N-Fertilizer (Urea) Enhances the Phytoextraction of Cadmium through Solanum nigrum L.

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Soil Pot Experiments
2.3. Plant Harvesting
2.4. Determination of Photosynthetic, Chlorophyll, and Carotenoid Content Parameters
2.5. Determination of EL, MDA, H2O2, and Antioxidant Enzyme Concentration
2.6. Cadmium Determination in Plants, Translocation Factor and Bioaccumulation Factor
2.7. Statistical Analysis
3. Results
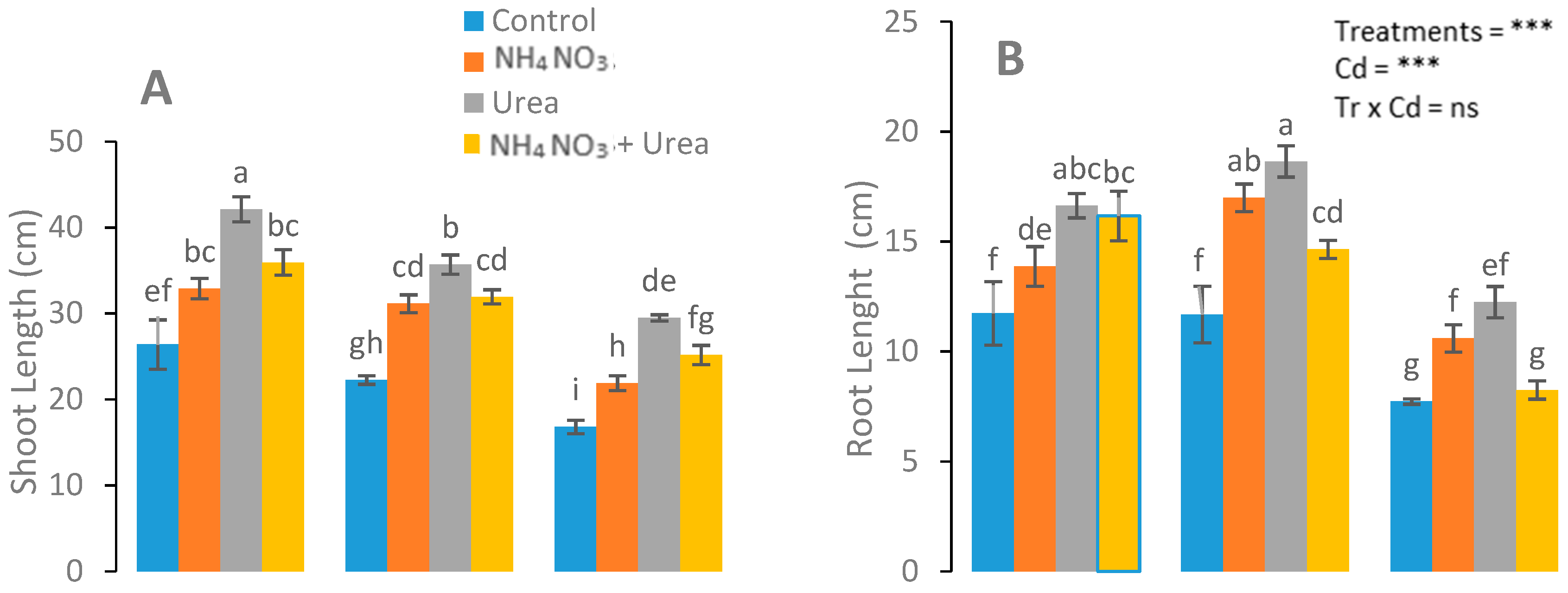
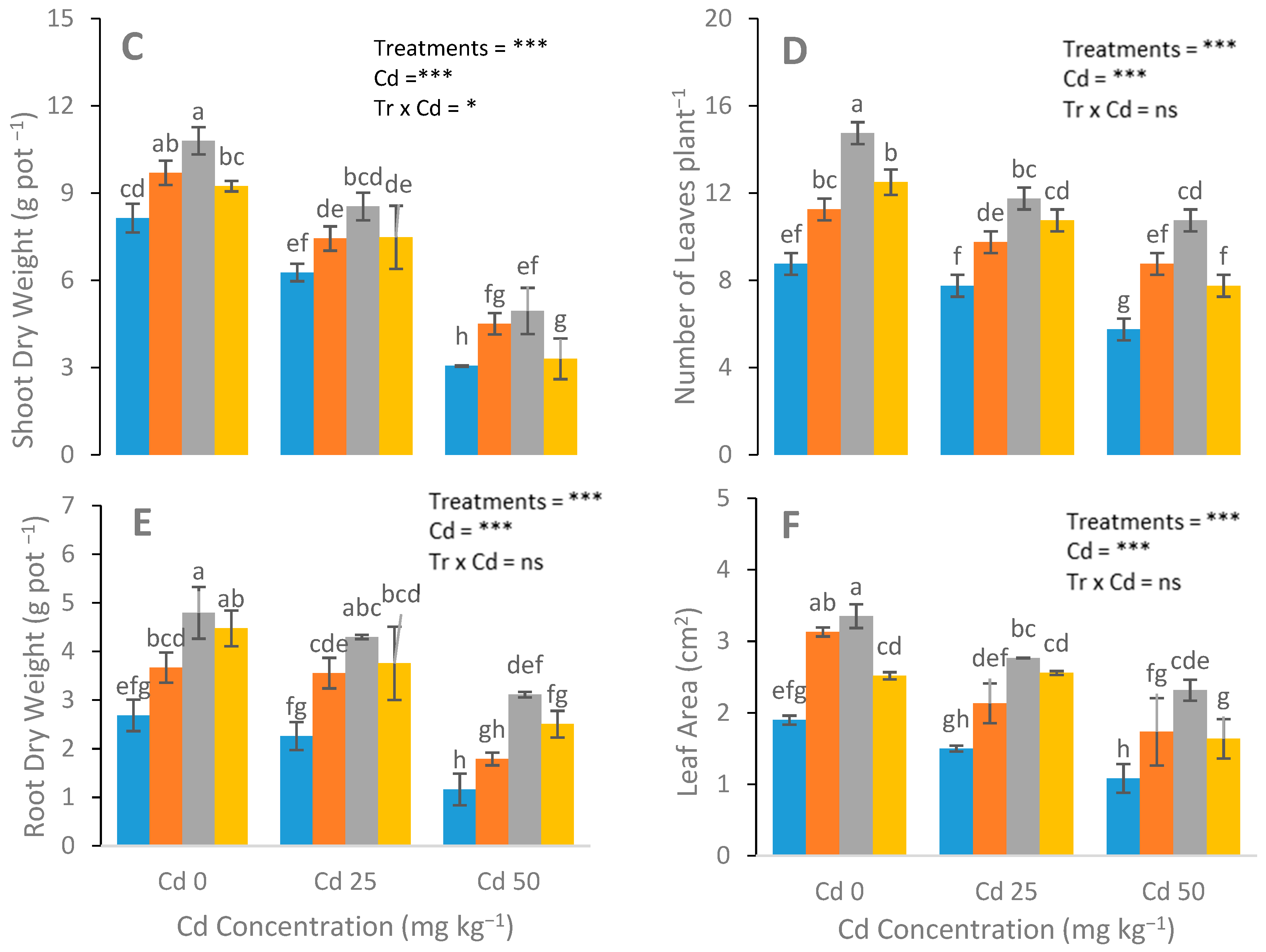
3.1. Assessment of N-Fertilizer on Growth and Biomass
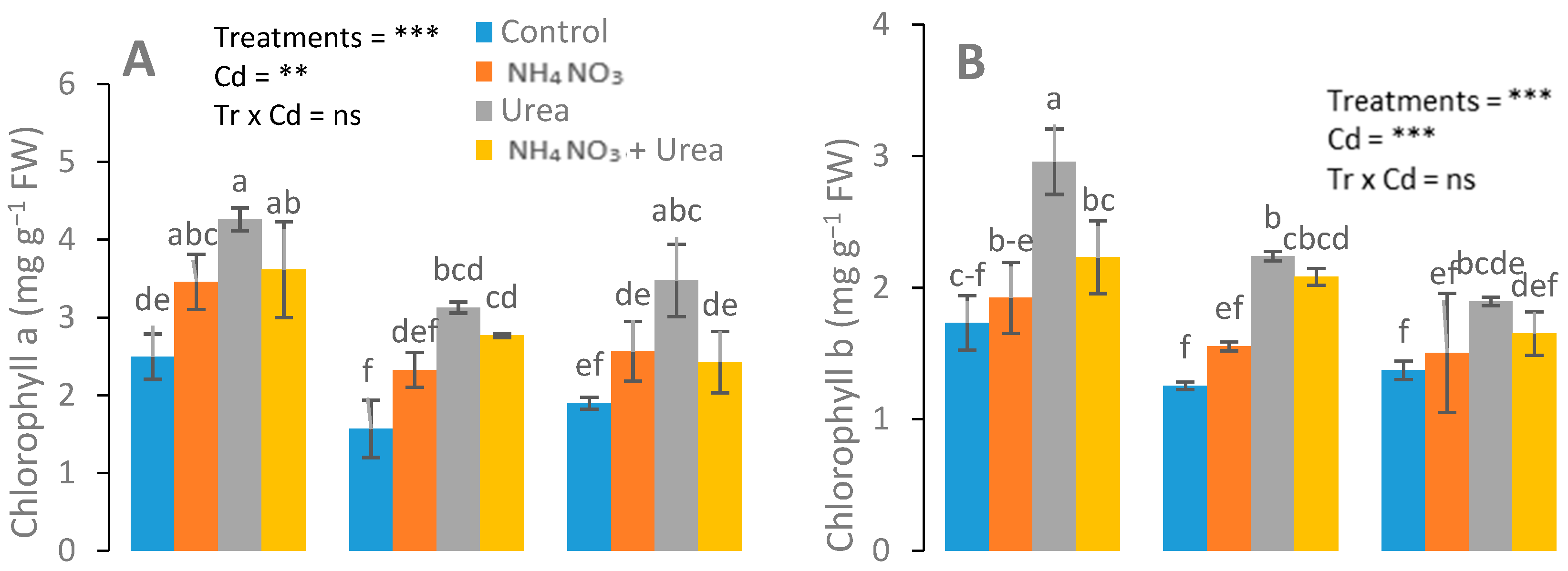
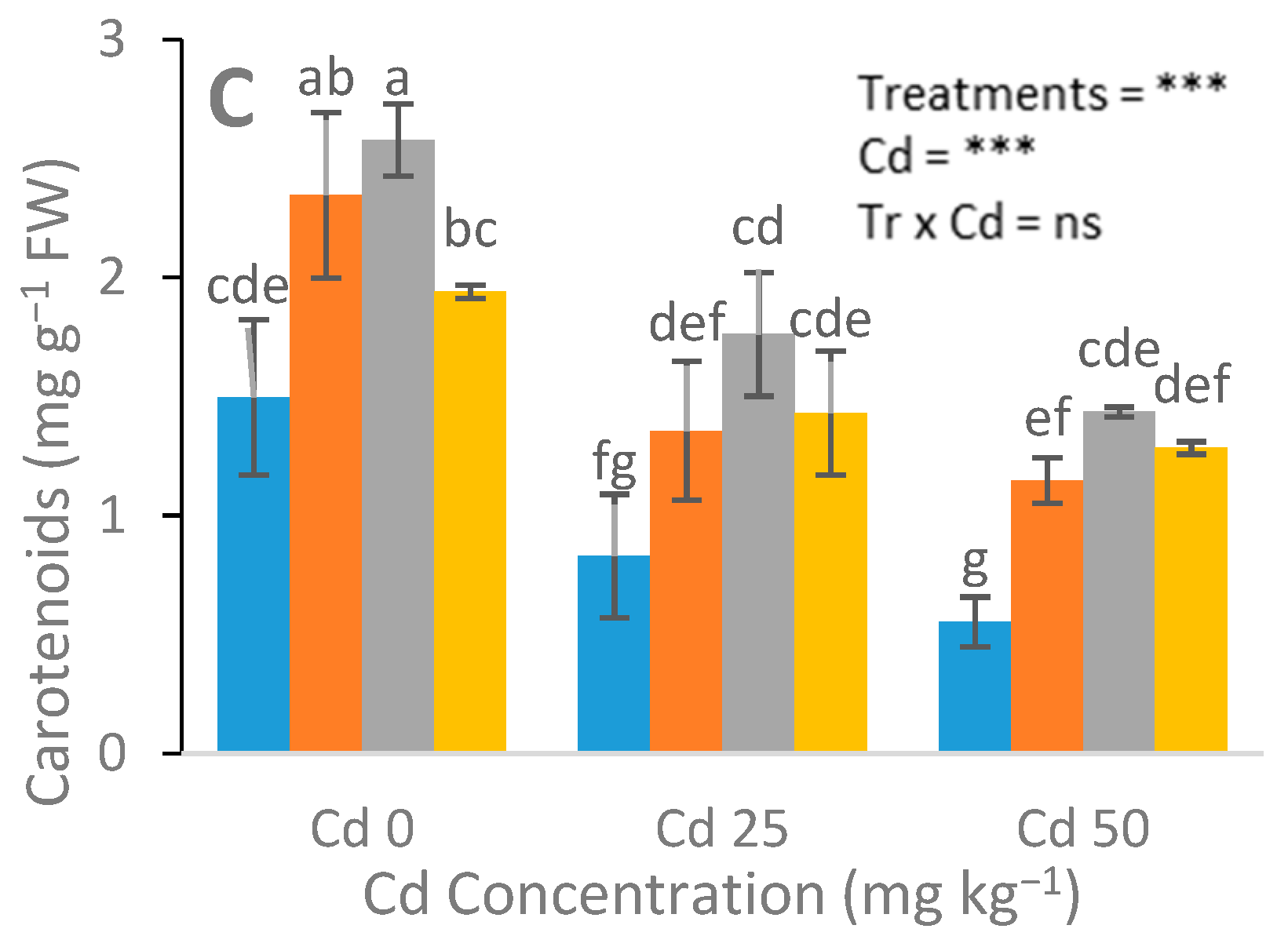
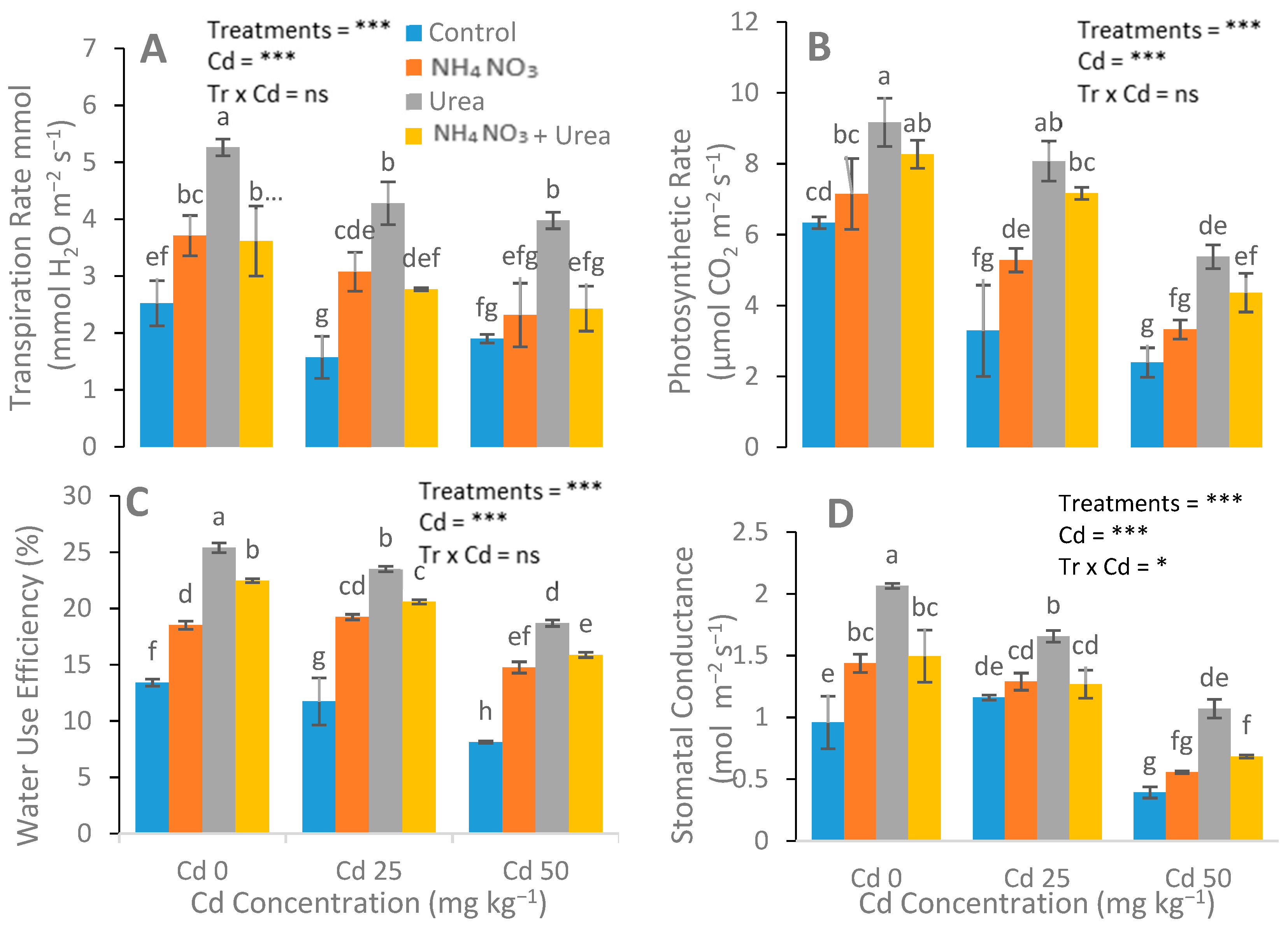
3.2. Assessment of Gas Exchange, Chlorophyll, and Carotenoid Content Attributes
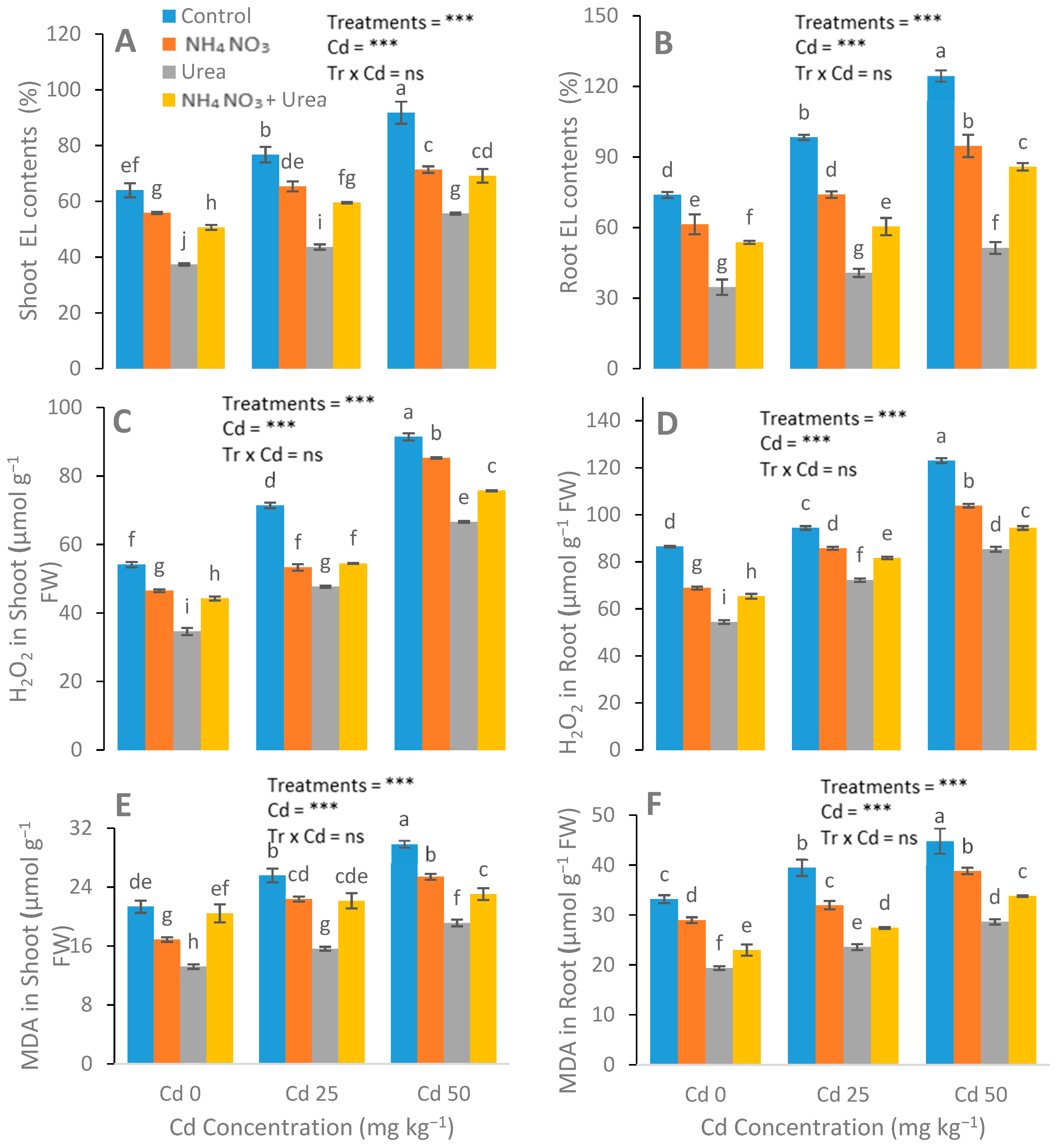
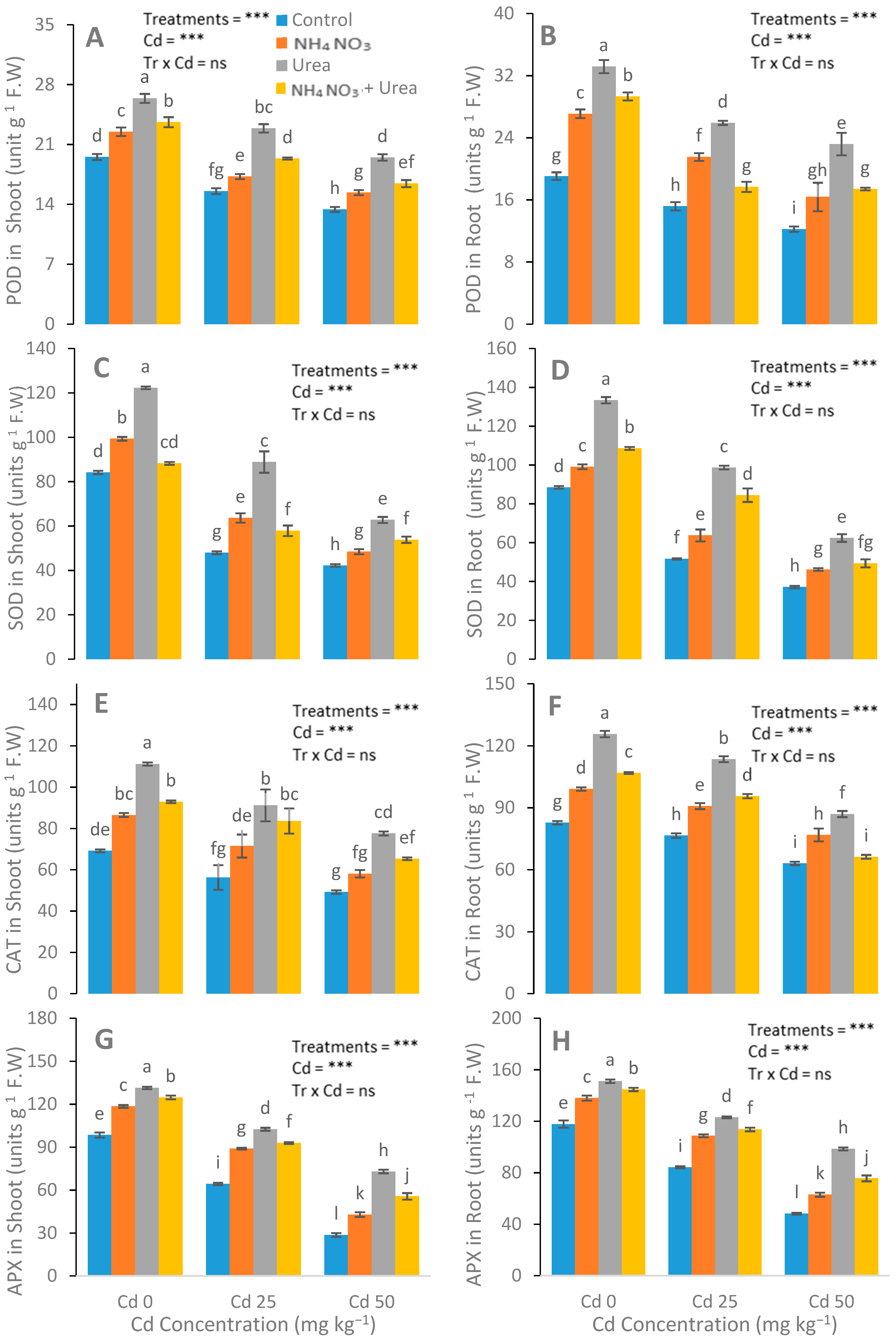
3.3. Assessment of Antioxidant Enzyme Activities and EL, MDA, and H2O2
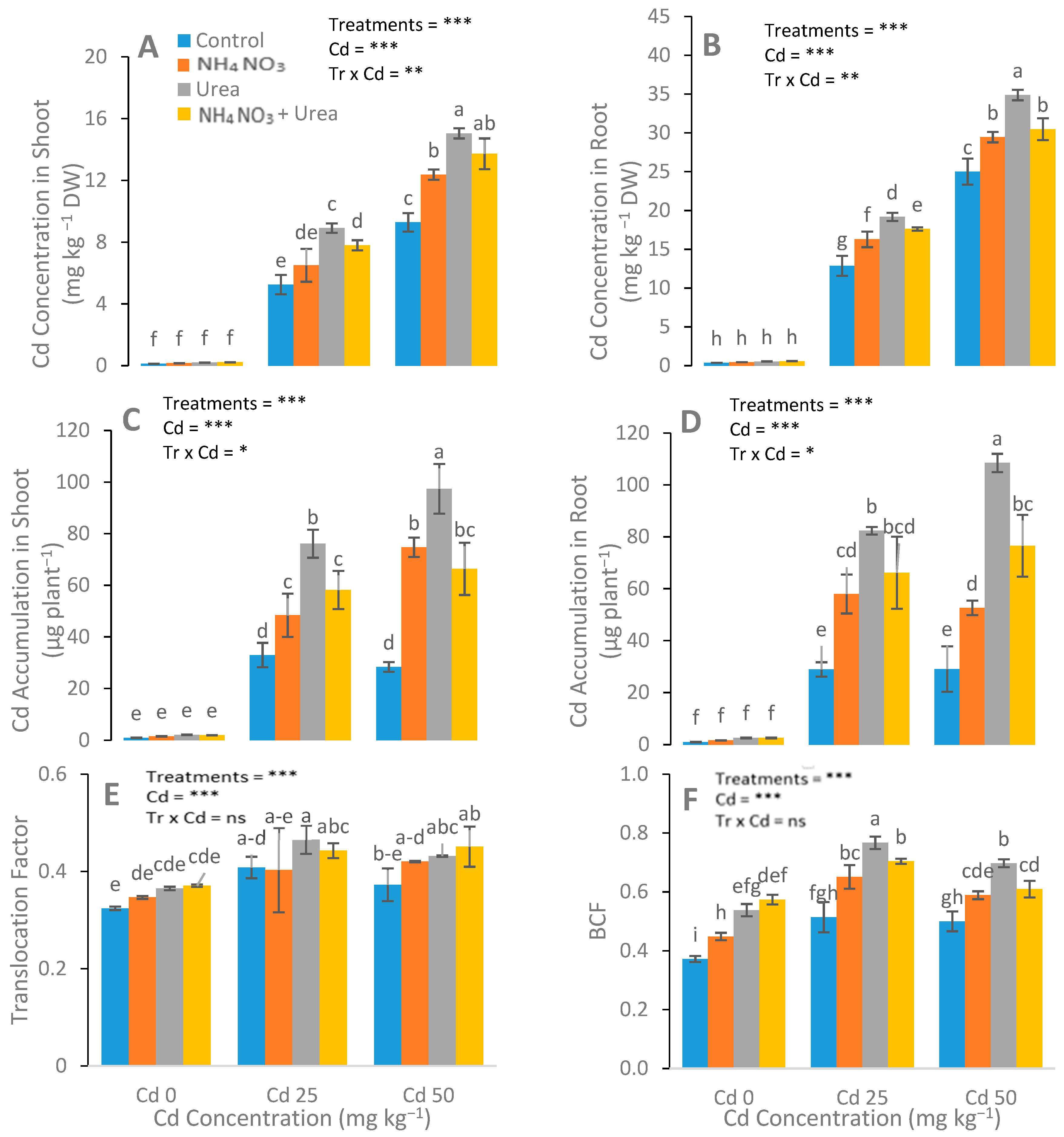
3.4. Assessment of Concentration of Cd in Plants
4. Discussion
4.1. Biomass and Plant Growth
4.2. Photosynthetic Pigments
4.3. Oxidative Stress, Antioxidant Enzymes, and Cd Concentration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldstone, J.A. Demography, environment, and security. Environ. Confl. Routledge 2018, 8, 84–108. [Google Scholar]
- Murtaza, G.; Javed, W.; Hussain, A.; Wahid, A.; Murtaza, B.; Owens, G. Metal uptake via phosphate fertilizer and city sewage in cereal and legume crops in Pakistan. Environ. Sci. Pollut. Res. 2015, 22, 9136–9147. [Google Scholar] [CrossRef] [PubMed]
