The Effects of Cadmium Toxicity

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Chemical Form and Properties of Cadmium
3. Exposure to Cadmium and Toxicity
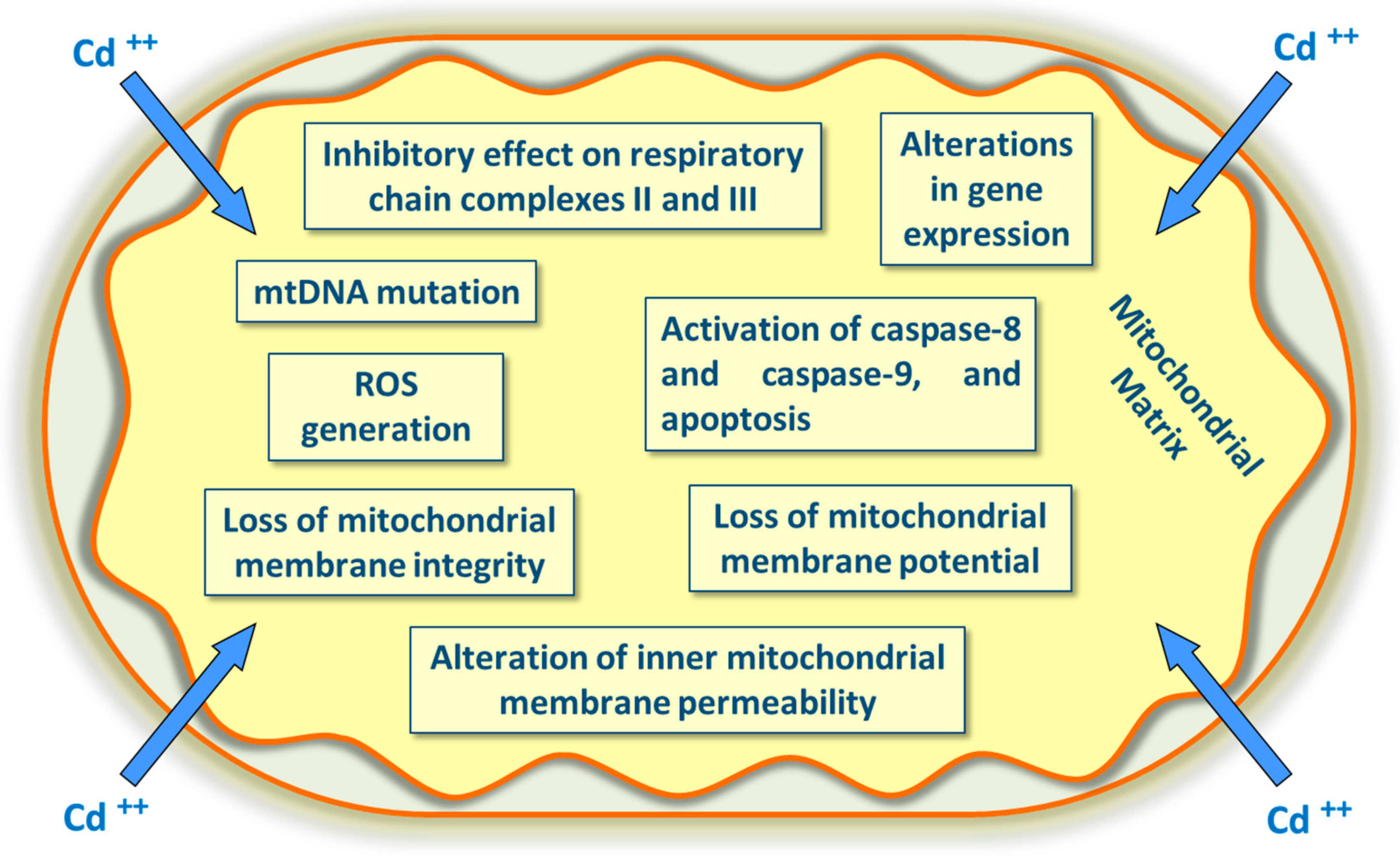
4. Cadmium’s Effects on Mitochondria and Cd2+-Induced Apoptosis
5. Cadmium and Metallothioneins
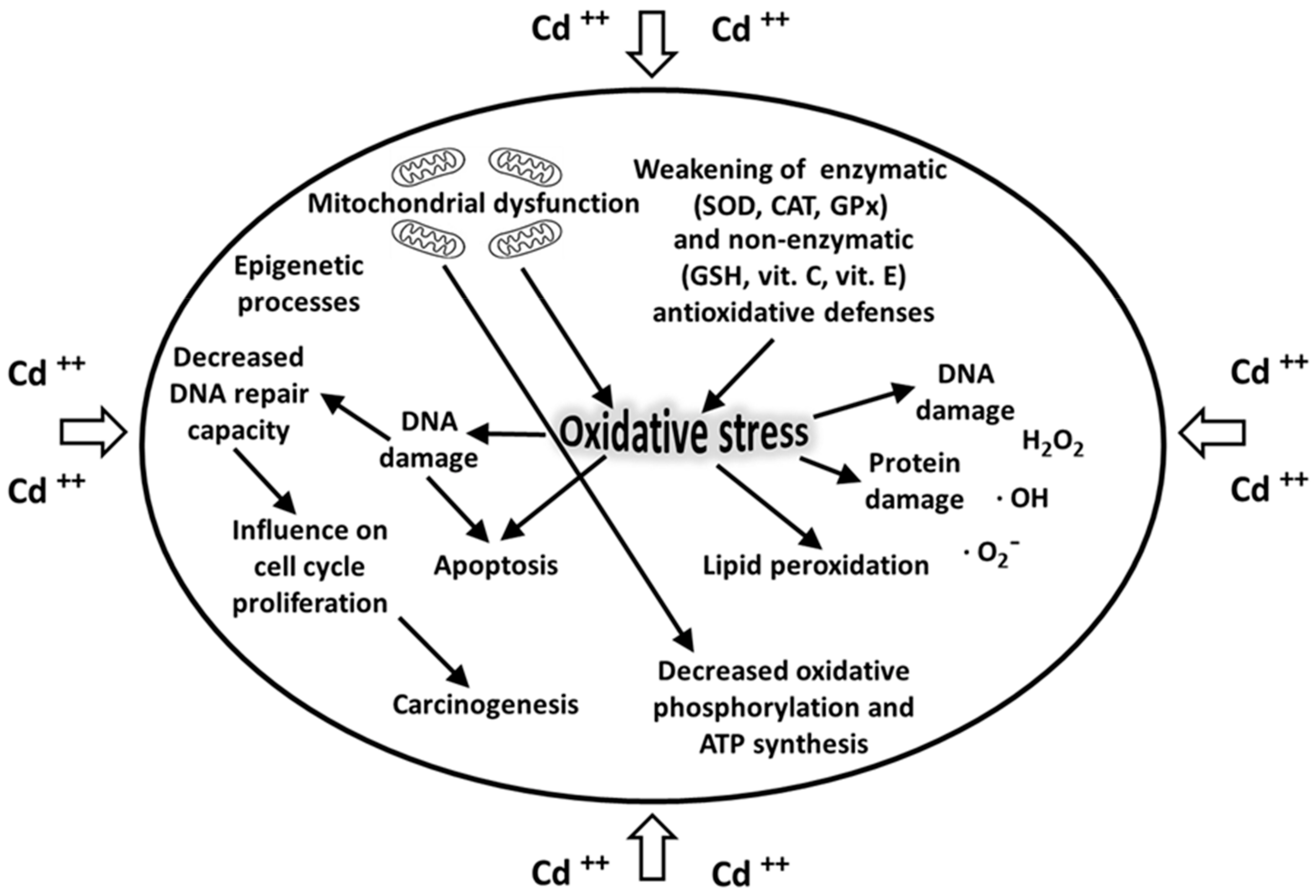
6. Role of Oxidative Stress in Cadmium Toxicity
7. The Epigenetic Effects of Cadmium Exposure
8. Cadmium Carcinogenesis
9. Phytoremediation, Nanoparticles and Microbial Fermentation against Cadmium Toxicity
10. Conclusions
11. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sinicropi, M.S.; Amantea, D.; Caruso, A.; Saturnino, C. Chemical and biological properties of toxic metals and use of chelating agents for the pharmacological treatment of metal poisoning. Arch. Toxicol. 2010, 84, 501–520. [Google Scholar] [CrossRef] [PubMed]
- Sinicropi, M.S.; Caruso, A.; Capasso, A.; Palladino, C.; Panno, A.; Saturnino, C. Heavy metals: Toxicity and carcinogenicity. Pharmacologyonline 2010, 2, 329–333. [Google Scholar]
- Carocci, A.; Rovito, N.; Sinicropi, M.S.; Genchi, G. Mercury toxicity and neurodegenerative effects. Rev. Environ. Contam. Toxicol. 2014, 229, 1–18. [Google Scholar]
- Carocci, A.; Catalano, A.; Lauria, G.; Sinicropi, M.S.; Genchi, G. Lead Toxicity, antioxidant defense and environment. Rev. Environ. Contam. Toxicol. 2016, 238, 45–67. [Google Scholar]
- Genchi, G.; Sinicropi, M.S.; Carocci, A.; Lauria, G.; Catalano, A. Mercury exposure and heart diseases. Int. Environ. Res. Public Health 2017, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genchi, G.; Sinicropi, M.S.; Carocci, A.; Lauria, G.; Catalano, A. Response to comment on Giuseppe Genchi et al. Mercury exposure and heart diseases. Int. Environ. Res. Public Health 2017, 14, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friberg, L.T.; Elinder, G.G.; Kjellstrom, T.; Nordberg, G.F. (Eds.) Cadmium and Health: A Toxicological and Epidemiological Appraisal: Volume 2: Effects and Response (Vol. 1); CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Rahimzadeh, M.R.; Rahimzadeh, M.R.; Kazemi, S.; Moghadamnia, A.A. Cadmium toxicity and treatment: An update. Caspian J. Intern. Med. 2017, 8, 135–145. [Google Scholar]
- Genchi, G.; Carocci, A.; Lauria, G.; Sinicropi, M.S.; Catalano, A. Nickel: Human health and environmental toxicology. Int. Environ. Res. Public Health 2020, 17, 679. [Google Scholar] [CrossRef] [Green Version]
- Hogervost, J.; Plusquin, M.; Vangronsvel, J.; Nawrot, T.; Cuypers, A.; Van Hecke, E.; Roels, H.A.; Carleer, R.; Staessen, J.A. House dust as possible route of environmental exposure to cadmium and lead in the adult general population. Environ. Res. 2007, 103, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Casado, M.; Anawar, H.M.; Garcia-Sanchez, A.; Santa Regina, I. Cadmium and zinc in polluted mining soils and uptake by plants (El Losar mine, Spain). Int. Environ. Pollut. 2008, 33, 146–159. [Google Scholar] [CrossRef]
- Satarug, S. Dietary Cadmium intake and its effects on kidneys. Toxics 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinkov, A.A.; Gritsenko, V.A.; Skalnaya, M.G.; Cherkasov, S.V.; Aaseth, J.; Skalny, A.V. Gut as a target for cadmium toxicity. Environ. Pollut. 2018, 235, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Filippini, T.; Ajsuvakovae, O.P.; Skalnaya, M.G.; Aasethf, J.; Bjørklundh, G.; Gatiatulinai, E.R.; Popova, E.V.; Nemereshinai, O.N.; Huangk, P.T.; et al. Cadmium and atherosclerosis: A review of toxicological mechanisms and a meta-analysis of epidemiologic studies. Environ. Res. 2018, 162, 240–260. [Google Scholar] [CrossRef] [PubMed]
- IARC (International Agency for Research on Cancer). Monographs on the Evaluation of the Carcinogenic Risks to Humans Beryllium, Cadmium, Mercury and Exposures in the Glass Manufacturing Industry; IARC Scientific Publications: Lyon, France, 1993; pp. 119–238. [Google Scholar]
- IARC. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans-Beryllium, Cadmium, Mercury, and Exposures in the Glass Manufacturing Industry. In Summary of Data Reported and Evaluation; International Agency for Cancer Research-World Health Organization: Geneva, Switzerland, 1997; Volume 58. [Google Scholar]
- Mezynska, M.; Brzóska, M.M. Environmental exposure to cadmium—A risk for health of the general population in industrialized countries and preventive strategies. Environ. Sci. Pollut. Res. 2018, 25, 3211–3232. [Google Scholar] [CrossRef] [PubMed]
