Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs


Abstract
:1. Introduction
2. Materials and Methods
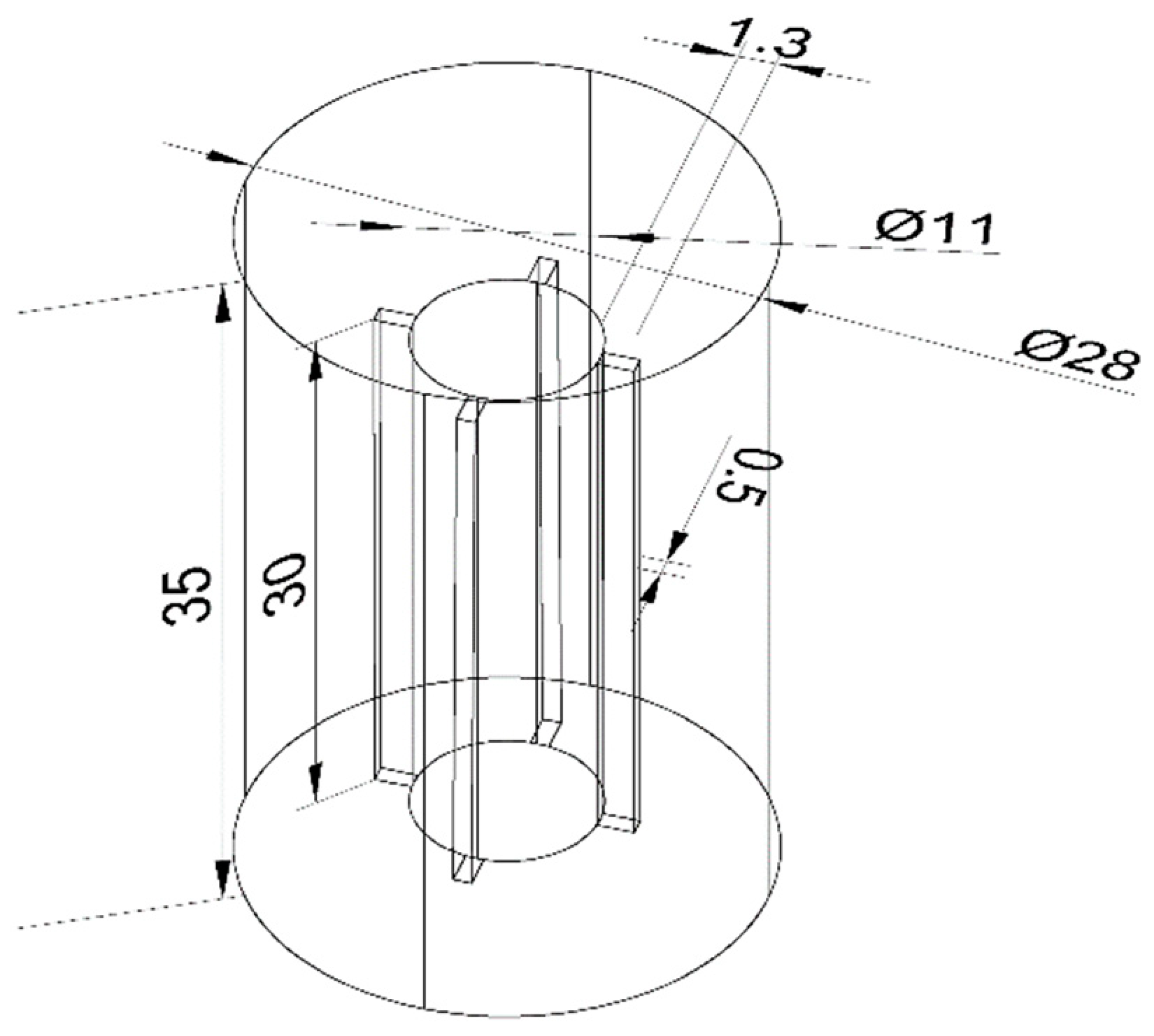
2.1. Algae and Experimental Apparatus
2.2. Experimental Design and Analytical Methods
2.3. Statistical Analysis
3. Results and Discussion
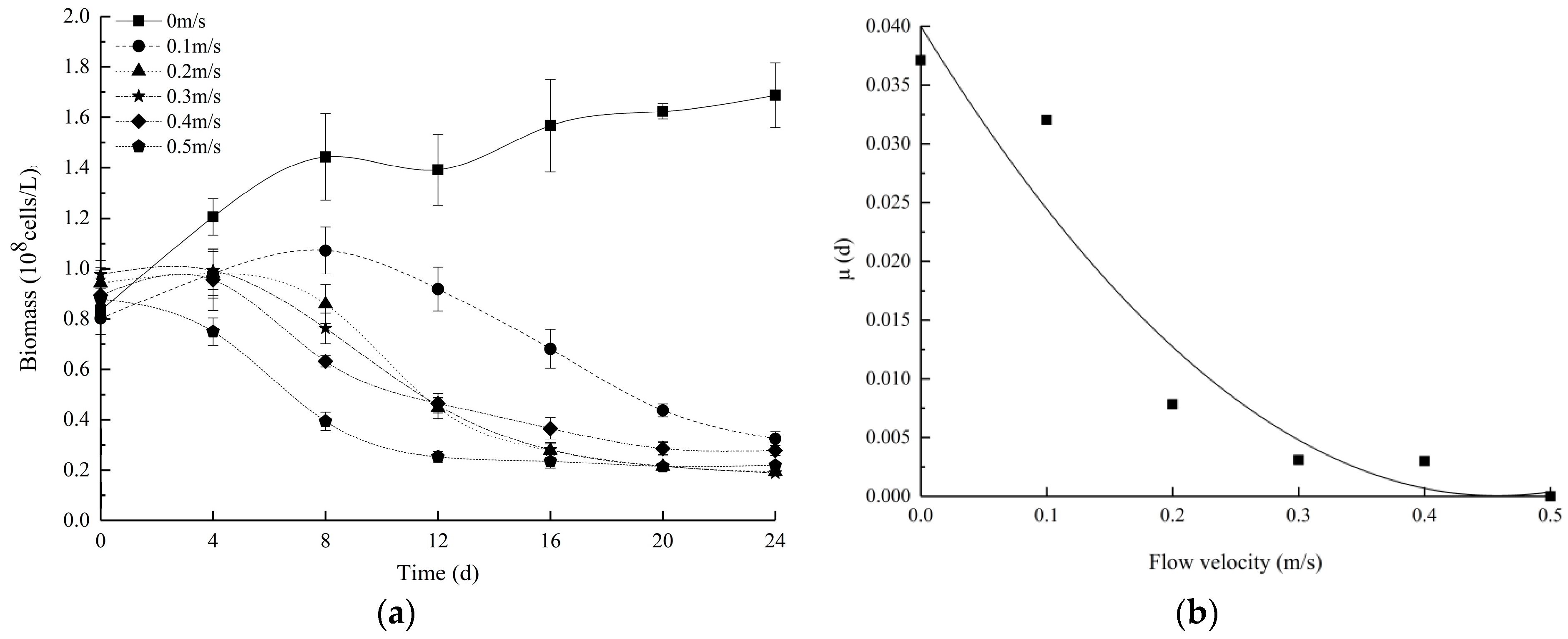
3.1. Changes in the Growth Status of Euglena at Different Flow Velocities
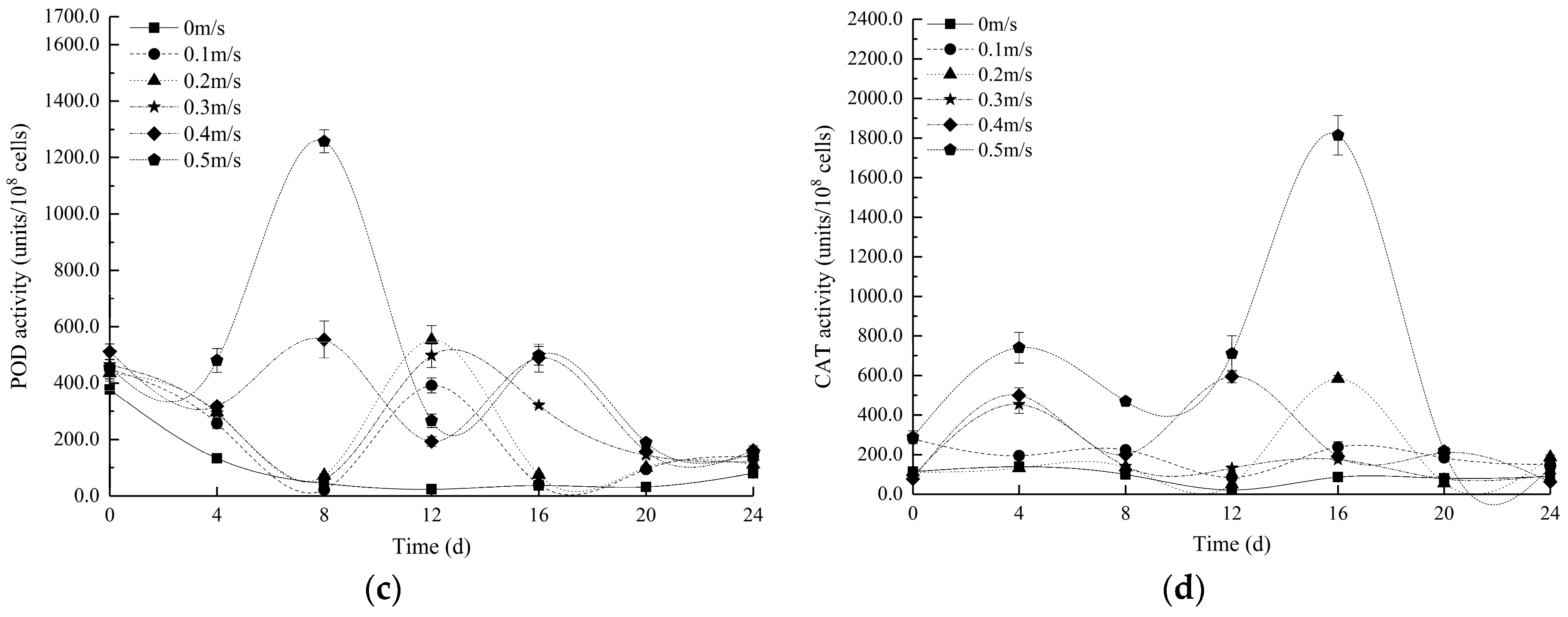
3.2. Changes in Lipid Oxidation and the Antioxidant Enzyme Activity of Euglena at Different Flow Velocities
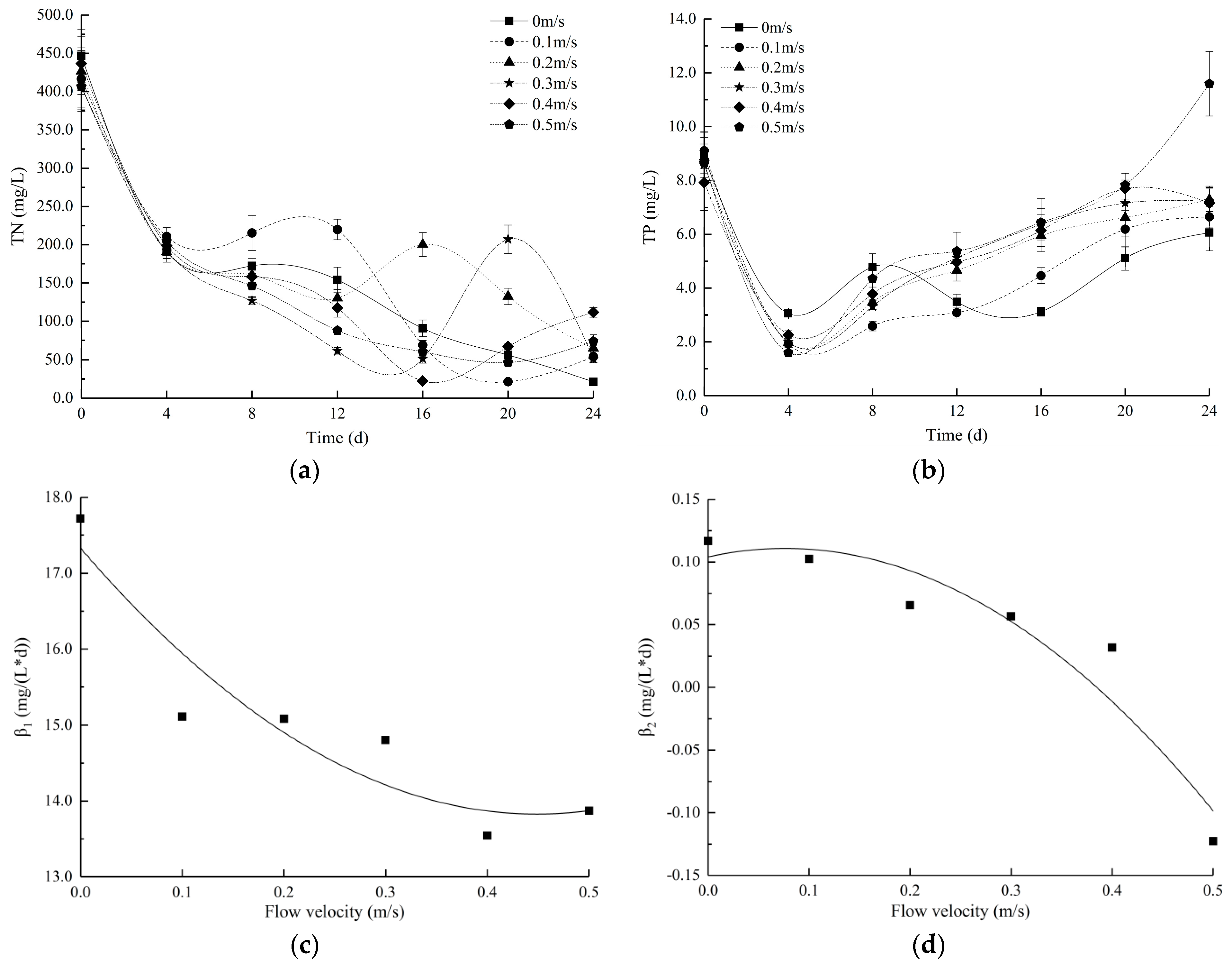
3.3. Changes in the Nutrient Levels and Extracellular Hydrolase Activities in Water at Different Flow Velocities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Herman, B.D.; Eberly, J.O.; Jung, C.M.; Medina, V.F. Review and Evaluation of Reservoir Management Strategies for Harmful Algal Blooms (No. ERDC/EL TR-17-11); ERDC-EL: Vicksburg, MS, USA, 2017. [Google Scholar]
- Mao, J.; Jiang, D.; Dai, H. Spatial–temporal hydrodynamic and algal bloom modelling analysis of a reservoir tributary embayment. J. Hydro-Environ. Res. 2015, 9, 200–215. [Google Scholar] [CrossRef]
- Sherman, B.S.; Webster, I.T.; Jones, G.J.; Oliver, R.L. Transitions between Aulacoseira and Anabaena dominance in a turbid river weir pool. Limnol. Oceanogr. 1998, 43, 1902–1915. [Google Scholar] [CrossRef]
- Lappalainen, J.; Virtanen, E.A.; Kallio, K.; Junttila, S.; Viitasalo, M. Substrate limitation of a habitat-forming genus Fucus under different water clarity scenarios in the northern Baltic Sea. Estuar. Coast. Shelf Sci. 2019, 218, 31–38. [Google Scholar] [CrossRef]
- Finlay, J.C.; Power, M.E.; Cabana, G. Effects of water velocity on algal carbon isotope ratios: Implications for river food web studies. Limnol. Oceanogr. 1999, 44, 1198–1203. [Google Scholar] [CrossRef]
- Li, Z.; Wang, S.; Guo, J.S.; Fang, F.; Gao, X.; Long, M. Responses of phytoplankton diversity to physical disturbance under manual operation in a large reservoir, China. Hydrobiologia 2012, 684, 45–56. [Google Scholar] [CrossRef]
