Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4


 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
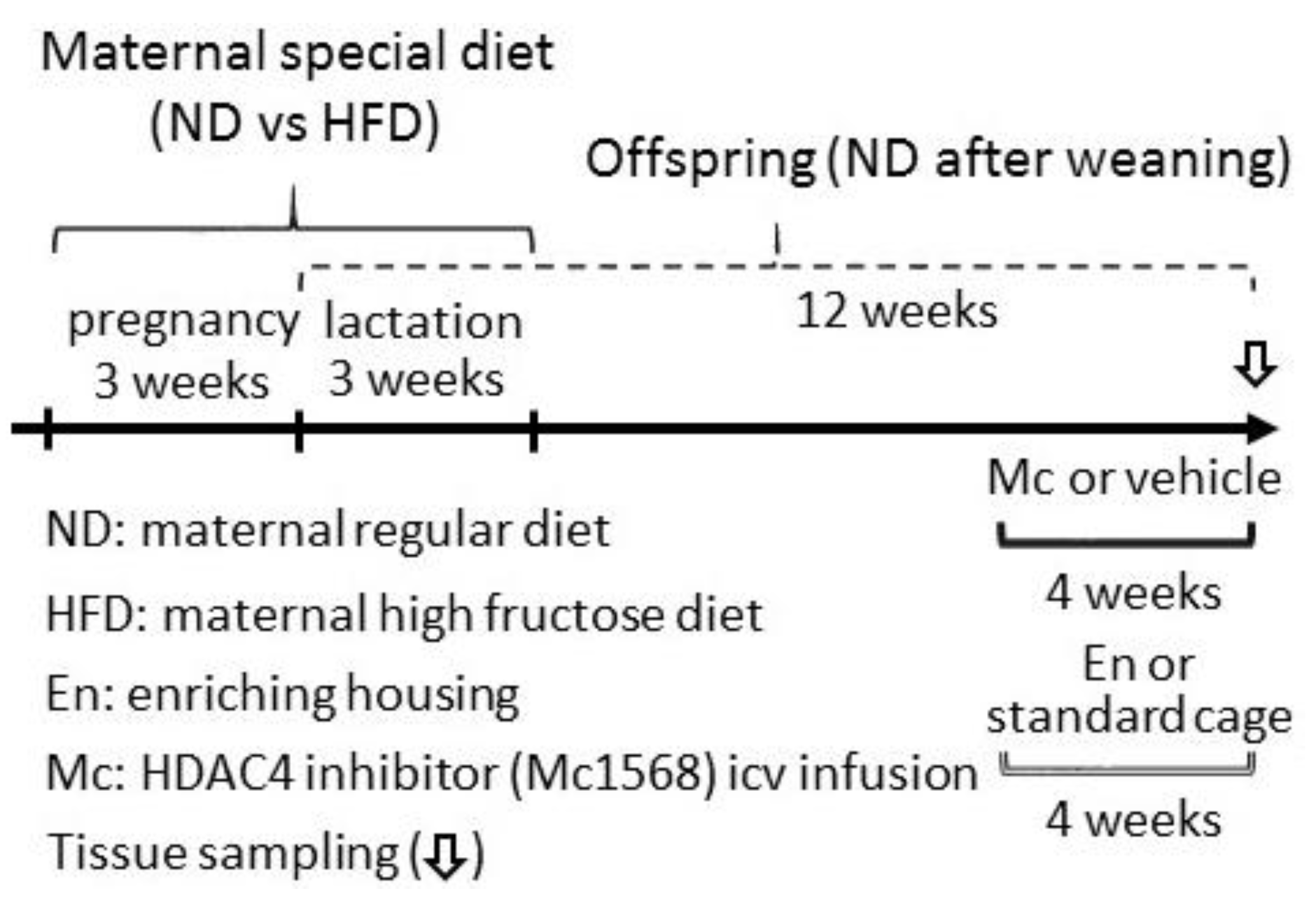
2.1. Animals
2.2. Enriched Housing
2.3. Intracerebroventricular (icv) Infusion
2.4. Total Protein Isolation
2.5. Nuclear Protein Extraction
2.6. Histone Deacetylase 4 Activity Assay
2.7. Western Blot Analysis
2.8. Brain Tissue Processing and Immunohistochemistry Labeling
2.9. Statistical Analysis
3. Results
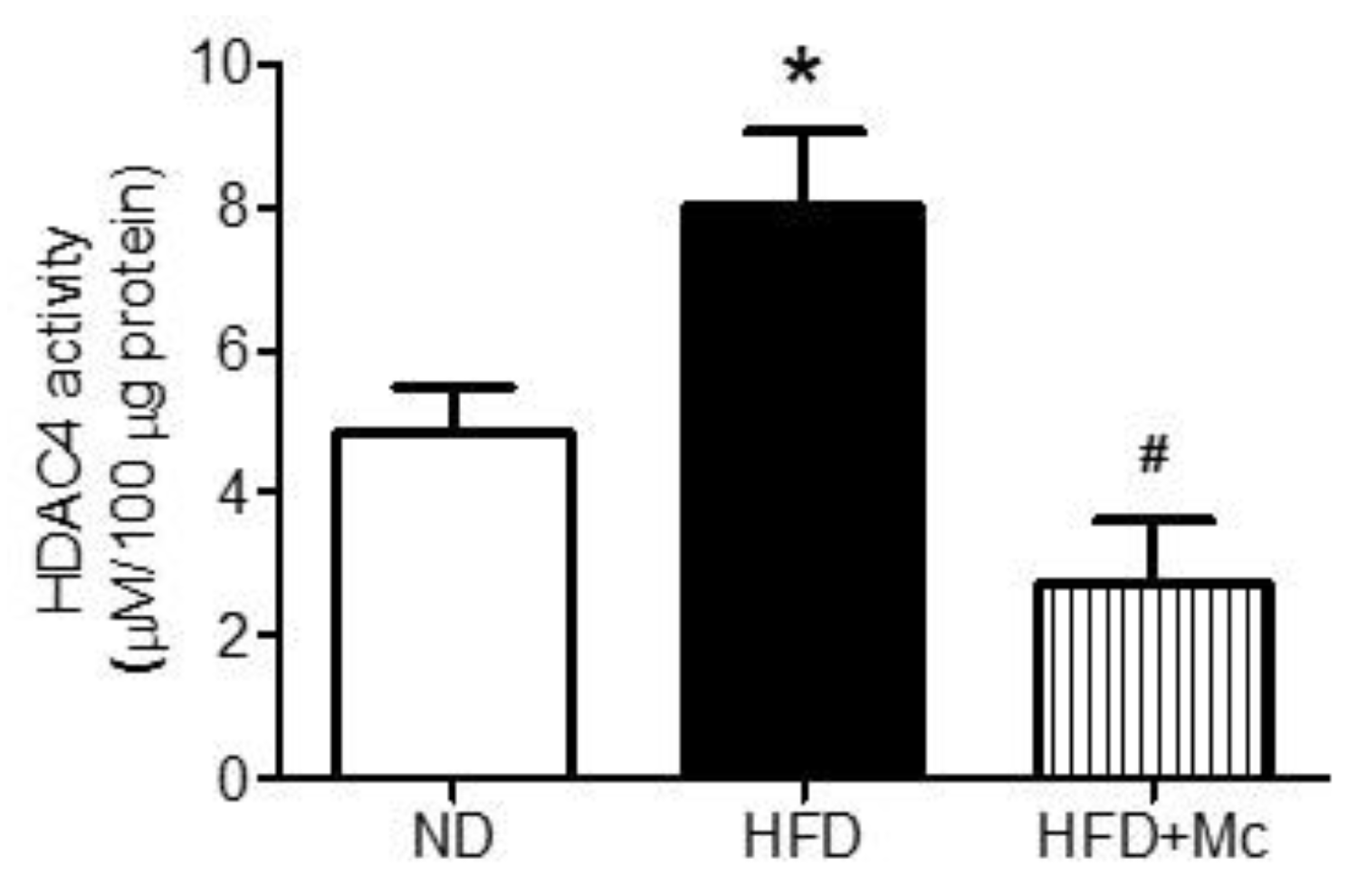
3.1. The HDAC4 Activity in the Hippocampus of Adult Female Offspring Was Increased by Maternal High-Fructose Diet
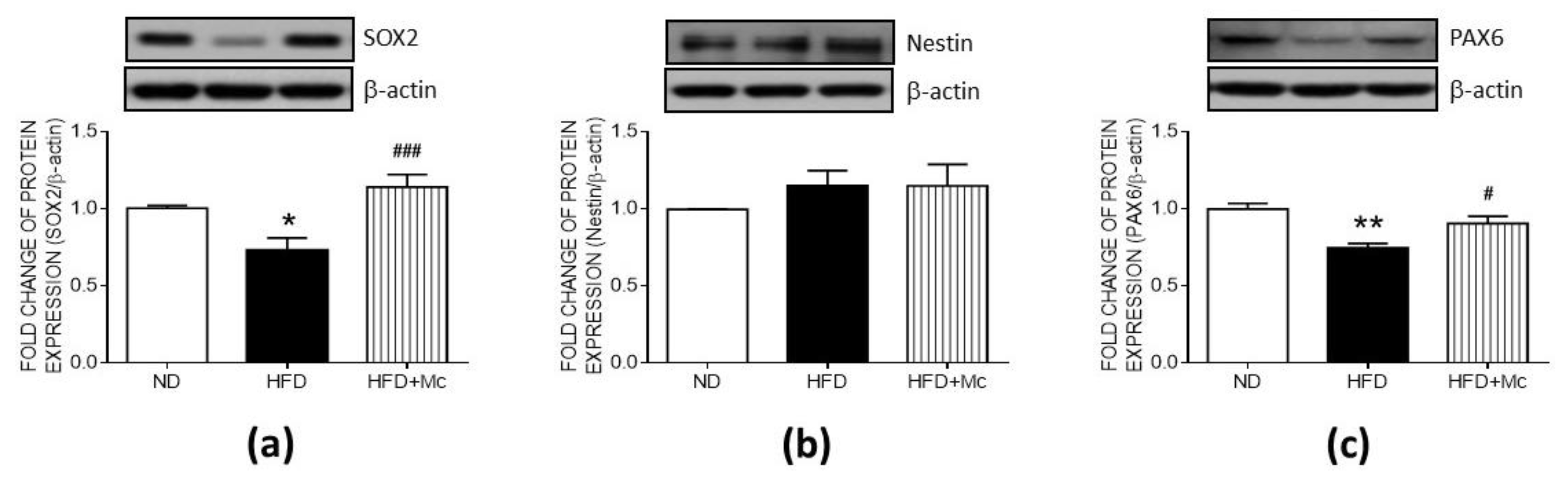
3.2. Inhibition of HDAC4 Reversed the Suppression Due to Maternal HFD as Assessed Using Markers for Hippocampal Neural Progenitor Cells