- Tamás, M.J.; Fauvet, B.; Christen, P.; Goloubinoff, P. Misfolding and aggregation of nascent proteins: A novel mode of toxic cadmium action in vivo. Curr. Genet. 2018, 64, 177–181. [Google Scholar] [CrossRef]
- Saturnino, C.; Iacopetta, D.; Sinicropi, M.S.; Rosano, C.; Caruso, A.; Caporale, A.; Marra, N.; Marengo, B.; Pronzato, M.A.; Parisi, O.I.; et al. N-Alkyl carbazole derivatives as new Tools for Alzheimer’s disease: Preliminary Studies. Molecules 2014, 19, 9307–9317. [Google Scholar] [CrossRef]
- Buha, A.; Jugdaohsingh, R.; Matovic, V.; Bulat, Z.; Antonijevic, B.; Kerns, J.G.; Goodship, A.; Hart, A.; Powell, J.J. Bone mineral health is sensitively related to environmental cadmium exposure-experimental and human data. Environ. Res. 2019, 176, 108539. [Google Scholar] [CrossRef]
- Jacobo-Estrada, T.; Cardenas-Gonzalez, M.; Santoyo-Sánchez, M.P.; Thevenod, F.; Barbier, O. Intrauterine exposure to cadmium reduces HIF-1 DNA-binding ability in rat fetal kidneys. Toxics 2018, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Cai, L.; Liu, Y.; Chen, W.; Wan, Q. Association between prenatal cadmium exposure and cognitive development of offspring: A systematic review. Environ. Pollut. 2019, 254, 113081. [Google Scholar] [CrossRef]
- Han, Y.L.; Sheng, Z.; Liu, G.D.; Long, L.L.; Wang, Y.F.; Yang, W.X.; Zhu, J.Q. Cloning, characterization and cadmium inducibility of metallothionein in the testes of the mudskipper Boleophthalmus pectinirostris. Ecotoxicol. Environ. Saf. 2015, 119, 1–8. [Google Scholar] [CrossRef]
- Prabu, S.M.; Shagirtha, K. Cadmium and Apoptosis: A Molecular Approach. Res. Rev. J. Toxicol. 2019, 6, 8–17. [Google Scholar]
- WHO World Health Organization. Environmental Health Criteria 134: Cadmium; World Health Organization: Geneva, Switzerland, 1992. [Google Scholar]
- Thompson, J.; Bannigan, J. Cadmium: Toxic effects on the reproductive system and the embryo. Reproduct. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M.T.; Nauman, M.; Nazir, N.; Ali, S.; Bangash, N. Environmental hazards of cadmium: Past, present, and future. In Cadmium Toxicity and Tolerance in Plants; Academic Press: Cambridge, MA, USA, 2019; pp. 163–183. [Google Scholar]
- Karunakaran, C.; Dhanalakshmi, R. Selectivity in photocatalysis by particulate semiconductors. Cent. Eur. Chem. 2009, 7, 134–137. [Google Scholar] [CrossRef]
- Lokhande, B.; Patil, P.S.; Uplane, M.D. Studies on cadmium oxide sprayed thin films deposited through non-aqueous medium. Mater. Chem. Phys. 2004, 84, 238–242. [Google Scholar] [CrossRef]
- Alterio, V.; Langella, E.; De Simone, G.; Monti, S.M. Cadmium-containing carbonic anhydrase CDCA1 in marine diatom Thalassiosira weissflogii. Mar. Drugs 2015, 13, 1688–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satarug, S. Cadmium sources and toxicity. Toxics 2019, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirot, V.; Samieri, C.; Volatier, L.; Leblanc, C. Cadmium dietary intake and biomarker data in French high seafood consumers. Expo. Sci. Environ. Epidemiol. 2008, 18, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Carey, M.; Meharg, C.; Williams, P.N.; Signes-Pastor, A.J.; Triwardhani, E.A.; Pandiangan, F.I.; Campbell, K.; Elliott, C.; Marwa, E.M.; et al. Rice grain cadmium concentrations in the global supply-chain. Expo. Health 2020, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Arao, T. Mitigation strategies for cadmium and arsenic in rice. In Cadmium Toxicity; Springer: Singapore, 2019; pp. 125–138. [Google Scholar]
- Horiguchi, H. Cadmium exposure and its effects on the health status of rice farmers in Akita prefecture. In Cadmium Toxicity; Springer: Singapore, 2019; pp. 75–83. [Google Scholar]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 104388. [Google Scholar] [CrossRef]
- Järup, L.; Akesson, A. Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef]
- ATSDR. Toxicological Profile for Cadmium. In Agency for Toxic Substances and Disease Registry; Public Health Service U.S. Department of Health and Human Services: Atlanta, GA, USA, 2012; pp. 1–487. [Google Scholar]
- Mona, T.; Heba, M.A.; Eman, S.; Khadiga, S.I.; Safaa, E. Impact of occupational cadmium exposure on bone in sewage workers. Int. J. Occup. Environ. Health 2018, 24, 101–108. [Google Scholar]
- Bulmer, F.M.R.; Rothwel, H.E.; Frankish, E.R. Industrial cadmium poisoning. Can. Public Health 1938, 29, 19–26. [Google Scholar]