- Hadiyanto, H.; Elmore, S.; Van Gerven, T.; Stankiewicz, A. Hydrodynamic evaluations in high rate algae pond (HRAP) design. Chem. Eng. J. 2013, 217, 231–239. [Google Scholar] [CrossRef]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria–ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Xiao, Y.; Li, Z.; Li, C.; Zhang, Z.; Guo, J.S. Effect of small-scale turbulence on the physiology and morphology of two bloom-forming cyanobacteria. PLoS ONE 2016, 11, e0168925. [Google Scholar] [CrossRef]
- Li, H.M.; Tang, H.J.; Shi, X.Y.; Zhang, C.S.; Wang, X.L. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased Harmful Algal Blooms. Harmful Algae 2014, 39, 92–101. [Google Scholar] [CrossRef]
- Long, T.Y.; Wu, L.; Meng, G.H.; Guo, W.H. Numerical simulation for impacts of hydrodynamic conditions on algae growth in Chongqing Section of Jialing River, China. Ecol. Model. 2011, 222, 112–119. [Google Scholar] [CrossRef]
- Ruiz-Villarreal, M.; García-García, L.M.; Cobas, M.; Díaz, P.A.; Reguera, B. Modelling the hydrodynamic conditions associated with Dinophysis blooms in Galicia (NW Spain). Harmful Algae 2016, 53, 40–52. [Google Scholar] [CrossRef] [PubMed]
- King, S.A. Hydrodynamic control of filamentous macroalgae in a sub-tropical spring-fed river in Florida, USA. Hydrobiologia 2014, 734, 27–37. [Google Scholar] [CrossRef]
- Acuna, V.; Vilches, C.; Giorgi, A. As productive and slow as a stream can be the metabolism of a Pampean stream. J. North Am. Benthol. Soc. 2011, 30, 71–83. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Hardwick, L.; Dorani, F. Use of flow management to mitigate cyanobacterial blooms in the Lower Darling River, Australia. J. Plankton Res. 2011, 33, 229–241. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, L.L.; Li, J.; Chen, M.; Zhang, Y.W. Mechanism of the influence of hydrodynamics on Microcystis aeruginosa, a dominant bloom species in reservoirs. Sci. Total Environ. 2018, 636, 230–239. [Google Scholar] [CrossRef]
- Mcintire, C.D.; Garrison, R.L.; Phinney, H.K.; Warren, C.E. Primary production in laboratory streams. Limnol. Oceanogr. 1964, 9, 92–102. [Google Scholar] [CrossRef]