- Markovic, J.; Jovic, M.; Smiciklas, I.; Sljivic-Ivanovic, M.; Onjia, A.; Trivunac, K.; Popovic, A. Cadmium retention and distribution in contaminated soil: Effects and interactions of soil properties, contamination level, aging time and in situ immobilization agents. Ecotoxicol. Environ. Saf. 2019, 174, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.; Rehman, M.Z.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Gill, R.A.; Yang, S.; Gill, M.B.; Ali, S.; Rafiq, M.T.; Zhou, W. Hydrogen sulfide alleviates cadmium-induced morpho-physiological and ultrastructural changes in Brassica napus. Ecotoxicol. Environ. Saf. 2014, 110, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Khalid, H.; Akmal, F.; Ali, S.; Rizwan, M.; Qayyum, M.F.; Iqbal, M.; Khalid, M.U.; Azhar, M. Effect of limestone, lignite and biochar applied alone and combined on cadmium uptake in wheat and rice under rotation in and effluent irrigated field. Environ. Pollut. 2017, 227, 560–568. [Google Scholar] [CrossRef]
- Gill, R.A.; Ali, B.; Islam, F.; Farooq, M.A.; Gill, M.B.; Mwamba, T.M.; Zhou, W. Physiological and molecular analyses of black and yellow seeded Brassica napus regulated by 5-aminolivulinic acid under chromium stress. Plant Physiol. Biochem. 2015, 94, 130–143. [Google Scholar] [CrossRef]
- Xia, S.; Song, Z.; Jeyakumar, P.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; Wang, H. A critical review on bioremediation technologies for Cr (VI)-contaminated soils and wastewater. Critic. Rev. Environ. Sci. Technol. 2019, 49, 1027–1078. [Google Scholar] [CrossRef]
- Deng, Z.; Cao, L. Fungal endophytes and their interactions with plants in phytoremediation: A review. Chemosphere 2017, 168, 1100–1106. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R.; Juzoń, K.; Borek, M.; Antonkiewicz, J. Photosynthetic response of cabbage in cadmium-spiked soil. Photosynthetica 2019, 57, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.H.; Song, Y.F.; Jiang, Y.; Tang, X.; Tong, Y.A.; Gao, P.; Han, W. A two-year field study of phytoremediation using Solanum nigrum L. in China. Int. J. Phytorem. 