The Role of Somatic Maturation on Bioimpedance Patterns and Body Composition in Male Elite Youth Soccer Players

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Body Composition
2.3. Bioelectric Impedance Vector Analysis
2.4. Somatic Maturation
2.5. Statistical Analysis
3. Results
3.1. Body Composition Parameters
3.2. BIVA Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abbott, A.; Button, C.; Pepping, G.J.; Collins, D. Unnatural selection: Talent identification and development in sport. Nonlinear Dyn. Psychol. Life Sci. 2005, 9, 61–88. [Google Scholar]
- Deprez, D.N.; Fransen, J.; Lenoir, M.; Philippaerts, R.M.; Vaeyens, R. A retrospective study on anthropometrical, physical fitness, and motor coordination characteristics that influence dropout, contract status, and first-team playing time in high-level soccer players aged eight to eighteen years. J. Strength Cond. Res. 2015, 29, 1692–1704. [Google Scholar] [CrossRef] [PubMed]
- Rommers, N.; Mostaert, M.; Goossens, L.; Vaeyens, R.; Witvrouw, E.; Lenoir, M.; D’Hondt, E. Age and maturity related differences in motor coordination among male elite youth soccer players. J. Sports Sci. 2019, 37, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Huertas, F.; Gines, H.J.; Hamidi, A.K.; Moratal, C.; Lupianez, J. Relative age effect in the sport environment. Role of physical fitness and cognitive function in youth soccer players. Int. J. Environ. Res. Public Health 2019, 16, 2837. [Google Scholar] [CrossRef]
- Lovell, R.; Towlson, C.; Parkin, G.; Portas, M.; Vaeyens, R.; Cobley, S. Soccer player characteristics in English lower-league development programmes: The relationships between relative age, maturation, anthropometry and physical fitness. PLoS ONE 2015, 10, e0137238. [Google Scholar] [CrossRef]
- Bezuglov, E.; Nikolaidis, P.T.; Khaitin, V.; Usmanova, E.; Luibushkina, A.; Repetiuk, A.; Waśkiewicz, Z.; Gerasimuk, D.; Rosemann, T.; Knechtle, B. Prevalence of relative age effect in russian soccer: The role of chronological age and performance. Int. J. Environ. Res. Public Health 2019, 16, 4055. [Google Scholar] [CrossRef] [PubMed]