- Alves-de-Souza, C.; Gonzalez, M.T.; Iriarte, J.L. Functional groups in marine phytoplankton assemblages dominated by diatoms in fjords of southern Chile. J. Plankton Res. 2008, 30, 1233–1243. [Google Scholar] [CrossRef]
- Mahapatra, D.M.; Chanakya, H.N.; Ramachandra, T.V. Euglena sp as a suitable source of lipids for potential use as biofuel and sustainable wastewater treatment. J. Appl. Phycol. 2013, 25, 855–865. [Google Scholar] [CrossRef]
- Mutshinda, C.M.; Finkel, Z.V.; Widdicombe, C.E.; Irwin, A.J. Ecological equivalence of species within phytoplankton functional groups. Funct. Ecol. 2016, 30, 1714–1722. [Google Scholar] [CrossRef]
- Ivanova, I.M.; Nepogodiev, S.A.; Saalbach, G.; O’Neill, E.C.; Urbaniak, M.D.; Ferguson, M.A.J.; Gurcha, S.S.; Besra, G.S.; Field, R.A. Fluorescent mannosides serve as acceptor substrates for glycosyltransferase and sugar-1-phosphate transferase activities in Euglena gracilis membranes. Carbohydr. Res. 2017, 438, 26–38. [Google Scholar] [CrossRef]
- Kingston, M.B. Effect of light on vertical migration and photosynthesis of Euglena proxima (Euglenophyta). J. Phycol. 1999, 35, 245–253. [Google Scholar] [CrossRef]
- Kingston, M.B. Wave effects on the vertical migration of two benthic microalgae: Hantzschia virgata var. intermedia and Euglena proxima. Estuaries 1999, 22, 91. [Google Scholar] [CrossRef]
- Jekely, G. Evolution of phototaxis. Philos. Trans. R. Soc. B 2009, 364, 2795–2808. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, M.; Heltai, L.; Millan, D.; DeSimone, A. Reverse engineering the euglenoid movement. Biophys. J. 2012, 109, 17874–17879. [Google Scholar] [CrossRef] [PubMed]
- Giometto, A.; Altermatt, F.; Maritan, A.; Stocker, R.; Rinaldo, A. Generalized receptor law governs phototaxis in the phytoplankton Euglena gracilis. Proc. Natl. Acad. Sci. USA 2015, 112, 7045–7050. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.C.H.; Lam, A.T.; Riedel-Kruse, I.H. Polygonal motion and adaptable phototaxis via flagellar beat switching in the microswimmer Euglena gracilis. Nat. Phys. 2018, 14, 1216–1222. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Li, H.; Li, J.; Song, Y.; Zhang, L.L.; Li, Y.; Pu, X.C.; Huang, W.D. Spatiotemporal Succession Characteristics of Algal Functional Groups and Its Impact Factors for a Typical Channel-Type Reservoir in a Southwest Mountainous Area. Chin. J. Environ. Sci. 2018, 39, 2680–2687. (In Chinese) [Google Scholar]
- Zimba, P.V.; Moeller, P.D.; Beauchesne, K.; Lane, H.E.; Triemer, R.E. Identification of Euglenophycin—A toxin found in certain Euglenoids. Toxicon 2010, 55, 100–104. [Google Scholar] [CrossRef]
- Zimba, P.V.; Huang, I.S.; Gutierrez, D.; Shin, W.; Bennett, M.S.; Triemer, R.E. Euglenophycin is produced in at least six species of Euglenoid algae and six of seven strains of Euglena sanguinea. Harmful Algae 2017, 63, 79–84. [Google Scholar] [CrossRef]
- Ligeza, S.; Wilk-Wozniak, E. The occurrence of a Euglena pascheri and Lepocinclis ovum bloom in an oxbow lake in southern Poland under extreme environmental conditions. Ecol. Indic. 2011, 11, 925–929. [Google Scholar] [CrossRef]
- Kouhanestani, Z.M.; Roelke, D.L.; Ghorbani, R.; Fujiwara, M. Assessment of Spatiotemporal Phytoplankton Composition in Relation to Environmental Conditions of Gorgan Bay, Iran. Estuaries Coasts 2019, 42, 173–189. [Google Scholar] [CrossRef]
- Da Rocha, A.R.; Danilo, G. Limnological characterisation and phytoplankton seasonal variation in a subtropical shallow lake (Guaiba Lake, Brazil): A long-term study. Acta Limnol. Bras. 2014, 26, 442–456. [Google Scholar]
- dos Santos, A.K.D.; Oliveira, A.L.L.; Furtado, J.A.; Ferreira, F.S.; Araujo, B.D.; Correa, J.J.M.; Cavalcanti, L.F.; Cutrim, A.C.G.D.A.; Cutrim, M.V.J. Spatial and seasonal variation of microphytoplankton community and the correlation with environmental parameters in a hypereutrophic tropical estuary—Maranhao—Brazil. Braz. J. Oceanogr. 2017, 65, 356–372. [Google Scholar] [CrossRef]