2016, 18, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.X.; Luo, Q.; Liu, S.L.; Zhao, Y.; Long, Y.; Pan, Y.Z. Screening ornamental plants to identify potential Cd hyperaccumulators for bioremediation. Ecotoxicol. Environ. Saf. 2018, 162, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Wei, S.; Twardowska, I.; Han, R.; Xu, L. Hyperaccumulating potential of Bidens pilosa L. for Cd and elucidation of its translocation behavior based on cell membrane permeability. Environ. Sci. Pollut. 2017, 24, 23161–23167. [Google Scholar] [CrossRef]
- Jacobs, A.; Noret, N.; Van Baekel, A.; Liénard, A.; Colinet, G.; Drouet, T. Influence of edaphic conditions and nitrogen fertilizers on cadmium and zinc phytoextraction efficiency of Noccaea caerulescens. Sci. Total Environ. 2019, 665, 649–659. [Google Scholar] [CrossRef]
- Li, T.Q.; Di, Z.Z.; Yang, X.E.; Sparks, D.L. Effects of dissolved organic matter from the rhizosphere of the hyperaccumulator Sedum alfredii on sorption of zinc and cadmium by different soils. J. Hazard. Mater. 2011, 192, 1616–1622. [Google Scholar] [CrossRef]
- Álvarez-Mateos, P.; Alés-Álvarez, F.J.; García-Martín, J.F. Phytoremediation of highly contaminated mining soils by Jatropha curcas L. and production of catalytic carbons from the generated biomass. J. Environ. Manag. 2019, 231, 886–895. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Y.F.; Chang, S.X.; Yang, Y.F.; Fu, S.L.; Jiang, P.K.; Luo, Y.; Yang, M.; Chen, Z.H.; Hu, S.D.; et al. Biochar reduces soil heterotrophic respiration in a subtropical plantation through increasing soil organic carbon recalcitrancy and decreasing carbon-degrading microbial activity. Soil Biol. Biochem. 2018, 122, 173–185. [Google Scholar] [CrossRef]
