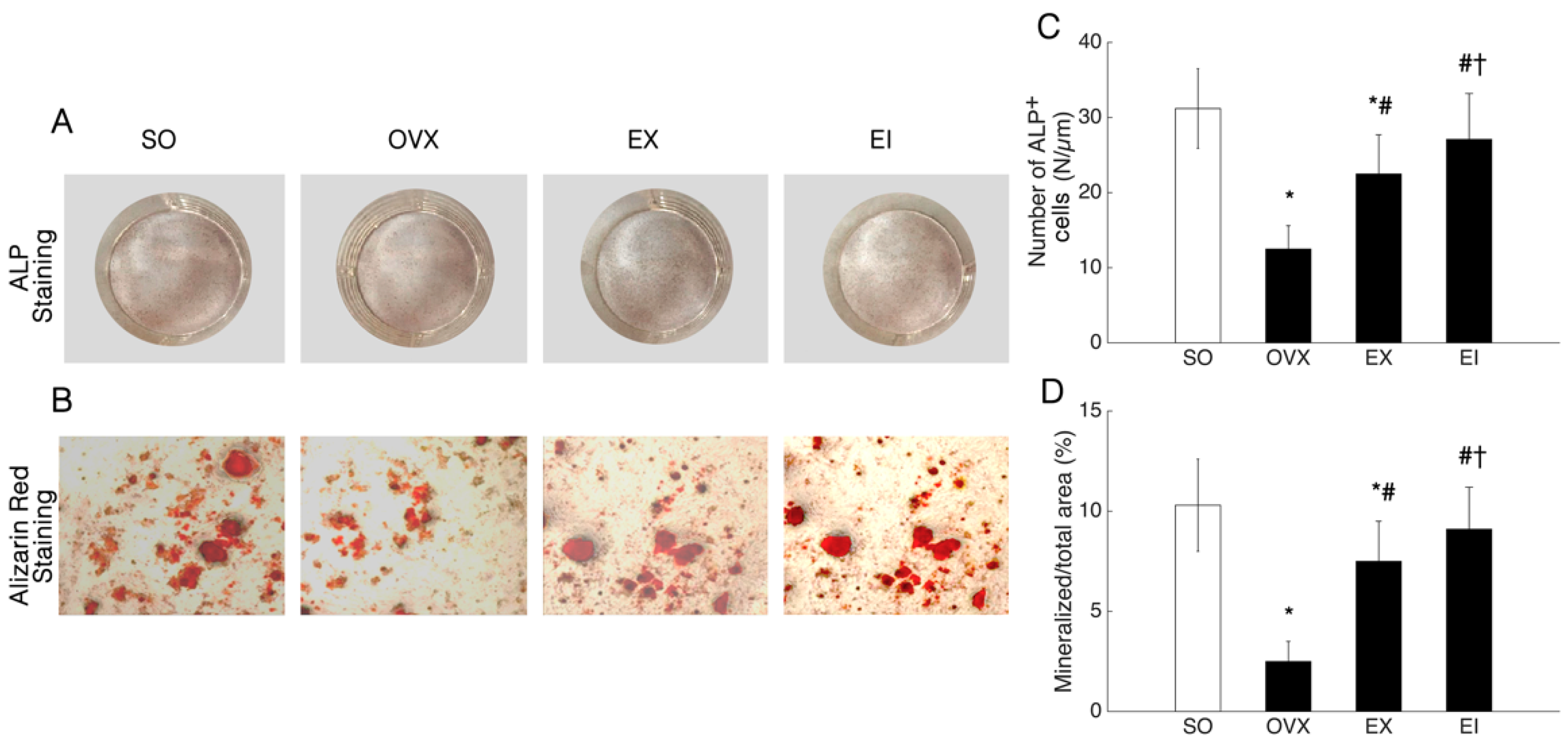
2.4. Alkaline Phosphatase Staining and Alizarin Red Staining
2.5. Serum E2, ALP and Tartrate-Resistant Acid Phosphatase Analyses
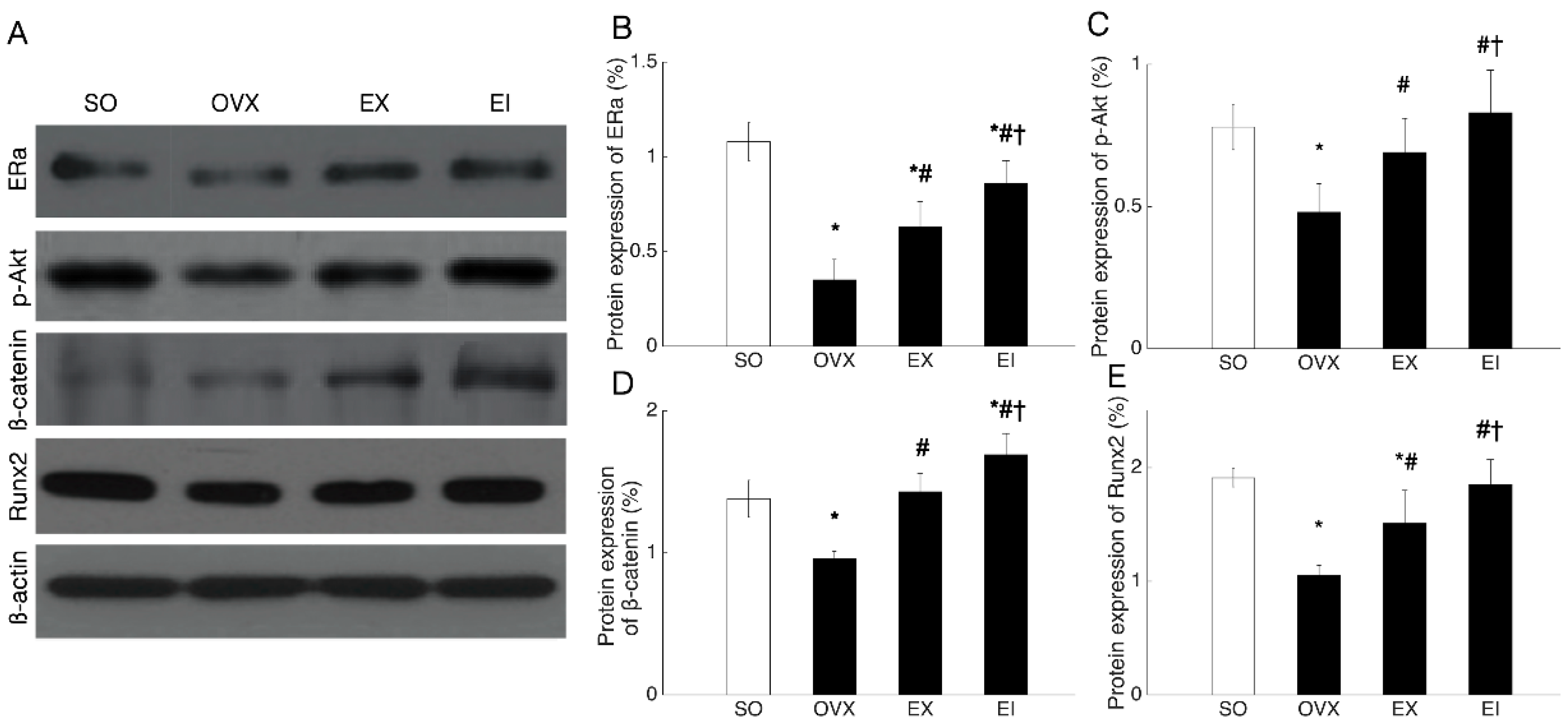
2.6. Western Blot Analysis
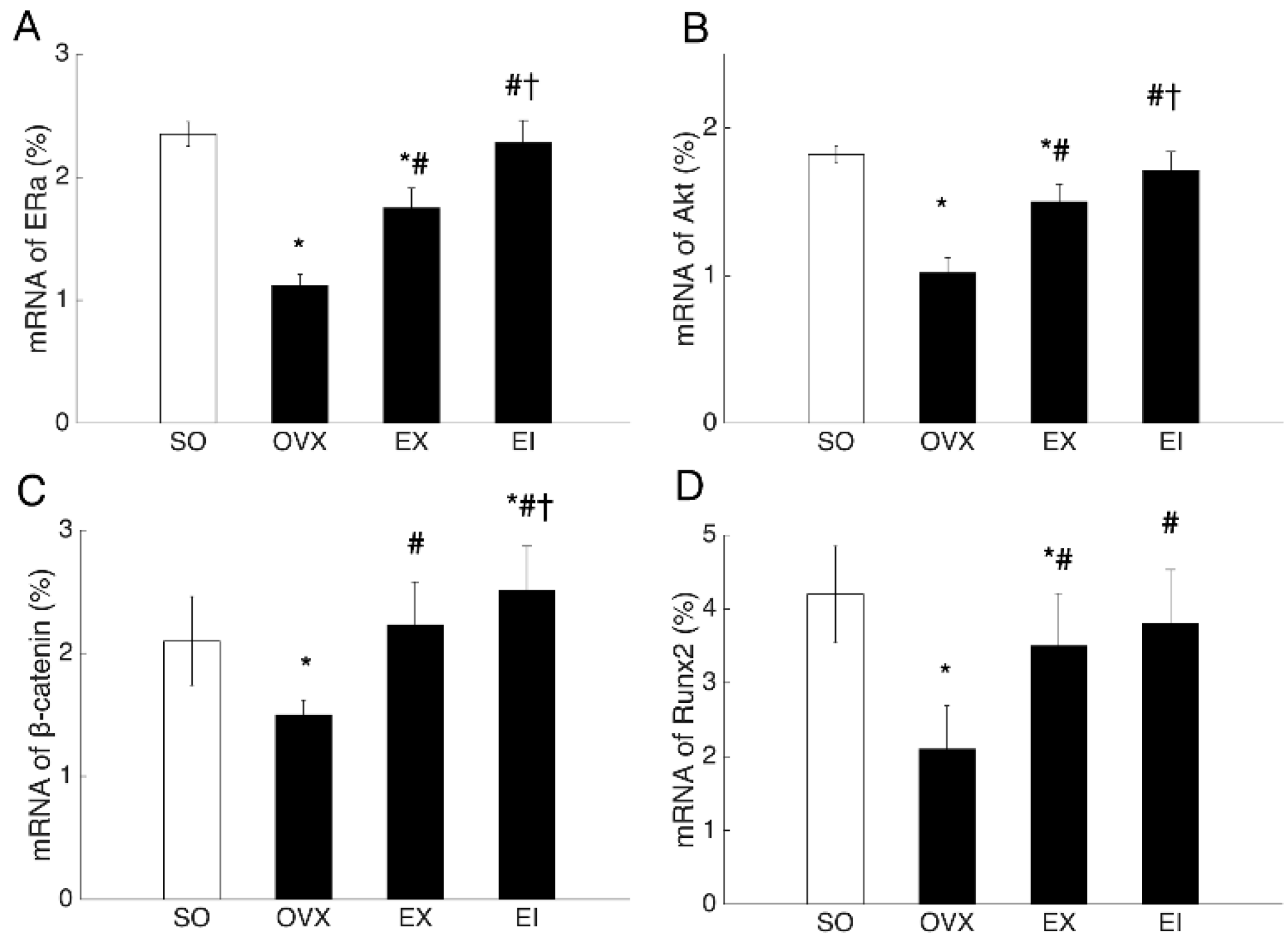
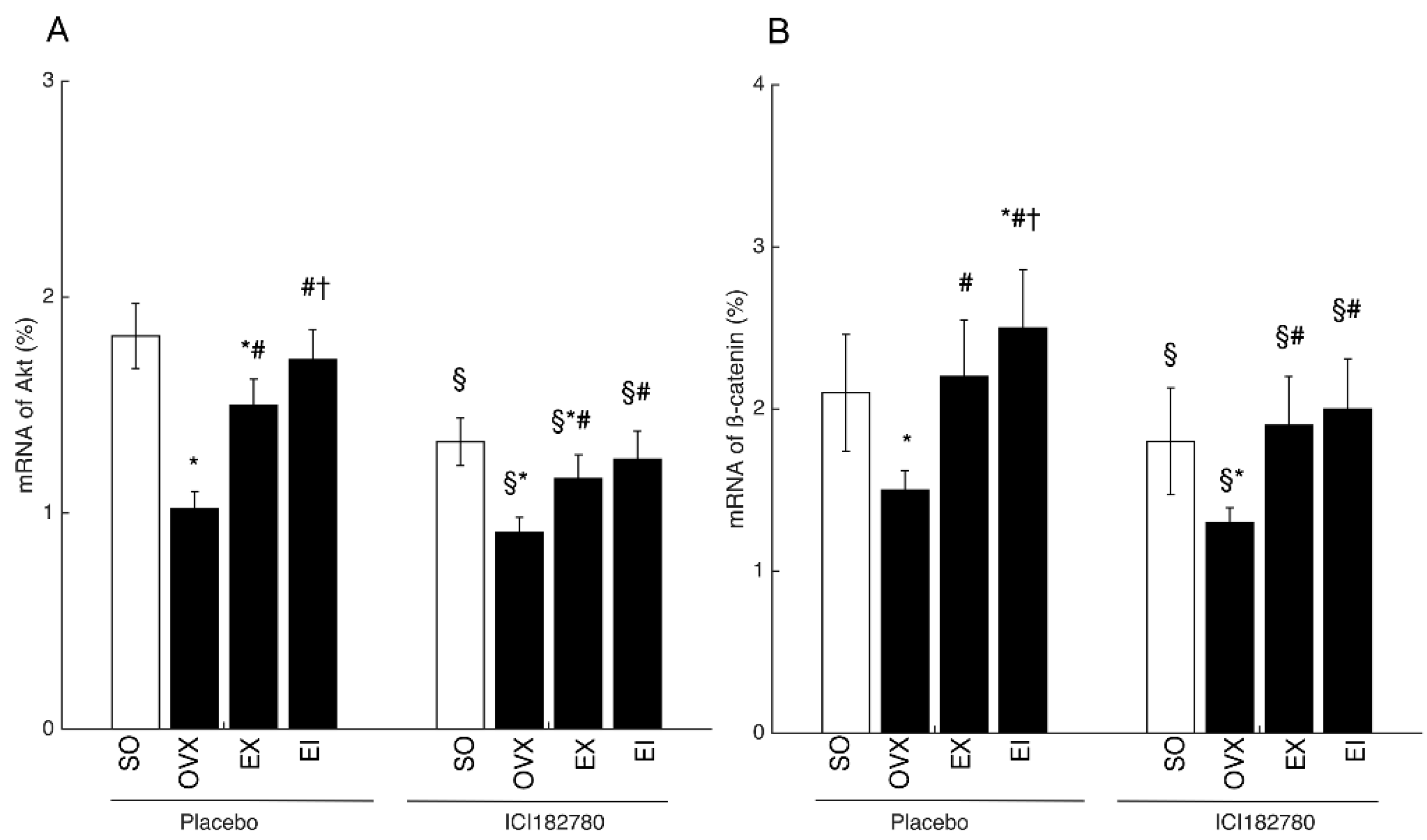
2.7. RT-PCR Analysis
2.8. StatisticalAnalysis
3. Results
3.1. General Characteristics
3.2. Bone Tissue Characteristic Analysis
3.3. Osteogenic Differentiation and Deposition
3.4. Western Blot and PCR Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Van den Bergh, J.P.; van Geel, T.A.; Geusens, P.P. Osteoporosis, frailty and fracture: Implications for case finding and therapy. Nat. Rev. Rheumatol. 2012, 8, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, R.S.; Col, N.F.; Wong, J.B. Estimating hip fracture morbidity, mortality and costs. J. Am. Geriatr. Soc. 2003, 51, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; Melton, L.J. Epidemiology and outcomes of osteoporotic fractures. Lancet 2002, 359, 1761–1767. [Google Scholar] [CrossRef]
- Cotts, K.G.; Cifu, A.S. Treatment of Osteoporosis. JAMA 2018, 319, 1040–1041. [Google Scholar] [CrossRef]
- Black, D.M.; Rosen, C.J. Clinical Practice. Postmenopausal Osteoporosis. N. Engl. J. Med. 2016, 374, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Kemmler, W.; Engelke, K.; Weineck, J.; Hensen, J.; Kalender, W.A. The Erlangen Fitness Osteoporosis Prevention Study: A controlled exercise trial in early postmenopausal women with low bone density-first-year results. Arch. Phys. Med. Rehabil. 2003, 84, 673–682. [Google Scholar]