- Lv, J.; Wu, H.J.; Chen, M.Q. Effects of nitrogen and phosphorus on phytoplankton composition and biomass in 15 subtropical, urban shallow lakes in Wuhan, China. Limnologica 2011, 41, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Hao, D.P.; Pei, H.Y.; Doblin, M.A.; Ren, Y.; Wei, J.L.; Feng, Y.W. Phytoplankton Functional Groups Variation and Influencing Factors in a Shallow Temperate Lake. Water Environ. Res. 2018, 90, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, M.A. Effects of flowing water on nitrogen-limited and phosphorus-limited photosynthesis and optimum N/P ratios by Spirogyra-fluviatilis (charophyceae). J. Phycol. 1994, 30, 418–430. [Google Scholar] [CrossRef]
- Fang, B.X.; Shi, J.Q.; Qin, L.; Feng, M.B.; Cheng, D.R.; Wang, T.T.; Zhang, X.S. Toxicity evaluation of 4,4′-di-CDPS and 4,4′-di-CDE on green algae Scenedesmus obliquus: Growth inhibition, change in pigment content, and oxidative stress. Environ. Sci. Pollut. Res. 2018, 25, 15630–15640. [Google Scholar] [CrossRef]
- Schoffelen, N.J.; Mohr, W.; Ferdelman, T.G.; Littmann, S.; Duerschlag, J.; Zubkov, M.V.; Ploug, H.; Kuypers, M.M.M. Single-cell imaging of phosphorus uptake shows that key harmful algae rely on different phosphorus sources for growth. Sci. Rep. 2018, 8, 17182. [Google Scholar] [CrossRef]
- Iummato, M.M.; Sabatini, S.E.; Cacciatore, L.C.; Cochon, A.C.; Cataldo, D.; de Molina, M.D.R.; Juarez, A.B. Biochemical responses of the golden mussel Limnoperna fortunei under dietary glyphosate exposure. Ecotoxicol. Environ. Saf. 2018, 163, 69–75. [Google Scholar] [CrossRef]
- Kooshki, H.; Ghollasi, M.; Halabian, R.; Kazemi, N.M. Osteogenic differentiation of preconditioned bone marrow mesenchymal stem cells with lipopolysaccharide on modified poly-l-lactic-acid nanofibers. J. Cell. Physiol. 2019, 234, 5343–5353. [Google Scholar] [CrossRef]
- Chaudhuri, D.; Ghate, N.B.; Deb, S.; Panja, S.; Sarkar, R.; Rout, J.; Mandal, N. Assessment of the phytochemical constituents and antioxidant activity of a bloom forming microalgae Euglena tuba. Biol. Res. 2014, 47, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, T.R.; Strickland, J.D.H. Discussion of spectrophotometric determination of marine-plant pigments, with revised equations for ascertaining chlorophylls and carotenoids. J. Mar. Res. 1963, 21, 155–163. [Google Scholar]