- Pan, H.; Xie, K.; Zhang, Q.; Jia, Z.; Xu, J.; Di, H.; Li, Y. Archaea and bacteria respectively dominate nitrification in lightly and heavily grazed soil in a grassland system. Biol. Fertil. Soils 2018, 54, 41–54. [Google Scholar] [CrossRef]
- Xiao, M.L.; Zang, H.D.; Liu, S.L.; Ye, R.Z.; Zhu, Z.K.; Su, Y.R.; Wu, J.S.; Ge, T.D. Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice–soil systems: A 13C-CO2 pulse labeling study. Plant Soil 2019, 445, 101–112. [Google Scholar] [CrossRef]
- Arnamwong, S.; Wu, L.H.; Hu, P.J.; Yuan, C.; Thiravetyan, P.; Luo, Y.M.; Christie, P. Phytoextraction of cadmium and zinc by Sedum plumbizincicola using different nitrogen fertilizers, a nitrification inhibitor and a urease inhibitor. Int. J. Phytorem. 2015, 17, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ding, Y.; Zhang, Q.; Liu, X.; Xu, J.; Li, Y.; Di, H. Heterotrophic nitrification and denitrification are the main sources of nitrous oxide in two paddy soils. Plant Soil 2018, 445, 39–53. [Google Scholar] [CrossRef]
- Yang, Z.B.; Liu, L.X.; Lv, Y.F.; Cheng, Z.; Xu, X.X.; Xian, J.R.; Zhu, X.M.; Yang, Y.X. Metal availability, soil nutrient, and enzyme activity in response to application of organic amendments in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2018, 25, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Yavari, S.; Malakahmad, A.; Sapari, N.B.; Yavari, S. Nutrients balancefor improvement of phytoremediation ability of teak seedlings (Tectona grandis). J. Plant Nutr. 2018, 41, 545–551. [Google Scholar] [CrossRef]
- Liu, Y.L.; Ge, T.D.; Jun, Y.; Liu, S.L.; Shibistova, O.; Wang, P.; Wang, J.K.; Li, Y.; Guggenberger, G.; Kuzyakov, Y.; et al. Initial utilization of rhizodeposits with rice growth in paddy soils: Rhizosphere and N fertilization effects. Geoderma. Res. 2019, 338, 30–39. [Google Scholar] [CrossRef]