- Campa, F.; Piras, A.; Raffi, M.; Toselli, S. Functional movement patterns and body composition of high-level volleyball, soccer, and rugby players. J. Sport Rehabil. 2019, 28, 740–745. [Google Scholar] [CrossRef]
- Campa, F.; Semprini, G.; Judice, P.B.; Messina, G.; Toselli, S. Anthropometry, physical and movement features, and repeated-sprint ability in soccer players. Int. J. Sports Med. 2019, 40, 100–109. [Google Scholar] [CrossRef]
- Leao, C.; Camões, M.; Clemente, F.M.; Nikolaidis, P.T.; Lima, R.; Bezerra, P.; Rosemann, T.; Knechtle, B. Anthropometric profile of soccer players as a determinant of position specificity and methodological issues of body composition estimation. Int. J. Environ. Res. Public Health 2019, 16, 2386. [Google Scholar] [CrossRef]
- Nalbant, O.; Ozer, K. Evaluation of the relationship between body composition and aerobic fitness in youth soccer players. Phys. Educ. Studens 2018, 22, 258–264. [Google Scholar] [CrossRef]
- Söğüt, M.; Clemente, F.M.; Clark, C.C.T.; Nikolaidis, P.T.; Rosemann, T.; Knechtle, B. Variations in central adiposity, cardiovascular fitness, and objectively measured physical activity according to weight status in children (9–11 years). Front. Physiol. 2019, 10, 936. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Campos, R.; Santi-Maria, T.; Arruda, M.; Maldonado, T.; Albernaz, A.; Schiavo, M.; Cossio-Bolaños, M. Fat-free mass and bone mineral density of young soccer players: Proposal of equations based on anthropometric variables. Front. Psychol. 2019, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Kyle, U.G.; Earthman, C.P.; Pichard, C.; Coss-Bu, J.A. Body composition during growth in children: Limitations and perspectives of bioelectrical impedance analysis. Eur. J. Clin. Nutr. 2015, 69, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Campa, F.; Gatterer, H.; Lukaski, H.; Toselli, S. Stabilizing bioimpedance-vector-analysis measures with a 10-minute cold shower after running exercise to enable assessment of body hydration. Int. J. Sports Physiol. Perform. 2019, 14, 1006–1009. [Google Scholar] [CrossRef]