- Martyn-St James, M.; Carroll, S. High-intensity resistance training and postmenopausal bone loss: A meta-analysis. Osteoporos. Int. 2006, 17, 1225–1240. [Google Scholar] [CrossRef]
- Snow, C.M.; Shaw, J.M.; Winters, K.M.; Witzke, K.A. Long-term exercise using weighted vests prevents hip bone loss in postmenopausal women. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2000, 55, M489-91. [Google Scholar] [CrossRef]
- Martyn-St James, M.; Carroll, S. A meta-analysis of impact exercise on postmenopausal bone loss: The case for mixed loading exercise programmes. Br. J. Sports Med. 2009, 43, 898–908. [Google Scholar] [CrossRef]
- Kelley, G.A.; Kelley, K.S.; Kohrt, W.M. Effects of ground and joint reaction force exercise on lumbar spine and femoral neck bone mineral density in postmenopausal women: A meta-analysis of randomized controlled trials. BMC Musculoskelet. Disord. 2012, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Howe, T.E.; Shea, B.; Dawson, L.J.; Downie, F.; Murray, A.; Ross, C.; Harbour, R.T.; Caldwell, L.M.; Creed, G. Exercise for preventing and treating osteoporosis in postmenopausal women. Cochrane Database Syst. Rev. 2011, 7, CD000333. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Feng, F.; Wang, X. Exercise interventions and prevention of fall-related fractures in older people: A meta-analysis of randomized controlled trials. Int. J. Epidemiol. 