- Hagino, N.; Yoshioka, Y. A study of the etiology of Itai-Itai disease. J. Jpn. Orthop. Assoc. 1960, 35, 812–815. [Google Scholar]
- Kumar, S.; Sharma, A. Cadmium toxicity: Effects on human reproduction and fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.X.; Wang, L. Cadmium: Toxic effects on placental and embryonic development. Environ. Toxicol. Pharmacol. 2019, 67, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Kippler, M.; Ekström, E.C.; Lönnerdal, B.; Goessler, W.; Akesson, A.; El Arifeen, S.; Persson, L.A.; Vahter, M. Influence of iron and zinc status on cadmium accumulation in Bangladeshi women. Toxicol. Appl. Pharmacol. 2007, 222, 221–226. [Google Scholar] [CrossRef]
- Söderholm, M.; Borné, Y.; Hedblad, B.; Persson, M.; Barregard, L.; Engström, G. Blood cadmium concentration and risk of subarachnoid haemorrhage. Environ. Res. 2020, 180, 108826. [Google Scholar] [CrossRef]
- Fatima, G.; Raza, A.M.; Hadi, N.; Nigam, N.; Mahd, A.A. Cadmium in human diseases: it’s more than just a mere metal. Ind. J. Clin. Biochem. 2019, 34, 371–378. [Google Scholar] [CrossRef]
- Reyes-Hinojosa, D.; Lozada-Pérez, C.A.; Zamudio Cuevas, Y.; López-Reyes, A.; Martínez-Nava, G.; Fernández-Torres, J.; Olivos-Meza, A.; Landa-Solis, C.; Gutiérrez-Ruiz, M.C.; Rojas del Castillo, E.; et al. Toxicity of cadmium in musculoskeletal diseases. Environ. Toxicol. Pharmacol. 2019, 72, 103219. [Google Scholar] [CrossRef]
- Bernhoft, R.A. Cadmium toxicity and treatment. Sci. World J. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
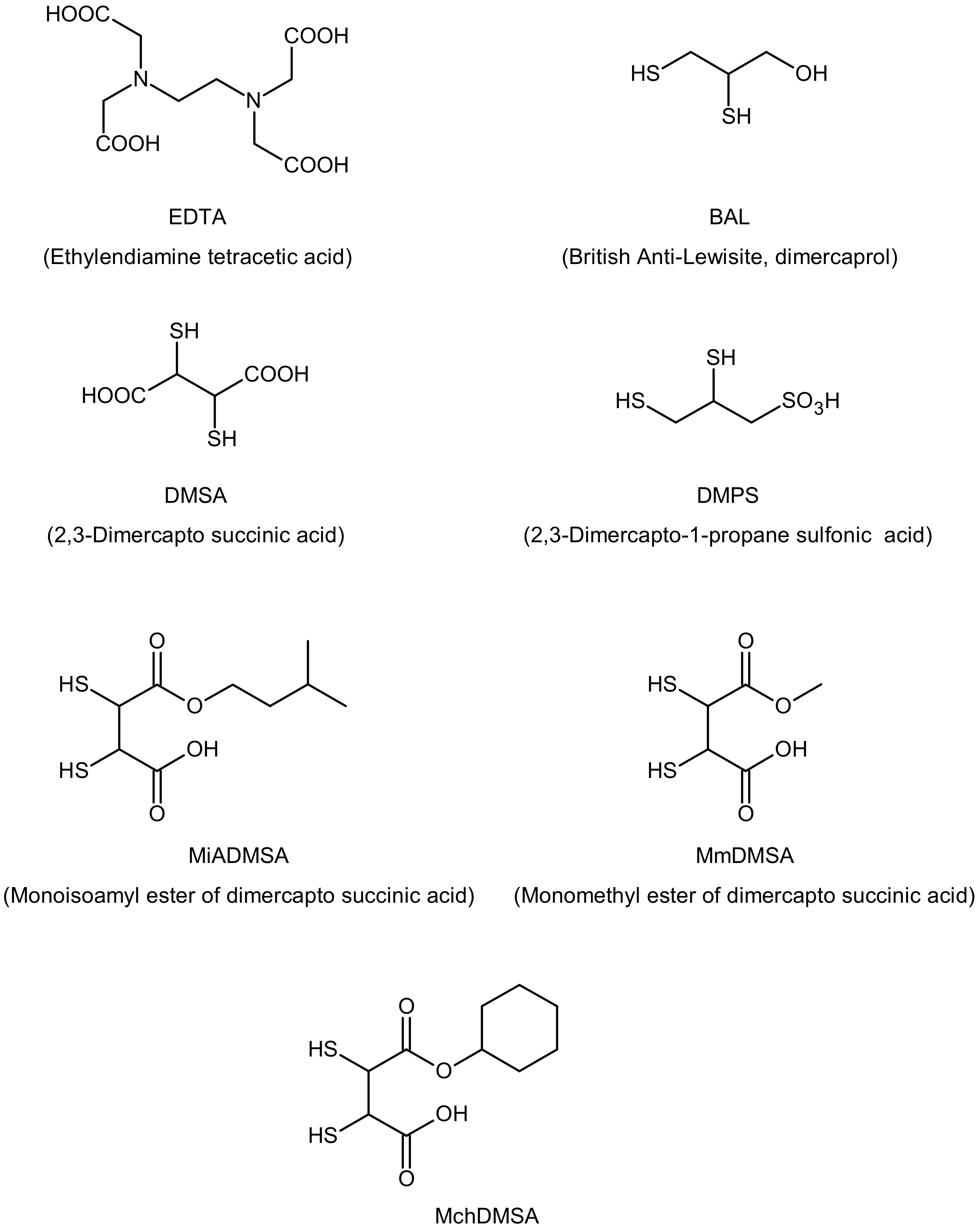
- Flora, S.S.; Pachauri, V. Chelation in metal intoxication. Int. Environ. Res. Public Health 2010, 7, 2745–2788. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Pant, S.C.; Flora, S. Monoisoamyl dimercaptosuccinic acid induced changes in pregnant female rats during late gestation and lactation. Reprod. Toxicol. 2006, 21, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Tandon, S.K.; Prasad, S.; Singh, S. Chelation in metal intoxication: Influence of cysteine or N-acetyl cysteine on the efficacy of 2,3-dimercaptopropane-1-sulphonate in the treatment of cadmium toxicity. Appl. Toxicol. 2002, 22, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.S. Metal poisoning: Threat and management. Al Ameen Med. Sci. 2009, 2, 4–26. [Google Scholar]
- Wang, Y.; Wu, Y.; Luo, K.; Liu, Y.; Zhou, M.; Yans, S.; Shi, H.; Cai, Y. The protective effects of selenium on cadmium-induced oxidative stress and apoptosis via mitochondria pathway in mice Sidney. Food Chem. Toxicol. 2013, 58, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, T.A.; Tarboush, H.A.; Alada, A.; Mohanna, M.A. The effect of selenium and lycopene on oxidative stress in bone tissue in rats exposed to cadmium. Food Nutr. Sci. 2014, 5, 1420–1429. [Google Scholar] [CrossRef] [Green Version]
- El-Boshy, M.E.; Risha, E.F.; Abdelhamid, F.M.; Mubarak, M.S.; Hadda, T.B. Protective effects of selenium against cadmium induced hematological disturbances, immunosuppressive, oxidative stress and hepatorenal damage in rats. Trace Elem. Med. Biol. 2015, 29, 104–110. [Google Scholar] [CrossRef]
- El-Sayed, Y.S.; El-Gazzar, A.M.; El-Nahas, A.F.; Ashry, K.M. Vitamin C modulates cadmium-induced hepatic antioxidants gene transcripts and toxicopathic changes in Nile tilapia, Oreochromis niloticus. Environ. Sci. Pollut. Res. 2016, 23, 1664–1670. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Y.; Liu, J.; Wang, J.; LI, Y.; Li, H.; Zhang, W.; Liao, H. Protective effect of theaflavins on cadmium-induced testicular toxicity in male rats. Food Chem. Toxicol. 2012, 50, 3243–3250. [Google Scholar] [CrossRef]
- Al-Gnami, S.A. Effect of polyphenols which extracted from green tea in reduce toxic effects of cadmium sulfate in rat’s livers. Pharm. Biol. Sci. 2014, 9, 53–58. [Google Scholar]
- Lakshmi, B.V.S.; Sudhakar, M.; Aparna, M. Protective effect of black grapes on cadmium induced hepatotoxicity in rats. World Pharm. Sci. 2014, 2, 276–282. [Google Scholar]
- Merra, E.; Calzaretti, G.; Bobba, A.; Storelli, M.M.; Casalino, E. Antioxidant role of hydroxytyrosol on oxidative stress in cadmium-intoxicated rats: Different effect in spleen and testes. Drug Chem. Toxicol. 2014, 37, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Baiomy, A.A.; Mansour, A.A. Genetic and histopathological responses to cadmium toxicity in rabbit’s kidney and liver: Protection by ginger (Zingiber officinale). Biol. Trace Elem. Res. 2016, 170, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, M.M.; Borowska, S.; Tomczyk, M. Antioxidants as a potential preventive and therapeutic strategy for cadmium. Curr. Drug Targets 2016, 17, 1350–1384. [Google Scholar] [CrossRef]