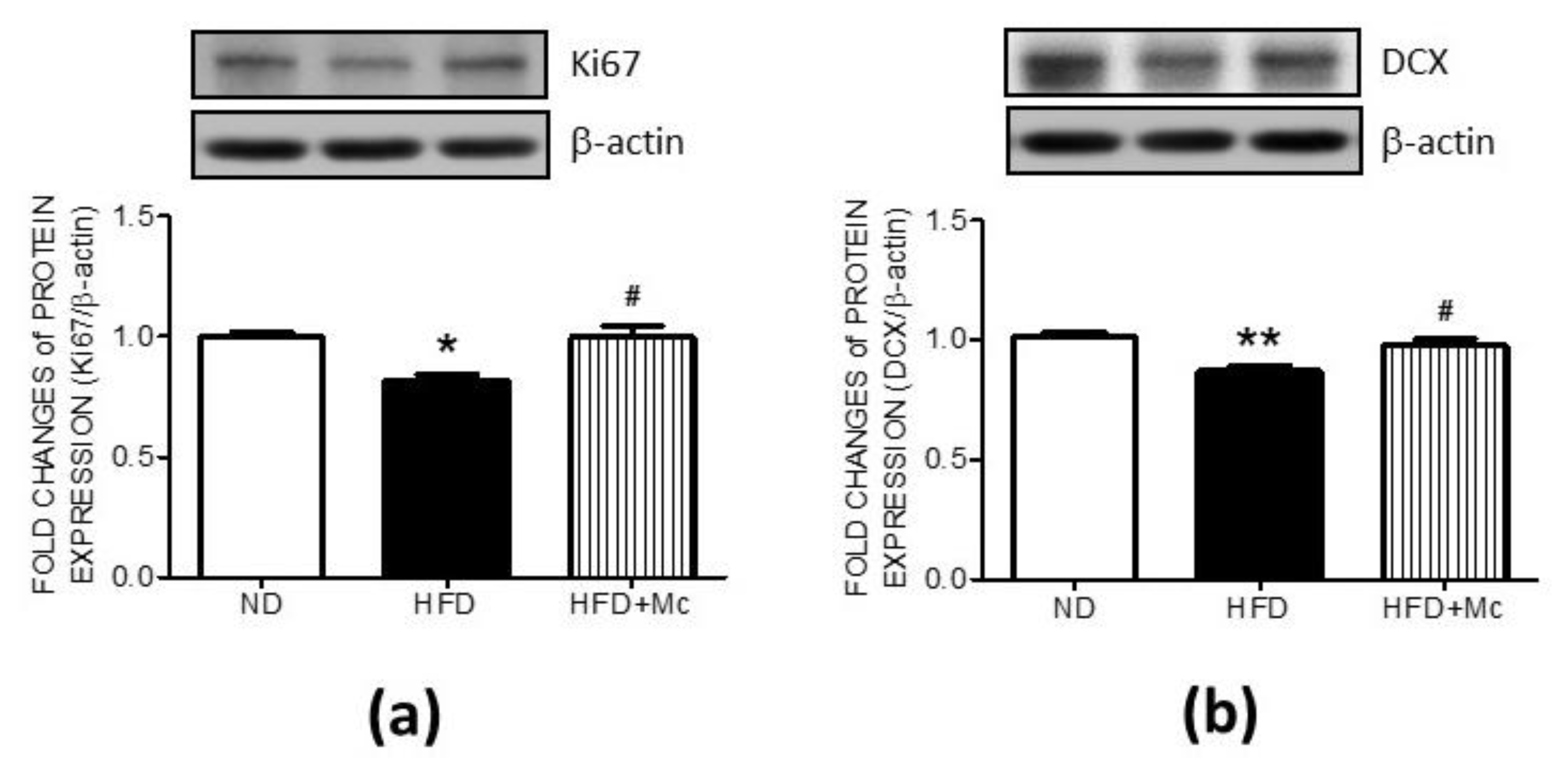
3.3. Inhibition of HDAC4 Reversed the Suppressed Markers of Cell Proliferation and Neuronal Differentiation in the Hippocampus of the HFD Group
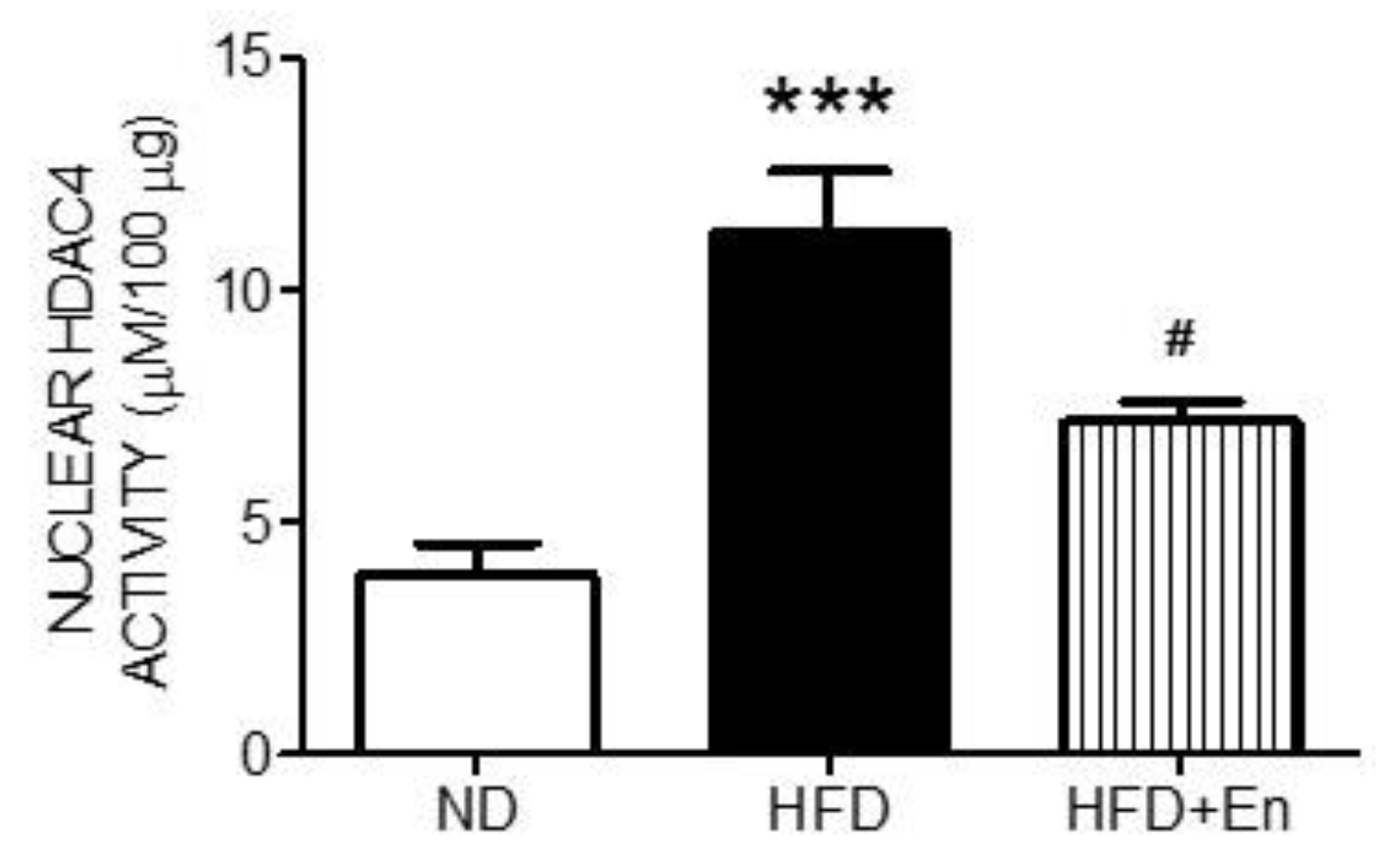
3.4. Enriched Housing Suppressed HDAC4 Activity in the Hippocampus of Adult Female Offspring Exposed to a Maternal High-Fructose Diet
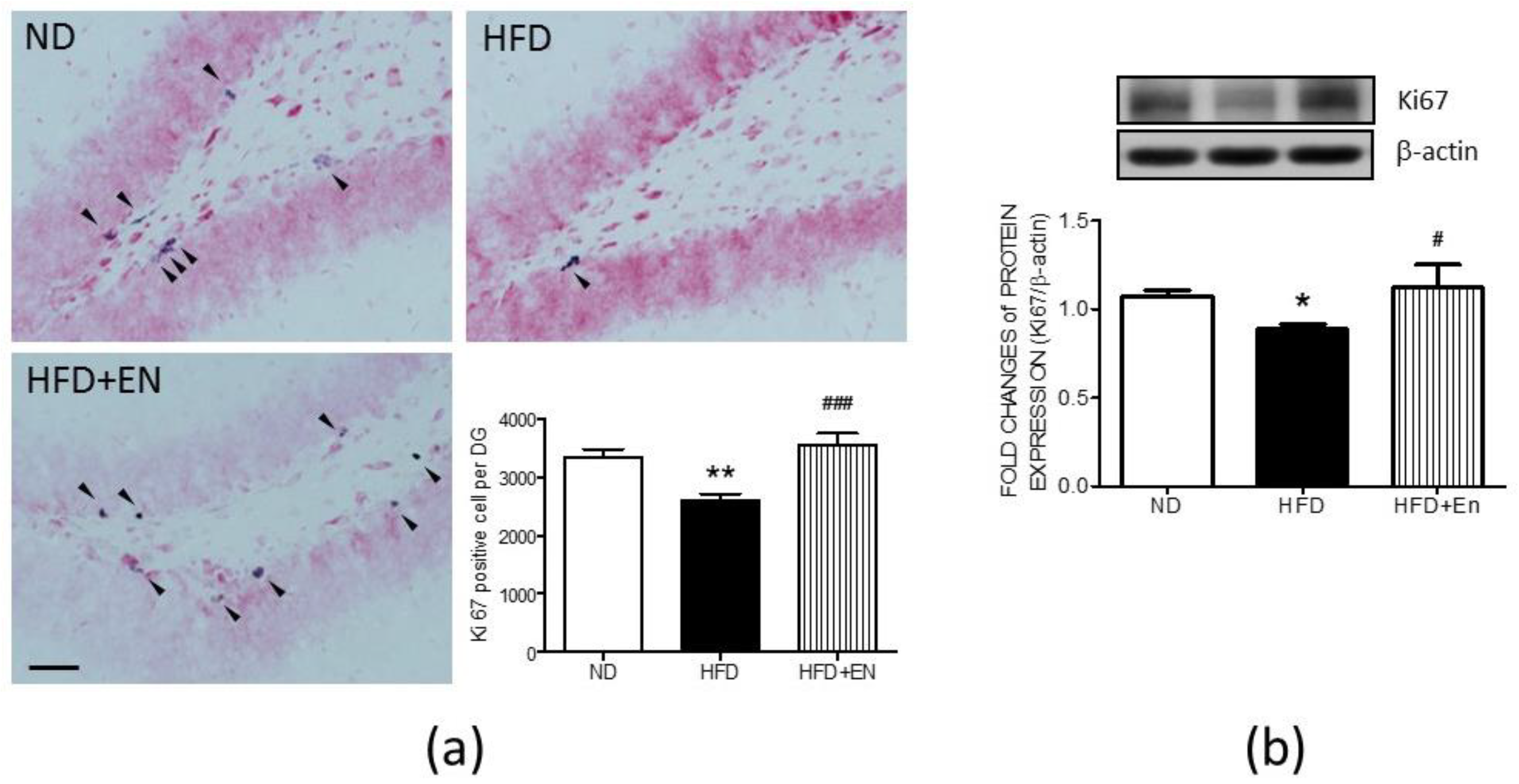
3.5. Enriched Housing Reversed the Expression of Hippocampal Ki67 in Adult Female Offspring Suppressed by Maternal High Fructose
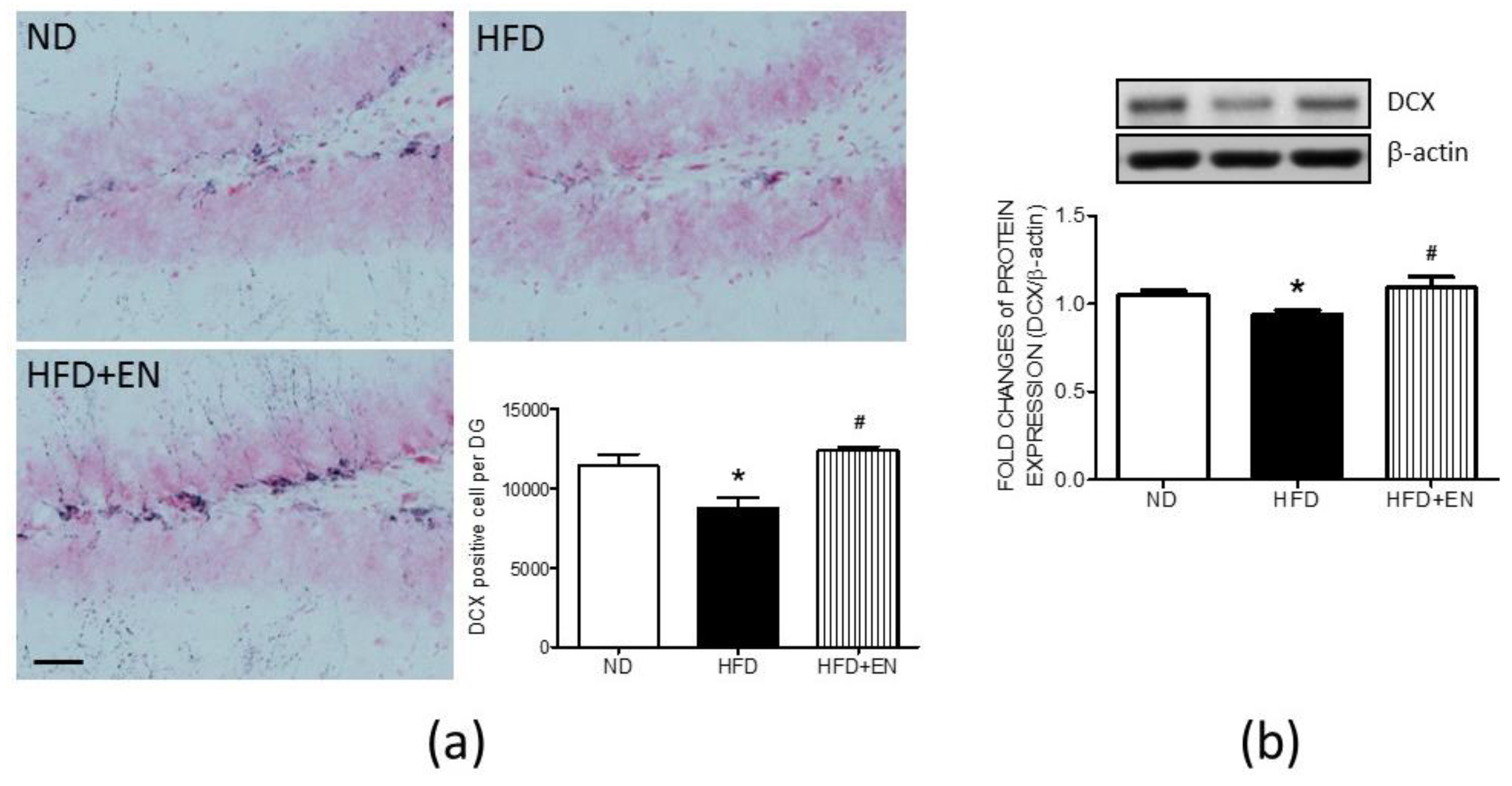
3.6. Enriched Housing Reversed the Suppression of Hippocampal DCX Related to Maternal High-Fructose Diet in Adult Female Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C. Developmental programming of health