- Campa, F.; Toselli, S. Bioimpedance vector analysis of elite, sub-elite and low-level male volleyball players. Int. J. Sports Physiol. Perform. 2018, 13, 1250–1253. [Google Scholar] [CrossRef]
- Carrasco-Marginet, M.; Castizo-Olier, J.; Rodríguez-Zamora, L.; Iglesias, X.; Rodríguez, F.A.; Chaverri, D.; Brotons, D.; Irurtia, A. Bioelectrical impedance vector analysis (BIVA) for measuring the hydration status in young elite synchronized swimmers. PLoS ONE 2017, 12, e0178819. [Google Scholar] [CrossRef]
- Castizo-Olier, J.; Irurtia, A.; Jemni, M.; Carrasco-Marginet, M.; Fernández-García, R.; Rodríguez, F.A. Bioelectrical impedance vector analysis (BIVA) in sport and exercise: Systematic review and future perspectives. PLoS ONE 2018, 13, e0197957. [Google Scholar] [CrossRef]
- Mascherini, G.; Gatterer, H.; Lukaski, H.; Burtscher, M.; Galanti, G. Changes in hydration, body-cell mass and endurance performance of professional soccer players through a competitive season. J. Sports Med. Phys. Fit. 2015, 55, 749–755. [Google Scholar]
- Piras, A.; Campa, F.; Toselli, S.; Di Michele, R.; Raffi, M. Physiological responses to partial-body cryotherapy performed during a concurrent strength and endurance session. Appl. Physiol. Nutr. Metab. 2019, 44, 59–65. [Google Scholar] [CrossRef]
- Piccoli, A.; Rossi, B.; Pillon, L.; Bucciante, G. A new method for monitoring body fluid variation by bioimpedance analysis: The RXc graph. Kidney Int. 1994, 46, 534–539. [Google Scholar] [CrossRef]
- Campa, F.; Matias, C.N.; Marini, E.; Heymsfield, S.B.; Toselli, S.; Sardinha, L.B.; Silva, A.M. Identifying athlete body-fluid changes during a competitive season with bioelectrical impedance vector analysis. Int. J. Sports Physiol. Perform. 2019. [Google Scholar] [CrossRef] [PubMed]
- Marini, E.; Campa, F.; Buffa, R.; Stagi, S.; Matias, C.N.; Toselli, S.; Sardinha, L.B.; Silva, A.M. Phase angle and bioelectrical impedance vector analysis in the evaluation of body composition in athletes. Clin. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed]