- Wei, F.S.; Qi, W.M.; Bi, T.; Sun, Z.G.; Huang, Y.R.; Shen, Y.W. Water and Wastewater Monitoring and Analysis Method, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 211–284. [Google Scholar]
- Ji, F.; Liu, Y.; Hao, R.; Li, G.; Zhou, Y.G.; Dong, R.J. Biomass production and nutrients removal by a new microalgae strain Desmodesmus sp. in anaerobic digestion wastewater. Bioresour. Technol. 2014, 161, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.G.; Li, A.J.; Qin, J.H.; Li, Q.; Ho, J.G.; Li, H.S. Seasonal patterns of water quality and phytoplankton dynamics in surface waters in Guangzhou and Foshan, China. Sci. Total Environ. 2017, 590, 361–369. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Chaudiere, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, 37, 949–962. [Google Scholar] [CrossRef]
- Farghl, A.M.; Shaddad, M.A.K.; Galal, H.R.; Hassan, E.A. Effect of Salt Stress on Growth, Antioxidant Enzymes, Lipid Peroxidation and Some Metabolic Activities in Some Fresh Water and Marine Algae. Egypt. J. Bot. 2015, 55, 1–15. [Google Scholar]
- Hong, Y.; Hu, H.Y.; Xie, X.; Sakoda, A.; Sagehashi, M.; Li, F.M. Gramine-induced growth inhibition, oxidative damage and antioxidant responses in freshwater cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2009, 91, 262–269. [Google Scholar] [CrossRef]
- Kumar, M.; Bijo, A.J.; Baghel, R.S.; Reddy, C.R.K.; Jha, B. Selenium and spermine alleviate cadmium induced toxicity in the red seaweed Gracilaria dura by regulating antioxidants and DNA methylation. Plant Physiol. Biochem. 2012, 51, 129–138. [Google Scholar] [CrossRef]
- Blum, J.; Fridovich, I. Inactivation of glutathione-peroxidase by superoxide radical. Arch. Biochem. Biophys. 1985, 240, 500–508. [Google Scholar] [CrossRef]
- Sinet, P.M.; Garber, P. Inactivation of the human CuZn superoxide-dismutase during exposure to O2− and H2O2. Arch. Biochem. Biophys. 1981, 212, 411–416. [Google Scholar] [CrossRef]