- Yang, W.; Dai, H.; Skuza, L.; Wei, S. Strengthening role and the mechanism of optimum nitrogen addition in relation to Solanum nigrum L. Cd hyperaccumulation in soil. Ecotoxicol. Environ. saf. 2019, 182, 109444. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Constable and Co. Ltd.: London, UK, 1962; p. 219. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeny, D.R. Methods of Soil Analysis (Part 2). In Chemical and Microbiological Properties; SSSA: Madison, WI, USA, 1982; Volume 9. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids pigments of photosynthetic biomembranes. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Ecosyst. Environ. Res. 1998, 253, 122–130. [Google Scholar] [CrossRef]
- Zhang, X.Z. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. In Research Methodology of Crop Physiology; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Zhang, R.R.; Liu, Y.; Xue, W.L.; Chen, R.X.; Du, S.T.; Jin, C.W. Slow release nitrogen fertilizers can improve yield and reduce Cd concentration in pakchoi (Brassica chinensis L.) grown in Cd contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 25074–25083. [Google Scholar] [CrossRef]
- Zhou, G.D.; Guo, J.M.; Yang, J.; Yang, J.X. Effect of fertilizers on Cd accumulation and subcellular distribution of two cosmos species (Cosmos sulphureus and Cosmos bipinnata). Int. J. Phytorem. Res. 2018, 20, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Dou, C.; Li, Y.; Wang, H.; Niazi, N.K.; Zhang, S.; Ye, Z. Nitrogen fertilizer enhances zinc and cadmium uptake by hyperaccumulator Sedum alfredii Hance. J. Soils Sediments 2019, 20, 320–329. [Google Scholar] [CrossRef]
- Rabelo, F.H.S.; Azevedo, R.A.; Monteiro, F.A. Proper supply of S increases GSH synthesis in the establishment and reduces tiller mortality during the regrowth of Tanzania guinea grass used for Cd phytoextraction. J. Soils Sediments. Res. 2017, 17, 1427–1436. [Google Scholar] [CrossRef]