- Koury, J.C.; Trugo, N.M.F.; Torres, A.G. Phase angle and bioelectrical impedance vectors in adolescent and adult male athletes. Int. J. Sports Physiol. Perform. 2014, 9, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Koury, J.C.; de Oliveira-Junior, A.V.; Portugal, M.R.C.; de Oliveira, K.J.F.; Donangelo, C.M. Bioimpedance parameters in adolescent athletes in relation to bone maturity and biochemical zinc indices. J. Trace Elem. Med. Biol. 2018, 46, 26–31. [Google Scholar] [CrossRef]
- Towlson, C.; Cobley, S.; Parkin, G.; Lovell, R. When does the influence of maturation on anthropometric and physical fitness characteristics increase and subside? Scand. J. Med. Sci. Sports 2018, 28, 1946–1955. [Google Scholar] [CrossRef]
- Frisancho, A.R. Anthropometric Standards: An Interactive Nutritional Reference of Body Size and Body Composition for Children and Adults; University of Michigan Press: Ann Arbor, MI, USA, 2008. [Google Scholar]
- Lukaski, H.; Piccoli, A. Bioelectrical impedance vector analysis for assessment of hydration in physiological states and clinical conditions. In Handbook of Anthropometry; Preedy, V., Ed.; Springer: London, UK, 2012; pp. 287–305. [Google Scholar]
- Micheli, M.L.; Pagani, L.; Marella, M.; Gulisano, M.; Piccoli, A.; Angelini, F.; Burtscher, M.; Gatterer, H. Bioimpedance and impedance vector patterns as predictors of league level in male soccer players. Int. J. Sports Physiol. Perform. 2014, 9, 532–539. [Google Scholar] [CrossRef]
- Mirwald, R.L.; Baxter-Jones, A.D.; Bailey, D.A.; Beunen, G.P. An assessment of maturity from anthropometric measurements. Med. Sci. Sports Exerc. 2002, 34, 689–694. [Google Scholar]
- Toselli, S.; Merni, F.; Campa, F. Height prediction in elite Italian rugby players: A prospective study. Am. J. Hum. Biol. 2019, 31, e23288. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Modeling temperature effects on mortality: Multiple segmented relationships with common break points. Biostatistics 2008, 9, 613–620. [Google Scholar] [CrossRef]
- Malina, R.M.O.; Bar-Or, O.; Bouchard, C. Growth, Maturation, and Physical Activity; Human Kinetics: Champaign, IL, USA, 2004. [Google Scholar]