2017, 46, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Zhao, M.; Xu, Z. The effects of differing resistance training modes on the preservation of bone mineral density in postmenopausal women: A meta-analysis. Osteoporos. Int. 2015, 26, 1605–1618. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Jessop, H.; Suswillo, R.; Zaman, G.; Lanyon, L. Endocrinology: Bone adaptation requires oestrogen receptor-alpha. Nature 2003, 424, 389. [Google Scholar] [CrossRef] [PubMed]
- Tobias, J.H. At the crossroads of skeletal responses to estrogen and exercise. Trends Endocrinol. Metab. 2003, 14, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Ankrom, M.A.; Patterson, J.A.; d’Avis, P.Y.; Vetter, U.K.; Blackman, M.R.; Sponseller, P.D.; Tayback, M.; Robey, P.G.; Shapiro, J.R.; Fedarko, N.S. Age-related changes in human oestrogen receptor alpha function and levels in osteoblasts. Biochem. J. 1998, 333 Pt 3, 787–794. [Google Scholar] [CrossRef]
- Lobo, R.A. Hormone-replacement therapy: Current thinking. Nat. Rev. Endocrinol. 2017, 13, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.D. Assessing benefits and harms of hormone replacement therapy: Clinical applications. JAMA 2002, 288, 882–884. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.L.; Liang, B.C.; Wu, P.; Wang, B.; Shi, Z.Y.; Kong, L.C.; Yao, J.L.; Huang, J.J.; Li, C.W. Effect and mechanism of total flavone of epimedium on primary callus formation in ovariectomized rats with fractures. China J. Orthop. Traumatol. 2017, 30, 743–750. [Google Scholar]