- Istvanovics, V.; Padisak, J.; Pettersson, K.; Pierson, D.C. Growth and phosphorus uptake of summer phytoplankton in lake Erken (Sweden). J. Plankton Res. 1994, 16, 1167–1196. [Google Scholar] [CrossRef] [Green Version]
- Ciugulea, I.; Triemer, R.E. Color atlas of photosynthetic euglenoids. J. Phycol. 2010, 42, 38. [Google Scholar]
- Gao, L.; Zhang, L.H.; Hou, J.Z.; Wei, Q.; Fu, F.; Shao, H.B. Decomposition of macroalgal blooms influences phosphorus release from the sediments and implications for coastal restoration in Swan Lake, Shandong, China. Ecol. Eng. 2013, 60, 19–28. [Google Scholar] [CrossRef]
- Wang, D.; Xie, L.L.; Zhu, X.B.; Bi, X.; Zheng, Y.Z.; Zhu, Y.K. Study on the metabolites of DH-e, a Halomonas marine bacterium, against three toxic dinoflagellate species. Water Sci. Technol. 2018, 78, 1535–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.Y.; Yang, Y.T.; Huang, M.J.; Zhou, C.H.; Lu, J.S. Photoelectrochemical determination of alkaline phosphatase activity based on a photo-excited electron transfer strategy. Talanta 2019, 196, 293–299. [Google Scholar] [CrossRef]
- Hoppe, H.G. Phosphatase activity in the sea. Hydrobiologia 2003, 493, 187–200. [Google Scholar] [CrossRef]
- Schaub, I.; Baum, C.; Schumann, R.; Karsten, U. Effects of an Early Successional Biological Soil Crust from a Temperate Coastal Sand Dune (NE Germany) on Soil Elemental Stoichiometry and Phosphatase Activity. Microb. Ecol. 2019, 77, 217–229. [Google Scholar] [CrossRef]
- Li, P.; Liu, W.H.; Gao, K.S. Effects of temperature, pH, and UV radiation on alkaline phosphatase activity in the terrestrial cyanobacterium Nostoc flagelliforme. J. Appl. Phycol. 2013, 25, 1031–1038. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Velocity (m/s) | Chla |
|---|---|
| 0 | 0.904 * |
| 0.1 | 0.822 * |
| 0.2 | 0.894 * |
| 0.3 | 0.931 ** |
| 0.4 | 0.878 * |
| 0.5 | 0.902 * |
| Flow Velocity (m/s) | Prediction Formula of Algal Growth | R2 |
|---|---|---|
| 0 | y = −0.0015x2 + 0.067x + 0.8971 | 0.9418 |
| 0.1 | y = −0.0025x2 + 0.0328x + 0.8614 | 0.9172 |
| 0.2 | y = 0.0004x2 – 0.0475x + 1.0554 | 0.8984 |