- Gouia, H.; Ghorbala, M.H.; Meyer, C. Effects of cadmium on activity of nitrate reductase and on other enzymes of the nitrate assimilation pathway in bean. Plant Physiol. Biochem. Res. 2000, 38, 629–638. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, P.; Kopittke, P.M.; Wang, A.; Sale, P.W.; Tang, C. Cadmium accumulation is enhanced by ammonium compared to nitrate in two hyperaccumulators, without affecting speciation. J. Exp. Bot. 2016, 67, 5041–5050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Li, Y.; Zhou, Q.; Srivastava, M.; Chiu, S.; Zhan, J.; Wu, Z.; Sun, T. Effect of fertilizer amendments on phytoremediation of Cd-contaminated soil by a newly discovered hyperaccumulator Solanum nigrum L. J. Hazard. Mater. 2010, 176, 269–273. [Google Scholar] [CrossRef]
- Ye, X.; Hu, H.; Li, H.; Xiong, Q.; Gao, H. Combined nitrogen fertilizer and wheat straw increases the cadmium phytoextraction efficiency of Tagetes patula. Ecotoxicol. Environ. Saf. 2019, 170, 210–217. [Google Scholar] [CrossRef]
- Yang, W.; Dai, H.; Dou, X.; Zhang, Q.; Wei, S. Effect and mechanism of commonly used four nitrogen fertilizers and three organic fertilizers on Solanum nigrum L. hyperaccumulating Cd. Environ. Sci. Pollut. Res. 2019, 26, 12940–12947. [Google Scholar] [CrossRef]
- Hassan, M.J.; Zhang, G.; Zhu, Z. Influence of cadmium toxicity on plant growth and nitrogen uptake in rice as affected by nitrogen form. J. Plant Nutr. 2008, 3, 251–262. [Google Scholar] [CrossRef]
- Sanita, L.; Di Toppi, S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Kastori, R.; Petrovic, N.; Arsenijevic-Maksimovic, I. Heavy metals and plants. In Heavy Metals in the Environment; Kastori, R., Ed.; Feljton: Novi Sad, Serbia, 1997; pp. 195–257. [Google Scholar]