and disease. Proc. Nutr. Soc. 2006, 65, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Narain, A.; Kwok, C.S.; Mamas, M.A. Soft drinks and sweetened beverages and the risk of cardiovascular disease and mortality: A systematic review and meta-analysis. Int. J. Clin. Pract. 2016, 70, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, A.C.; Adeli, K. Fructose and the metabolic syndrome: Pathophysiology and molecular mechanisms. Nutr. Rev. 2007, 65 Pt 2, S13–S23. [Google Scholar] [CrossRef]
- Cohen, J.F.W.; Rifas-Shiman, S.L.; Young, J.; Oken, E. Associations of Prenatal and Child Sugar Intake With Child Cognition. Am. J. Prev. Med. 2018, 54, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.L.; Wu, C.W.; Tain, Y.L.; Huang, L.T.; Chao, Y.M.; Hung, C.Y.; Wu, J.C.; Chen, S.R.; Tsai, P.C.; Chan, J.Y. Environmental stimulation rescues maternal high fructose intake-impaired learning and memory in female offspring: Its correlation with redistribution of histone deacetylase 4. Neurobiol. Learn. Mem. 2016, 130, 105–117. [Google Scholar] [CrossRef]
- Saad, A.F.; Alshehri, W.; Lei, J.; Kechichian, T.B.; Gamble, P.; Alhejaily, N.; Shabi, Y.; Saade, G.R.; Costantine, M.M.; Burd, I. Maternal Fructose Consumption Disrupts Brain Development of Offspring in a Murine Model of Autism Spectrum Disorder. Am. J. Perinatol. 2016, 33, 1357–1364. [Google Scholar] [CrossRef]
- Eriksson, P.S.; Perfilieva, E.; Bjork-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Goncalves, J.T.; Schafer, S.T.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 2016, 167, 897–914. [Google Scholar] [CrossRef] [Green Version]