- Kara, H.; Cevik, A.; Konar, V.; Dayangac, A.; Servi, K. Effects of selenium with vitamin E and melatonin on cadmium-induced oxidative damage in rat liver and kidneys. Biol. Trace Elem. Res. 2008, 125, 236–244. [Google Scholar] [CrossRef]
- Littlefield, N.A.; Hass, B.S. Damage to DNA by cadmium or nickel in the presence of ascorbate. Ann. Clin. Lab. Sci. 1995, 25, 485–492. [Google Scholar]
- Kini, R.D.; Arunkumar, N.; Shetty, B.S.; Bhagyalakshmi, K.; Gokul, M. Potential protective role of beta carotene on cadmium induced brain and kidney damage. Ind. J. Pub. Health Res. Dev. 2019, 10, 532–535. [Google Scholar] [CrossRef]
- Almeer, R.S.; AlBasher, G.I.; Alarifi, S.; Alkahtani, S.; Ali, D.; Moneim, A.E.A. Royal jelly attenuates cadmium-induced nephrotoxicity in male mice. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Jiang, J.; Li, S.; Li, M.; Tan, Y.; Song, S.; Shu, Q.; Huang, J. Glutamate alleviates cadmium toxicity in rice via suppressing cadmium uptake and translocation. J. Hazard. Mater. 2020, 384, 121319. [Google Scholar] [CrossRef]
- Chaudhary, S.; Iram, S.; Raisuddin, S.; Parvez, S. Manganese pretreatment attenuates cadmium induced hepatotoxicity in Swiss albino mice. Trace Elem. Med. Biol. 2015, 29, 284–288. [Google Scholar] [CrossRef]
- Zhang, H.; Reynolds, M. Cadmium exposure in living organisms: A short review. Sci. Total Environ. 2019, 678, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Carocci, A.; Catalano, A.; Sinicropi, M.S. Melatonergic drugs in development. Clin. Pharmacol. 2014, 6, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carocci, A.; Catalano, A.; Sinicropi, M.S.; Genchi, G. Oxidative stress and neurodegeneration: The involvement of iron. Biometals 2018, 31, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Himeno, S.; Sumi, D.; Fujishiro, H. Toxicometallomics of cadmium, manganese and arsenic with special reference to the roles of metal transporters. Toxicol. Res. 2019, 35, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Modica-Napolitano, S.; Kulawiec, M.; Singh, K.K. Mitochondria and human cancer. Curr. Mol. Med. 2007, 7, 121–131. [Google Scholar] [CrossRef]
- Soubannier, V.; McBride, H.M. Positioning mitochondrial plasticity within cellular signaling cascades. Biochim. Biophys. Acta 2009, 1793, 154–170. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F.; Rousseau, D.; Burke, S. The layered structure of human mitochondrial DNA nucleoids. Biol. Chem. 2008, 283, 3665–3675. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, S.; Deecaraman, M.; Kumar, R.; Shamsi, M.B.; Dada, R. Role of reactive oxygen species in the pathogenesis of mitochondrial DNA (mtDNA) mutations in male infertility. Ind. Med. Res. 2009, 129, 127–137. [Google Scholar]
- Belyaeva, E.A.; Dymkowska, D.; Wieckowski, M.R.; Wotczak, L. Mitochondria as an important target in heavy metal toxicity in rat hepatoma AS-30D cells. Toxicol. Appl. Pharmacol. 2008, 231, 34–42. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, J.; Leonard, S.S.; Rao, K.M. Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef]
- Lee, W.K.; Thévenod, F. Cell organelles as targets of mammalian cadmium toxicity. Arch. Toxicol. 2020, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Cannino, G.; Feruggia, E.; Luparello, C.; Rinaldi, A.M. Cadmium and mitochondria. Mitochondrion 2009, 9, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, M.; Pifferi, S.; Ragghianti, M.; Bucci, S.; Rizzuto, R.; Pinton, P. Endoplasmic reticulum stress and alteration in calcium homeostasis are involved in cadmium-induced apoptosis. Cell Calcium 2008, 43, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Belyaeva, E.A.; Dymkowska, D.; Wieckowski, M.R.; Wotczak, L. Reactive oxygen species produced by the mitochondrial respiratory chain are involved in Cd2+-induced injury of rat ascites hepatoma AS-30D cells. Biochim. Biophys. Acta 2006, 1757, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Kundu, S.; Bhattacharyya, A. Mechanism of cadmium induced apoptosis in the immunocyte. Toxicol. Lett. 2008, 177, 83–89. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orreniu, S.; Zhivotovsk, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Gobe, G.; Crane, D. Mitochondria, reactive oxygen species and cadmium toxicity in the kidney. Toxicol. Lett. 2010, 198, 49–55. [Google Scholar] [CrossRef]