- Pakovic, D.; Plesnicar, M.; Arsenijevic-Marksimovic, I.; Petrovic, N. Effects of nitrogen nutrition on photosynthesis in Cd-treated plants. Ann. Bot. 2000, 86, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Descourviers, P.; Kunert, K.J. Protection against oxygen radicals: An important defense mechanism studied in transgenic plants. Plant Cell Environ. Res. 1994, 17, 507–523. [Google Scholar] [CrossRef]
- Jalloh, M.A.; Chen, J.; Zhen, F.; Zhang, G. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard. Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef]
- Malmir, H.A. Comparison of antioxidant enzyme activities in leaves stems and roots of Sorghum (Sorghum bicolor L.) exposed to Chromium (VI). Afr. J. Plant Sci. 2011, 5, 436–444. [Google Scholar]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Pandey, V.; Dixit, V.; Shyam, R. Antioxidative responses in relation to growth of mustard (Brassica juncea cv. Pusa Jaikisan) plants exposed to hexavalent chromium. Chemosphere 2005, 61, 40–47. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Fujita, M. EDTA reduces cadmium toxicity in mustard (Brassica juncea L.) by enhancing metal chelation, antioxidant defense and glyoxalase systems. Acta Agrobot. 2019, 72, 1722. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Nahar, K.; Rahman, M.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Physiological and biochemical mechanisms of spermine-induced cadmium stress tolerance in mung bean (Vigna radiata L.) seedlings. Environ. Sci. Pollut. Res. 2016, 23, 21206–21218. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Element Res. 2012, 149, 248–261. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Texture | Sandy Loam |
|---|---|
| Silt | 14.72% |
| Sand | 68.07% |
| Clay | 17.21% |