- Kang, M.J.; Hong, H.S.; Chung, S.J.; Lee, Y.A.; Shin, C.H.; Yang, S.W. Body composition and bone density reference data for Korean children, adolescents, and young adults according to age and sex: Results of the 2009–2010 Korean National Health and Nutrition Examination Survey (KNHANES). J. Bone Miner. Metab. 2016, 34, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Agostinete, R.R.; Ito, I.H.; Kemper, H.; Pastre, C.M.; Rodrigues-Júnior, M.A.; Luiz-de-Marco, R.; Fernandes, R.A. Somatic maturation and the relationship between bone mineral variables and types of sports among adolescents: Cross-sectional study. Sao Paulo Med. J. 2017, 135, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Campa, F.; Silva, A.M.; Toselli, S. Changes in phase angle and handgrip strength induced by suspension training in older women. Int. J. Sports Med. 2018, 39, 442–449. [Google Scholar] [CrossRef] [PubMed]
- de Palo, T.; Messina, G.; Edefonti, A.; Perfumo, F.; Pisanello, L.; Peruzzi, L.; Di Iorio, B.; Mignozzi, M.; Vienna, A.; Conti, G.; et al. Normal values of the bioelectrical impedance vector in childhood and puberty. Nutrition 2000, 16, 417–424. [Google Scholar] [CrossRef]
- Toselli, S.; Campa, F. Anthropometry and functional movement patterns in elite male volleyball players of different competitive levels. J. Strength Cond. Res. 2018, 32, 2601–2611. [Google Scholar] [CrossRef]
- Malina, R.M.; Coelho, E.; Silva, M.J.; Figueiredo, A.J.; Carling, C.; Beunen, G.P. Interrelationships among invasive and non-invasive indicators of biological maturation in adolescent male soccer players. J. Sports Sci. 2012, 30, 1705–1717. [Google Scholar] [CrossRef]
- Backous, D.D.; Farrow, J.A.; Friedl, K.E. Assessment of pubertal maturity in boys, using height and grip strength. J. Adolesc. Health Care 1990, 11, 497–500. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | U10 N = 26 | U11 N = 26 | U12 N = 26 | U13 N = 41 | U14 N = 31 | U15 N = 28 | U16 N = 29 | U19 N = 35 |
|---|---|---|---|---|---|---|---|---|
| Age (y) | 9.9 ± 0.2 | 10.8 ± 0.3 | 11.7 ± 0.3 | 12.8 ± 0.3 | 13.9 ± 0.2 | 14.9 ± 0.2 | 15.5 ± 0.5 | 17.7 ± 0.5 |