- Indran, I.R.; Liang, R.L.; Min, T.E.; Yong, E.L. Preclinical studies and clinical evaluation of compounds from the genus Epimedium for osteoporosis and bone health. Pharmacol. Ther. 2016, 162, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, Y.; Zhang, Y.; Li, A.; Tian, M.; Fan, S.; Yang, S.; Li, Y. Compounds with Estrogen-like Effects from Epimedii Folium Based on Cell Sorption and Molecular Docking. Chin. J. Exp. Tradit. Med. Formulae 2016, 22, 62–66. [Google Scholar]
- Wu, T.; Zhang, Y.; Wen, Y.; Zhang, J. The Effects of Icariin on Serum Levels of Estrogen in Ovariectomized Mice. Guizhou Med. J. 2011, 34, 79–80. [Google Scholar]
- Mok, S.K.; Chen, W.F.; Lai, W.P.; Leung, P.C.; Wang, X.L.; Yao, X.S.; Wong, M.S. Icariin protects against bone loss induced by oestrogen deficiency and activates oestrogen receptor-dependent osteoblastic functions in UMR 106 cells. Br. J. Pharmacol. 2010, 159, 939–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nian, H.; Ma, M.H.; Nian, S.S.; Xu, L.L. Antiosteoporotic activity of icariin in ovariectomized rats. Phytomedicine 2009, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Zhang, J.; Hong, G.; Chen, Z.; Deng, W.; He, W.; Chen, M.H. Icariin promotes osteogenic differentiation of rat bone marrow stromal cells by activating the ERalpha-Wnt/beta-catenin signaling pathway. Biomed. Pharmacother. 2016, 84, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.; Chiba, H.; Higuchi, M.; Nakatani, T.; Ezaki, O.; Cui, H.; Yamada, K.; Ishimi, Y. Combined intervention of soy isoflavone and moderate exercise prevents body fat elevation and bone loss in ovariectomized mice. Metabolism 2004, 53, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.H.; Kapur, S.; Kesavan, C.; Baylink, D.J. Upregulation of the Wnt, estrogen receptor, insulin-like growth factor-I, and bone morphogenetic protein pathways in C57BL/6J osteoblasts as opposed to C3H/HeJ osteoblasts in part contributes to the differential anabolic response to fluid shear. J. Biol. Chem. 2006, 281, 9576–9588. [Google Scholar] [CrossRef]
- Lennon, D.P.; Caplan, A.I. Isolation of rat marrow-derived mesenchymal stem cells. Exp. Hematol. 2006, 34, 1606–1607. [Google Scholar] [CrossRef]
- Bu, S.; Chen, Y.; Wang, S.; Zhang, F.; Ji, G. Treadmill training regulates beta-catenin signaling through phosphorylation of GSK-3beta in lumbar vertebrae of ovariectomized rats. Eur. J. Appl. Physiol. 2012, 112, 3295–3304. [Google Scholar] [CrossRef]
- Armstrong, V.J.; Muzylak, M.; Sunters, A.; Zaman, G.; Saxon, L.K.; Price, J.S.; Lanyon, L.E. Wnt/beta-catenin signaling is a component of osteoblastic bone cell early responses to load-bearing and requires estrogen receptor alpha. J. Biol. Chem. 2007, 282, 20715–20727. [Google Scholar] [CrossRef] [PubMed]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | SO | OVX | EX | EI |
|---|---|---|---|---|
| Weight pre (g) | 210.70 ± 14.95 | 212.46 ± 14.60 | 211.01 ± 9.21 | 209.25 ± 14.69 |
| Weight post (g) | 288.36 ± 15.51 | 323.65 ± 16.09 * | 307.48 ± 12.94 *# | 302.67 ± 13.72 *# |
| BMD spine (g/cm2) | 0.175 ± 0.01 | 0.155 ± 0.011 * | 0.162 ± 0.01 *# | 0.171 ± 0.01 #† |
| Serum E2 (pg/mL) | 23.61 ± 2.27 | 12.94 ± 2.95 * | 17.33 ± 1.81 *# | 20.50 ± 1.98 *#† |