| EC | 1.83 dS m−1 |
| pH | 7.72 |
| SAR | 1.93 (mmol L−1)1/2 |
| Available P | 2.20 mg kg−1 |
| Organic matter | 0.64% |
| HCO3−1 | 2.58 mmol L−1 |
| SO4−2 | 11.69 mmol L−1 |
| Cl− | 5.35 mmol L−1 |
| Ca2+ + Mg2+ | 14.26 mmol L−1 |
| K+ | 0.03 mmol L−1 |
| Na+ | 5.48 mmol L−1 |
| Available Zn2+ | 0.81 mg kg−1 |
| Available Cu2+ | 0.34 mg kg−1 |
| Available Cd2+ | 0.09 mg kg−1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maqbool, A.; Ali, S.; Rizwan, M.; Arif, M.S.; Yasmeen, T.; Riaz, M.; Hussain, A.; Noreen, S.; Abdel-Daim, M.M.; Alkahtani, S. N-Fertilizer (Urea) Enhances the Phytoextraction of Cadmium through Solanum nigrum L. Int. J. Environ. Res. Public Health 2020, 17, 3850. https://doi.org/10.3390/ijerph17113850
Maqbool A, Ali S, Rizwan M, Arif MS, Yasmeen T, Riaz M, Hussain A, Noreen S, Abdel-Daim MM, Alkahtani S. N-Fertilizer (Urea) Enhances the Phytoextraction of Cadmium through Solanum nigrum L. International Journal of Environmental Research and Public Health. 2020; 17(11):3850. https://doi.org/10.3390/ijerph17113850
Chicago/Turabian StyleMaqbool, Arosha, Shafaqat Ali, Muhammad Rizwan, Muhammad Saleem Arif, Tahira Yasmeen, Muhammad Riaz, Afzal Hussain, Shamaila Noreen, Mohamed M. Abdel-Daim, and Saad Alkahtani. 2020. "N-Fertilizer (Urea) Enhances the Phytoextraction of Cadmium through Solanum nigrum L." International Journal of Environmental Research and Public Health 17, no. 11: 3850. https://doi.org/10.3390/ijerph17113850
APA StyleMaqbool, A., Ali, S., Rizwan, M., Arif, M. S., Yasmeen, T., Riaz, M., Hussain, A., Noreen, S., Abdel-Daim, M. M., & Alkahtani, S. (2020). N-Fertilizer (Urea) Enhances the Phytoextraction of Cadmium through Solanum nigrum L. International Journal of Environmental Research and Public Health, 17(11), 3850. https://doi.org/10.3390/ijerph17113850