| Weight (kg) | 31.6 ± 4.0 | 31.6 ± 4.6 | 35.9 ± 5.2 | 39.7 ± 7.4 | 48.0 ± 7.5 | 55.4 ± 5.8 | 63.8 ± 7.1 | 67.7 ± 6.0 |
| Height (cm) | 131.6 ± 6.2 | 144.5 ± 5.3 | 151.1 ± 7.5 | 152.8 ± 11.1 | 167.7 ± 8.0 | 174.0 ± 7.2 | 176.1 ± 5.9 | 177.4 ± 5.8 |
| BMI (kg/m2) | 16.9 ± 1.4 | 17.1 ± 1.5 | 18.7 ± 1.7 | 19.3 ± 2.1 | 20.7 ± 1.9 | 21.1 ± 1.0 | 21.8 ± 1.8 | 22.0 ± 1.3 |
| Sitting height (cm) | 61.1 ± 2.9 | 71.5 ± 2.9 | 73.3 ± 3.9 | 77.9 ± 3.5 | 83.7 ± 4.1 | 88.3 ± 3.5 | 93.2 ± 2.5 | 88.9 ± 3.6 |
| Leg length (cm) | 70.5 ± 5.0 | 73.0 ± 4.0 | 77.8 ± 13.8 | 74.9 ± 10.6 | 84.0 ± 4.5 | 85.7 ± 4.7 | 82.9 ± 4.5 | 88.5 ± 5.6 |
| YPHV (y) | −3.8 ± 0.3 | −3.0 ± 0.3 | −2.4 ± 0.5 | −1.3 ± 0.5 | −0.1 ± 0.6 | 1.0 ± 0.5 | 2.0 ± 0.5 | 2.5 ± 0.5 |
| FM (%) | 12.8 ± 2.2 | 14.0 ± 3.2 | 15.9 ± 3.9 | 13.9 ± 4.0 | 12.3 ± 3.0 | 10.9 ± 1.9 | 13.3 ± 2.5 | 14.7 ± 2.5 |
| FFM (kg) | 27.6 ± 3.1 | 30.8 ± 3.5 | 33.3 ± 4.0 | 41.3 ± 5.9 | 48.5 ± 6.4 | 58.0 ± 5.8 | 58.7 ± 5.8 | 59.0 ± 4.2 |
| UMA (cm2) | 20.6 ± 3.5 | 24.6 ± 4.7 | 25.7 ± 5.0 | 29.3 ± 5.6 | 36.3 ± 6.1 | 45.8 ± 4.7 | 45.9 ± 7.5 | 44.3 ± 6.4 |
| UFA (cm2) | 7.6 ± 1.8 | 8.6 ± 2.5 | 9.8 ± 2.8 | 8.8 ± 3.3 | 8.5 ± 2.8 | 7.8 ± 1.9 | 11.1 ± 2.9 | 11.5 ± 3.5 |
| CMA (cm2) | 44.8 ± 5.4 | 48.0 ± 8.6 | 54.2 ± 19.1 | 60.8 ± 9.3 | 69.1 ± 13.4 | 65.7 ± 6.9 | 81.6 ± 12.6 | 74.6 ± 11.2 |
| CFA (cm2) | 18.5 ± 5.7 | 20.5 ± 6.6 | 24.8 ± 7.1 | 23.2 ± 7.5 | 22.9 ± 7.1 | 19.6 ± 3.4 | 24.4 ± 5.9 | 27.1 ± 5.6 |
| TMA (cm2) | 99.7 ± 12.5 | 109.8 ± 13.5 | 119.9 ± 25.7 | 132.9 ± 21.8 | 156.6 ± 27.9 | 183.7 ± 14.2 | 183.5 ± 22.2 | 195.4 ± 20.8 |
| TFA (cm2) | 19.4 ± 4.3 | 22.3 ± 6.3 | 23.8 ± 8.8 | 23.1 ± 7.7 | 22.6 ± 6.1 | 21.3 ± 4.2 | 24.5 ± 7.2 | 27.7 ± 9.0 |
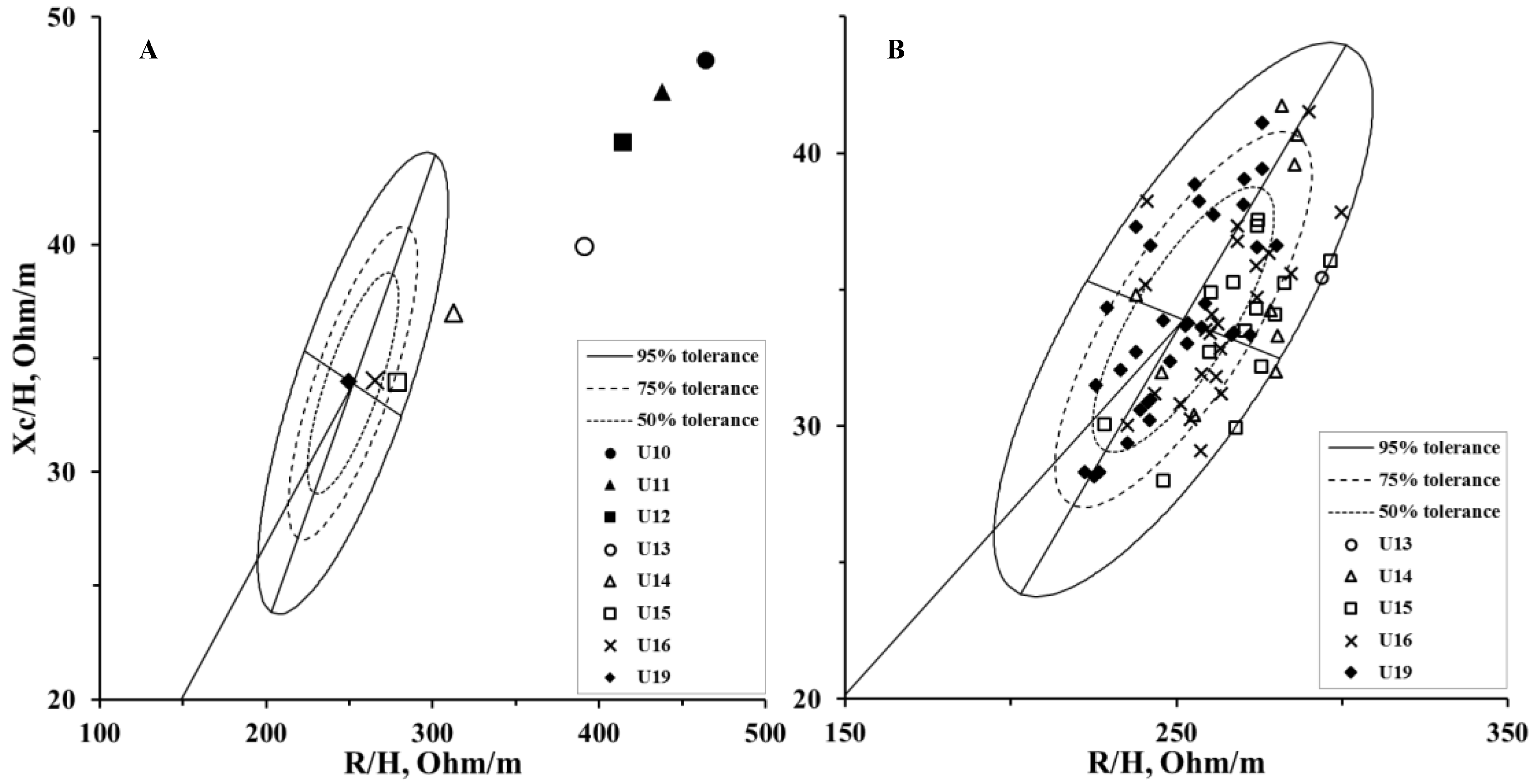
| R/H (ohm/m) | 465.4 ± 53.0 | 440.8 ± 42.7 | 416.2 ± 57.4 | 392.9 ± 55.9 | 313.5 ± 39.7 | 276.0 ± 19.4 | 265.4 ± 17.6 | 249.7 ± 20.2 |
| Xc/H (ohm/m) | 48.3 ± 5.7 | 46.8 ± 5.6 | 44.7 ± 6.9 | 40.1 ± 5.3 | 37.1 ± 3.9 | 33.8 ± 3.0 | 34.1 ± 3.1 | 34.0 ± 3.8 |
| PA (°) | 6.0 ± 0.6 | 6.1 ± 0.5 | 6.2 ± 0.5 | 5.9 ± 0.5 | 6.8 ± 0.8 | 7.0 ± 0.5 | 7.4 ± 0.6 | 7.8 ± 0.6 |