| 0.3 | y = 0.0007x2 – 0.055x + 1.0773 | 0.938 |
| 0.4 | y = 0.0008x2 – 0.0489x + 0.984 | 0.9238 |
| 0.5 | y = 0.0019x2 – 0.075x + 0.9193 | 0.9636 |
| Flow Velocity (m/s) | Biomass | SOD |
|---|---|---|
| 0 | −0.720 | −0.020 |
| 0.1 | −0.877 ** | 0.333 |
| 0.2 | −0.985 ** | 0.645 |
| 0.3 | −0.962 ** | 0.817 * |
| 0.4 | −0.955 ** | 0.737 |
| 0.5 | −0.978 ** | 0.904 ** |
| Flow Velocity (m/s) | Instantaneous Consumption Rate | Steady-state Consumption Rate | ||
|---|---|---|---|---|
| TN | TP | TN | TP | |
| 0 | 64.00 | 1.45 | 17.72 | 0.12 |
| 0.1 | 51.46 | 1.80 | 15.11 | 0.10 |
| 0.2 | 59.00 | 1.72 | 15.08 | 0.07 |
| 0.3 | 53.63 | 1.65 | 14.80 | 0.06 |
| 0.4 | 59.41 | 1.42 | 13.54 | 0.03 |
| 0.5 | 50.53 | 1.76 | 13.87 | −0.12 |
| Flow Velocity (m/s) | TP | Biomass | MDA | SOD |
|---|---|---|---|---|
| 0 | 0.386 | −0.617 | 0.44 | 0.412 |
| 0.1 | 0.323 | −0.894 ** | 0.804 * | 0.670 |
| 0.2 | 0.393 | −0.966 ** | 0.976 ** | 0.772 * |
| 0.3 | 0.460 | −0.975 ** | 0.988 ** | 0.858 * |
| 0.4 | 0.510 | −0.970 ** | 0.922 ** | 0.896 ** |
| 0.5 | 0.394 | −0.969 ** | 0.945 ** | 0.826 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Y.; Li, J.; Zhang, L.; Chen, M.; Zhang, Y.; An, R. Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs. Int. J. Environ. Res. Public Health 2019, 16, 4641. https://doi.org/10.3390/ijerph16234641
Tan Y, Li J, Zhang L, Chen M, Zhang Y, An R. Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs. International Journal of Environmental Research and Public Health. 2019; 16(23):4641. https://doi.org/10.3390/ijerph16234641
Chicago/Turabian StyleTan, Yi, Jia Li, Linglei Zhang, Min Chen, Yaowen Zhang, and Ruidong An. 2019. "Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs" International Journal of Environmental Research and Public Health 16, no. 23: 4641. https://doi.org/10.3390/ijerph16234641
APA StyleTan, Y., Li, J., Zhang, L., Chen, M., Zhang, Y., & An, R. (2019). Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs. International Journal of Environmental Research and Public Health, 16(23), 4641. https://doi.org/10.3390/ijerph16234641