| Serum ALP (IU/dl) | 9.67 ± 2.82 | 15.69 ± 3.69 * | 12.52 ± 2.75 *# | 9.39 ± 2.12 #† |
| Serum TRAP (IU/dl) | 39.23 ± 6.21 | 69.12 ± 8.61 * | 51.25 ± 7.83 *# | 40.53 ± 6.39 #† |
| Variables | SO | OVX | EX | EI |
|---|---|---|---|---|
| BV/TV (%) | 59.2 ± 8.5 | 32.6 ± 6.3 * | 45.2 ± 7.2 *# | 55.5 ± 7.6 #† |
| Tb.Th (mm) | 83.1 ± 13.5 | 63.5 ± 7.3 * | 78.3 ± 12.1 # | 80.9 ± 11.1 # |
| Tb.N (N/mm2) | 13.2 ± 2.1 | 7.3 ± 1.9 * | 10.1 ± 2.5 *# | 12.8 ± 2.9 # |
| Tb.Sp (mm) | 103.3 ± 19.5 | 149.2 ± 27.3 * | 128.5 ± 21.8 # | 106.8 ± 23.6 #† |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Bu, W.; Chen, Y. Icariin Treatment Enhanced the Skeletal Response to Exercise in Estrogen-Deficient Rats. Int. J. Environ. Res. Public Health 2019, 16, 3779. https://doi.org/10.3390/ijerph16193779
Zhao R, Bu W, Chen Y. Icariin Treatment Enhanced the Skeletal Response to Exercise in Estrogen-Deficient Rats. International Journal of Environmental Research and Public Health. 2019; 16(19):3779. https://doi.org/10.3390/ijerph16193779
Chicago/Turabian StyleZhao, Renqing, Wenqian Bu, and Yingfeng Chen. 2019. "Icariin Treatment Enhanced the Skeletal Response to Exercise in Estrogen-Deficient Rats" International Journal of Environmental Research and Public Health 16, no. 19: 3779. https://doi.org/10.3390/ijerph16193779
APA StyleZhao, R., Bu, W., & Chen, Y. (2019). Icariin Treatment Enhanced the Skeletal Response to Exercise in Estrogen-Deficient Rats. International Journal of Environmental Research and Public Health, 16(19), 3779. https://doi.org/10.3390/ijerph16193779