| VL/H (ohm/m) | 467.9 ± 53.1 | 443.3 ± 42.9 | 418.6 ± 57.7 | 395.0 ± 56.1 | 315.7 ± 39.7 | 278.1 ± 19.5 | 267.6 ± 17.6 | 252.0 ± 20.4 |
| TBW (l) | 20.9 ± 2.1 | 22.9 ± 3.2 | 25.1 ± 4.1 | 30.9 ± 3.0 | 35.5 ± 3.6 | 42.4 ± 3.6 | 43.0 ± 2.7 | 42.8 ± 3.2 |
| BCM (kg) | 21.9 ± 1.4 | 22.4 ± 1.8 | 23.5 ± 1.8 | 23.9 ± 2.2 | 28.7 ± 4.1 | 31.7 ± 2.0 | 33.4 ± 2.0 | 35.7 ± 2.4 |
| Breakpoints in Development (Years) | Rate of Player Development | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Breakpoint 1. (95% CI) | Breakpoint 2. (95% CI) | Slope 1 (95% CI) | p-Value | Slope 2 (95% CI) | p-Value | Slope 3 (95% CI) | p-Value | r2 |
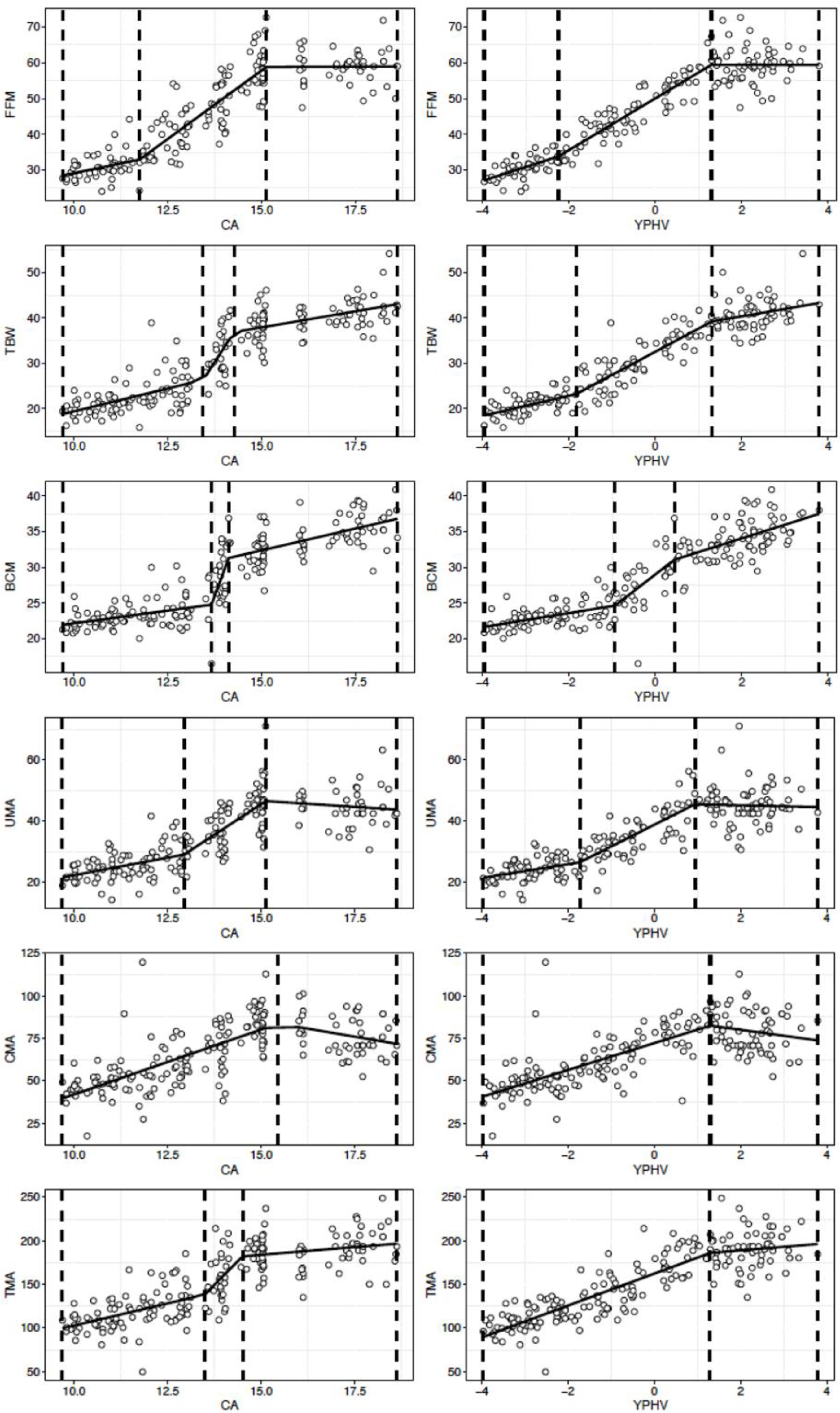
| FFM | 11.7 (11.1; 12.3) | 15.1 (14.5; 15.7) | 2.1 (−0.4; 4.7) | 0.056 | 7.6 (6.7; 8.5) | <0.001 | 0.1 (−1.7; 1.8) | 0.48 | 0.84 |
| TBW | 13.4 (12.9; 13.9) | 14.2 (13.8; 14.6) | 2.1 (1.3; 2.8) | <0.001 | 12.7 (5.0; 20.4) | 0.01 | 1.4 (0.8; 2.0) | <0.001 | 0.84 |
| BCM | 13.6 (13.2; 14.0) | 14.1 (13.8; 14.4) | 0.7 (0.2; 1.1) | <0.001 | 14.1 (3.6; 24.6) | <0.01 | 1.2 (0.8; 1.6) | <0.001 | 0.83 |
| UMA | 12.9 (12.2; 13.5) | 15.1 (14.4; 15.8) | 2.2 (0.9; 3.6) | <0.001 | 7.9 (6.0; 9.8) | <0.001 | −0.8 (−2.8; 1.2) | 0.22 | 0.72 |
| CMA | 15.4 (14.6; 16.2) | - | 7.6 (6.5; 8.8) | <0.001 | −3.7 (−8.0; 0.5) | 0.04 | - | - | 0.53 |
| TMA | 13.5 (12.7; 14.2) | 14.5 (13.6; 15.3) | 10.1 (5.8; 14.4) | <0.001 | 43.3 (4.9; 81.6) | 0.01 | 3.5 (0.1; 7.0) | 0.02 | 0.72 |
| VL/H | 12.8 (12.1; 13.5) | 14.3 (13.9; 14.7) | −19.4 (−29.5; −9.3) | <0.001 | −76.7 (−103.5; −49.9) | <0.001 | −9.0 (−15.6; −2.5) | <0.01 | 0.79 |
| PA | 12.7 (11.9; 13.4) | 16.9 (16.3; 17.5) | −0.1 (−0.2; 0.2) | 0.48 | 0.4 (0.3; 0.5) | <0.001 | −0.4 (−0.8; 0.0) | 0.055 | 0.58 |
| Breakpoints in Development (Years) | Rate of Player Development | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Breakpoint 1. (95% CI) | Breakpoint 2. (95% CI) | Slope 1 (95% CI) | p-Value | Slope 2 (95% CI) | p-Value | Slope 3 (95% CI) | p-Value | r2 |
| FFM | −2.2 (−2.8; −1.6) | 1.3 (1.0; 1.6) | 3.9 (1.7; 6.0) | <0.001 | 7.2 (6.4; 8.0) | <0.001 | −0.1 (−1.5; 1.5) | 0.50 | 0.91 |