- Thévenod, F.; Lee, W.K. Cadmium and cellular signaling cascades: Interactions between cell death and survival pathways. Arch. Toxicol. 2013, 87, 1743–1786. [Google Scholar] [CrossRef]
- Lee, W.K.; Abouhamed, M.; Thévenod, F. Caspase-dependent and -independent pathways for cadmium-induced apoptosis in cultured kidney proximal tubule cells. Am. J. Physiol. Renal Physiol. 2006, 291, F823–F832. [Google Scholar] [CrossRef] [Green Version]
- Margoshes, M.; Vallee, B.L. A cadmium protein from equine kidney cortex. Am. Chem. Soc. 1957, 79, 4813–4814. [Google Scholar] [CrossRef]
- Nielsen, A.E.; Bohr, A.; Penkowa, M. The balance between life and death of cells: Roles of metallothioneins. Biomark. Insights 2007, 1, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabolić, I.; Brelak, D.; Skarica, M.; Herak-Kramberger, C.M. Role of metallothionein in cadmium traffic and toxicity in kidneys and other mammalian organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Moulis, M. The bioinorganic chemistry of cadmium in the context of its toxicity. Met. Ions Life Sci. 2013, 11, 1–29. [Google Scholar] [PubMed]
- Nordberg, G.; Nogawa, K.; Nordberg, M.; Friberg, L. Cadmium. In Handbook of Toxicology of Metals; Nordberg, G., Fowler, B., Nordberb, M., Friberg, L., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 445–486. [Google Scholar]
- Sakulsak, N. Metallothionein: An overview on its metal homeostatic regulation in mammals. Int. Morphol. 2012, 30, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). Scientific opinion of the panel on contaminants in the food chain on a request from the European Commission on cadmium in food. EFSA 2009, 980, 1–139. [Google Scholar]
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, environmental exposure and health outcomes. Environ. Health Perspect. 2010, 118, 182–190. [Google Scholar] [CrossRef]
- Bernard, A. Cadmium and its adverse effects on human health. Indian Med. Res. 2008, 128, 557–564. [Google Scholar]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Health risk assessment of dietary cadmium intake: Do current guidelines indicate how much is safe? Environ. Health Perspect. 2017, 125, 284–288. [Google Scholar] [CrossRef]
- Nishijo, M.; Satarug, S.; Honda, R.; Tsuritani, I.; Aoshima, K. The gender differences in health effects of environmental cadmium exposure and potential mechanisms. Mol. Cell. Biochem. 2004, 255, 87–92. [Google Scholar] [CrossRef]
- Satarug, S.; Moore, M.R. Adverse health effects of chronic exposure to low-level cadmium in foodstuffs and cigarette smoke. Environ. Health Perspect. 2004, 112, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Ujjin, P.; Vanavanitkun, Y.; Baker, R.; Moore, M.R. Influence of body iron store status and cigarette smoking on cadmium body burden of healthy Thai women and men. Toxicol. Lett. 2004, 148, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Final Review of Scientific Information on Cadmium. Available online: https://www.unenvironment.org/resources/report/final-review-scientific-information-cadmium (accessed on 25 May 2020).
- European Food Safety Authority (EFSA). Cadmium dietary exposure in the European population. EFSA 2012, 10, 2551. [Google Scholar] [CrossRef]
- Satarug, S.; Swaddiwudhipong, W.; Ruangyuttikarn, W.; Nishio, M.; Ruiz, P. Modeling cadmium exposures in low- and high-exposure areas in Thailand. Environ. Health Perspect. 2013, 121, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Jamakala, O.; Rani, U. Mitigating role of zinc and iron against cadmium induced toxicity in liver and kidney of male albino rat: A study with reference to metallothionein quantification. Internat. Pharm Sci. 2014, 6, 411–417. [Google Scholar]
- Lin, Y.S.; Ho, W.C.; Caffrey, L.; Sonawane, B. Low serum zinc is associated with elevated risk of cadmium nephrotoxicity. Environ. Res. 2014, 134, 33–38. [Google Scholar] [CrossRef]
- Matović, V.; Buha, A.; Ðukić-Ćosić, D.; Bulat, Z. Insight into the oxidative stress induced by lead and/or cadmium in blood, liver and kidneys. Food Chem. Toxicol. 2015, 78, 130–140. [Google Scholar] [CrossRef]