- Tronel, S.; Belnoue, L.; Grosjean, N.; Revest, J.M.; Piazza, P.V.; Koehl, M.; Abrous, D.N. Adult-born neurons are necessary for extended contextual discrimination. Hippocampus 2012, 22, 292–298. [Google Scholar] [CrossRef]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Spalding, K.L.; Bhardwaj, R.D.; Buchholz, B.A.; Druid, H.; Frisen, J. Retrospective birth dating of cells in humans. Cell 2005, 122, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the Adult Hippocampus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018812. [Google Scholar] [CrossRef] [Green Version]
- Gilyarov, A.V. Nestin in central nervous system cells. Neurosci. Behav. Physiol. 2008, 38, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Avilion, A.A.; Nicolis, S.K.; Pevny, L.H.; Perez, L.; Vivian, N.; Lovell-Badge, R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes. Dev. 2003, 17, 126–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferri, A.L.; Cavallaro, M.; Braida, D.; Di Cristofano, A.; Canta, A.; Vezzani, A.; Ottolenghi, S.; Pandolfi, P.P.; Sala, M.; DeBiasi, S.; et al. Sox2 deficiency causes neurodegeneration and impaired neurogenesis in the adult mouse brain. Development 2004, 131, 3805–3819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzino, A. Sox2 and Oct-3/4: A versatile pair of master regulators that orchestrate the self-renewal and pluripotency of embryonic stem cells. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Heins, N.; Malatesta, P.; Cecconi, F.; Nakafuku, M.; Tucker, K.L.; Hack, M.A.; Chapouton, P.; Barde, Y.A.; Gotz, M. Glial