| TBW | −1.8 (−2.4; −1.2) | 1.3 (0.6; 2.0) | 2.2 (1.0; 3.4) | <0.001 | 5.1 (4.3; 5.9) | <0.001 | 1.6 (0.4; 2.8) | <0.01 | 0.91 |
| BCM | −0.9 (−1.6; −0.2) | 0.4 (−0.5;1.3) | 1.0 (0.5; 1.5) | <0.001 | 4.7 (2.9; 6.5) | <0.001 | 1.9 (1.3; 2.5) | <0.001 | 0.85 |
| UMA | −1.7 (−2.4; −1.0) | 0.9 (0.3; 1.5) | 2.2 (0.1; 4.4) | 0.02 | 7.1 (5.4; 8.8) | <0.001 | −0.3 (−2.3; 1.7) | 0.37 | 0.76 |
| CMA | 1.3 (0.7; 1.9) | - | 7.9 (6.6; 9.2) | <0.001 | −3.5 (−8.5; 1.5) | 0.08 | - | - | 0.56 |
| TMA | 1.3 (0.5; 2.1) | - | 18.3 (16.1; 20.5) | <0.001 | 4.1 (−3.7; 12.0) | 0.15 | - | - | 0.78 |
| VL/H | 1.1 (0.6; 1.6) | - | −42.8 (−47.0; −38.6) | <0.001 | −9.6 (−23.4; 4.2) | 0.09 | - | - | 0.83 |
| PA | −1.5 (−2.3; −0.7) | - | −0.0 (−0.2; 0.2) | 0.44 | 0.4 (0.3; 0.5) | <0.001 | - | - | 0.56 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campa, F.; Silva, A.M.; Iannuzzi, V.; Mascherini, G.; Benedetti, L.; Toselli, S. The Role of Somatic Maturation on Bioimpedance Patterns and Body Composition in Male Elite Youth Soccer Players. Int. J. Environ. Res. Public Health 2019, 16, 4711. https://doi.org/10.3390/ijerph16234711
Campa F, Silva AM, Iannuzzi V, Mascherini G, Benedetti L, Toselli S. The Role of Somatic Maturation on Bioimpedance Patterns and Body Composition in Male Elite Youth Soccer Players. International Journal of Environmental Research and Public Health. 2019; 16(23):4711. https://doi.org/10.3390/ijerph16234711
Chicago/Turabian StyleCampa, Francesco, Analiza Monica Silva, Vincenzo Iannuzzi, Gabriele Mascherini, Luca Benedetti, and Stefania Toselli. 2019. "The Role of Somatic Maturation on Bioimpedance Patterns and Body Composition in Male Elite Youth Soccer Players" International Journal of Environmental Research and Public Health 16, no. 23: 4711. https://doi.org/10.3390/ijerph16234711
APA StyleCampa, F., Silva, A. M., Iannuzzi, V., Mascherini, G., Benedetti, L., & Toselli, S. (2019). The Role of Somatic Maturation on Bioimpedance Patterns and Body Composition in Male Elite Youth Soccer Players. International Journal of Environmental Research and Public Health, 16(23), 4711. https://doi.org/10.3390/ijerph16234711