- Amamou, F.; Nemmiche, S.; Meziane, R.K.; Didi, A.; Yazit, S.A.; Chabane-Sari, D. Protective effect of olive oil and colocynth oil against cadmium-induced oxidative stress in the liver of Wistar rats. Food Chem. Toxicol. 2015, 78, 177–184. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, H.; Liu, X.; Liu, Z. Oxidative stress and Ca2+ signals involved on cadmium-induced apoptosis in rat hepatocyte. Biol. Trace Elem. Res. 2014, 161, 180–189. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; Yousef, M.I.; Kedwany, F.S.; Baghdadi, H.H. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and β-carotene. Food Chem. Toxicol. 2004, 42, 1563–1571. [Google Scholar] [CrossRef]
- Pizzino, G.; Bitto, A.; Interdonato, M.; Galfo, F.; Irrera, N.; Mecchio, A.; Pallio, G.; Ramistella, V.; De Luca, F.; Minutoli, L.; et al. Oxidative stress and DNA repair and detoxification gene expression in adolescents exposed to heavy metals living in the Milazzo-Valle del Mela area (Sicily, Italy). Redox Biol. 2014, 2, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Wang, C.; Liu, H.; Huang, Q.; Wang, M.; Le, Y. Cadmium induced cell apoptosis, DNA damage, decreased DNA repair capacity, and genomic instability during malignant transformation of human bronchial epithelial cells. Int. Med. Sci. 2013, 10, 1485–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.Y.; Zhu, Y.; Chan, K.M. Effects of cadmium on cell proliferation, apoptosis, and proto-oncogene expression in zebrafish liver cells. Aquat. Toxicol. 2014, 157, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Koturbash, I.; Beland, F.A.; Pogribny, I.P. Role of epigenetic events in chemical carcinogenesis-a justification for incorporating epigenetic evaluations in cancer risk assessment. Toxicol. Mech. Methods 2011, 21, 289–297. [Google Scholar] [CrossRef]
- Wallace, D.R.; Taalab, Y.M.; Heinze, S.; Lovaković, B.T.; Pizent, A.; Renieri, E.; Tsatsakis, A.; Farooqi, A.A.; Javorac, D.; Andjelkovic, M.; et al. Toxic-metal-induced alteration in miRNA expression profile as a proposed mechanism for disease development. Cells 2020, 9, 901. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.S.; Bhattacharyya, S.N.; Filipowicz, W. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef]
- Huang, S.Q.; Peng, J.; Qiu, C.X.; Yang, Z.M. Heavy metal-regulated new microRNAs from rice. Inorg. Biochem. 2009, 103, 282–287. [Google Scholar] [CrossRef]
- Szyf, M. The dynamic epigenome and its implications in toxicology. Toxicol. Sci. 2007, 10, 7–23. [Google Scholar] [CrossRef] [Green Version]
- Takiguchi, M.; Achanzar, W.E.; Qu, W.; Li, G.; Waalkes, M.P. Effects of cadmium on DNA-(Cytosine-5) methyltransferase activity and DNA methylation status during cadmium-induced cellular transformation. Exp. Cell Res. 2003, 286, 355–365. [Google Scholar] [CrossRef]
- Benbrahim-Tallaa, L.; Waterland, R.A.; Dill, A.L.; Webber, M.M.; Waalkes, M.P. Tumor suppressor gene inactivation during cadmium-induced malignant transformation of human prostate cells correlates with overexpression of de novo DNA methyltransferase. Environ. Health Perspect. 2007, 115, 1454–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Cullen, B.R. Viral and cellular messenger RNA targets of viral microRNAs. Nature 2009, 457, 421–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, H.; Qi, Y.; Tu, J.; Bai, Y.; Tian, T.; Huang, N.; Wang, Y.; Xiong, F.; Lu, Z.; et al. Assessment of nanomaterial cytotoxicity with SOLiD sequencing-based microRNA expression profiling. Biomaterials 2011, 32, 9021–9030. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, Z.; Zhu, C. Microarray-based analysis of cadmium responsive microRNAs in rice (Oryza sativa). Exp. Bot. 2011, 62, 3563–3573. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Environmental Health Criteria 135. In Cadmium-Environmental Aspects; World Health Organization: Geneva, Switzerland, 1992. [Google Scholar]
- Buha, A.; Matovic, V.; Antonijevic, B.; Bulat, Z.; Curcic, M.; Renieri, E.A.; Tsatsakis, A.M.; Schweitzer, A.; Wallace, D. Overview of Cadmium Thyroid Disrupting Effects and Mechanisms. Int. J. Mol. Sci. 2018, 19, 1501. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Liu, Y.; Xie, C.; Tu, W.; Xia, Y.; Costa, M.; Zhou, X. Cadmium induces histone H3 lysine methylation by inhibiting histone demethylase activity. Toxicol. Sci. 2015, 145, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Kumar, A.; Lal, A.; Pant, M. Cellular mechanisms of cadmium-induced toxicity: A review. Int. Environ. Health Res. 2014, 24, 378–399. [Google Scholar] [CrossRef]
- Giaginis, C.; Gatzidou, E.; Theocharis, S. DNA repair systems as targets of cadmium toxicity. Toxicol. Appl. Pharmacol. 2006, 213, 282–290. [Google Scholar] [CrossRef]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef]
- Martinez-Zamudio, R.; Ha, H.C. Environmental epigenetics in metal exposure. Epigenetics 2011, 6, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.; Takeda, S.; Teraoka-Nishitani, N.; Yamagata, A.; Tanaka, T.; Sasaki, M.; Yasuda, N.; Oda, M.; Okano, T.; Yamahira, K.; et al. Cadmium-induced malignant transformation of rat liver cells: Potential key role and regulatory mechanism of altered apolipoprotein E expression in enhanced invasiveness. Toxicology 2017, 382, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zimtaa, A.-A.; Schitcub, V.; Gurzaud, E.; Stavarug, C.; Mandah, G.; Szedlacseki, S.; Berindan-Neagoe, I. Biological and molecular modifications induced by cadmium and arsenic during breast and prostate cancer development. Environ. Res. 2019, 178, 108700. [Google Scholar] [CrossRef] [PubMed]
- Nagata, C.; Nagao, Y.; Nakamura, K.; Wada, K.; Tamai, Y.; Tsuji, M.; Yamamoto, S.; Kashiki, Y. Cadmium exposure and the risk of breast cancer in Japanese women. Breast Cancer Res. Treat. 2013, 138, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.S.; Martens, D.S.; Hara, A.; Plusquin, M.; Vangronsveld, J.; Roels, H.A.; Staessen, J.A. Association of total cancer and lung cancer with environmental exposure to cadmium: The meta-analytical evidence. Cancer Causes Control. 2015, 26, 1281–1288. [Google Scholar] [CrossRef]
- Peng, L.; Huang, Y.; Zhang, J.; Peng, Y.; Lin, X.; Wu, K.; Huo, X. Cadmium exposure and the risk of breast cancer in Chaoshan population of southeast China. Environ. Sci. Pollut. Res. Int. 2015, 22, 19870–19878. [Google Scholar] [CrossRef]
- Peng, L.; Wang, X.; Huo, X.; Xu, X.; Lin, K.; Zhang, J.; Huang, Y.; Wu, K. Blood cadmium burden and the risk of nasopharyngeal carcinoma: A case-control study in Chinese Chaoshan population. Environ. Sci. Pollut. Res. Int. 2015, 22, 12323–12331. [Google Scholar] [CrossRef]
- Julin, B.; Wolk, A.; Johansson, E.; Andersson, S.O.; Andrén, O.; Akesson, A. Dietary cadmium exposure and prostate cancer incidence: A population-based prospective cohort study. Br. J. Cancer 2012, 107, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Feki-Tounsi, M.; Hamza-Chaffai, A. Cadmium as a possible cause of bladder cancer: A review of accumulated evidence. Environ. Sci. Pollut. Res. 2014, 18, 10561–10573. [Google Scholar] [CrossRef]
- Chen, C.; Xun, P.; Nishio, M.; Sekikawa, A.; He, K. Cadmium exposure and risk of pancreatic cancer: A meta-analysis of prospective cohort studies and case-control studies among individuals without occupational exposure history. Environ. Sci. Pollut. Res. 2015, 22, 17465–17474. [Google Scholar] [CrossRef] [Green Version]
- Song, J.K.; Luo, H.; Yin, X.H.; Huang, G.L.; Luo, S.Y.; Lin, D.R.; Yuan, D.B.; Zhang, W.; Zhu, J.G. Association between cadmium exposure and renal cancer risk: A meta-analysis of observational studies. Sci. Rep. 2015, 5, 17976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Agrawal, M. Effects of sewage sludge amendment on heavy metal accumulation and consequent responses of Beta vulgaris plants. Chemosphere 2007, 67, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Ghoochani, M.; Rastkari, N.; Yunesian, M.; Nodehi, R.N.; Mesdaghinia, A.; Houshiarra, A.; Shamsipour, M.; Dehghani, M.H. What do we know about exposure of Iranians to cadmium? Findings from a systematic review. Environ. Sci. Pollut. Res. 2018, 25, 1033–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratão, P.L.; Polle, A.; Lea, P.; Azevedo, R.A. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol. 2005, 32, 481–494. [Google Scholar] [CrossRef]
- Ahn, C.K.; Park, D.; Woo, S.H.; Park, M. Removal of cationic heavy metal from aqueous solution by activated carbon impregnated with anionic surfactants. Hazard. Mater. 2009, 164, 1130–1136. [Google Scholar] [CrossRef]
- Jadia, C.D.; Fulekar, M.H. Phytoremediation: The application of vermicompost to remove zinc, cadmium, copper, nickel and lead by sunflower plant. Environ. Eng. Manag. 2008, 7, 547–558. [Google Scholar]
- Al-Dhaibani, A.N.; EL-Nakhlawy, F.S.; Alsolaimani, S.G.; Almehmadi, F.M. Phytoremediation of cadmium contaminated soil by sunflower. Aust. Basic Appl. Sci. 2013, 7, 888–894. [Google Scholar]
- Jain, M.; Garg, V.K.; Paliwal, R.; Kadirvelu, K.; Chaudhry, S. Optimization of cadmium (II) removal from water using sunflower waste carbon–a statistical approach. Toxin Rev. 2020, 1–10. [Google Scholar] [CrossRef]
- Goswami, S.; Das, S. A Study on cadmium phytoremediation potential of Indian mustard, Brassica juncea. Int. Phytoremediat. 2015, 17, 583–588. [Google Scholar] [CrossRef]
- Iori, V.; Pietrini, F.; Bianconi, D.; Mughini, G.; Massacci, A.; Zacchini, M. Analysis of biometric, physiological, and biochemical traits to evaluate the cadmium phytoremediation ability of eucalypt plants under hydroponics. iForest 2017, 10, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Bora, T.; Dutta, J. Applications of nanotechnology in wastewater treatment—A review. Nanosci. Nanotechnol. 2014, 14, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Stietiya, M.H.; Wang, J.J. Zinc and cadmium adsorption to aluminum oxide nanoparticles affected by naturally occurring ligands. J. Environ. Qual. 2014, 43, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Al-Qahtani, K.M. Cadmium removal from aqueous solution by green synthesis zero valent silver noanoparticles with Benjamina leaves extract. Egypt. J. Aquat. Res. 2017, 43, 269–274. [Google Scholar] [CrossRef]
- Koju, N.K.; Song, X.; Wang, Q.; Hu, Z.; Colombo, C. Cadmium removal from simulated groundwater using alumina nanoparticles: Behaviors and mechanisms. Environ. Pollut. 2018, 240, 255–266. [Google Scholar] [CrossRef]
- Schaefer, H.R.; Dennis, S.; Fitzpatrick, S. Cadmium: Mitigation strategies to reduce dietary exposure. J. Food Sci. 2020, 85. [Google Scholar] [CrossRef]
- Zhang, L.; Lei, Q.; Cheng, Y.; Xie, Y.; Qian, H.; Guo, Y.; Chen, Y.; Yao, W. Study on the removal of cadmium in rice using microbial fermentation method. J. Food Sci. 2017, 82, 1467–1474. [Google Scholar] [CrossRef]
- Zhai, Q.; Guo, Y.; Tang, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Removal of cadmium from rice by Lactobacillus plantarum fermentation. Food Control 2019, 96, 357–364. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, Y.; Lv, C.; Wu, W.; Qin, S. Study on optimization of removing cadmium by lactobacillus fermentation and its effect on physicochemical and quality properties of rice noodles. Food Control 2019, 106, 106740. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Atomic number | 48 |
| Atomic weight | 112.41 u |
| Atomic radius | 155 pm |
| Electronic configuration | [Kr]4d105s2 |
| Melting point | 321.07 °C |
| Boiling point | 767.3 °C |
| Density at 20 °C | 8.65 g/cm3 |
| Reduction potential Cd2+ + 2e− → Cd(s) | −0.40 E° |
| Heat of fusion | 6.21 kJ/mol |
| Heat of vaporization | 99.6 kJ/mol |
| Electronegativity (Pauling scale) | 1.69 |
| First ionization energy | 867.8 kJ/mol |
| Second ionization energy | 1631.4 kJ/mol |
| Polyphenolic Substances | mg Polyphenol/100 g |
|---|---|
| Cloves | 15,200 |
| Peppermint | 11,960 |
| Anise | 5460 |
| Cocoa powder | 3450 |
| Dark chocolate | 1660 |
| Fruits | |
| Black chokeberries | 1700 |
| Highbush blueberries | 560 |
| Blackberries | 260 |
| Strawberries | 235 |
| Red raspberries | 215 |
| Black currants | 758 |
| Plums | 377 |
| Sweet cherries | 274 |
| Apples | 136 |
| Nuts | |
| Hazelnuts | 495 |
| Walnuts | 28 |
| Almonds | 187 |
| Pecans | 493 |
| Vegetables | |
| Green olives | 346 |
| Artichokes | 260 |
| Red onions | 168 |
| Chicory | 166 |
| Spinach | 120 |
| Extra-virgin olive oil | 62 |
| Beverages | |
| Coffee | 214 |
| Black tea | 102 |
| Red wine | 100 |
| Green tea | 90 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. https://doi.org/10.3390/ijerph17113782
Genchi G, Sinicropi MS, Lauria G, Carocci A, Catalano A. The Effects of Cadmium Toxicity. International Journal of Environmental Research and Public Health. 2020; 17(11):3782. https://doi.org/10.3390/ijerph17113782
Chicago/Turabian StyleGenchi, Giuseppe, Maria Stefania Sinicropi, Graziantonio Lauria, Alessia Carocci, and Alessia Catalano. 2020. "The Effects of Cadmium Toxicity" International Journal of Environmental Research and Public Health 17, no. 11: 3782. https://doi.org/10.3390/ijerph17113782
APA StyleGenchi, G., Sinicropi, M. S., Lauria, G., Carocci, A., & Catalano, A. (2020). The Effects of Cadmium Toxicity. International Journal of Environmental Research and Public Health, 17(11), 3782. https://doi.org/10.3390/ijerph17113782