cells generate neurons: The role of the transcription factor Pax6. Nat. Neurosci. 2002, 5, 308–315. [Google Scholar] [CrossRef]
- Brown, J.P.; Couillard-Despres, S.; Cooper-Kuhn, C.M.; Winkler, J.; Aigner, L.; Kuhn, H.G. Transient expression of doublecortin during adult neurogenesis. J. Comp. Neurol. 2003, 467, 1–10. [Google Scholar] [CrossRef]
- Gage, F.H.; Kempermann, G.; Palmer, T.D.; Peterson, D.A.; Ray, J. Multipotent progenitor cells in the adult dentate gyrus. J. Neurobiol. 1998, 36, 249–266. [Google Scholar] [CrossRef]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [Green Version]
- Saxe, M.D.; Battaglia, F.; Wang, J.W.; Malleret, G.; David, D.J.; Monckton, J.E.; Garcia, A.D.; Sofroniew, M.V.; Kandel, E.R.; Santarelli, L.; et al. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc. Natl. Acad. Sci. USA 2006, 103, 17501–17506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, L.M.; Alexander, A.S.; Aimone, J.B.; Wiles, J.; Gage, F.H.; Chiba, A.A.; Quinn, L.K. Temporally selective contextual encoding in the dentate gyrus of the hippocampus. Nat. Commun. 2014, 5, 3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clelland, C.D.; Choi, M.; Romberg, C.; Clemenson, G.D., Jr.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L.M.; Barker, R.A.; Gage, F.H.; et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009, 325, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Nakashiba, T.; Cushman, J.D.; Pelkey, K.A.; Renaudineau, S.; Buhl, D.L.; McHugh, T.J.; Rodriguez Barrera, V.; Chittajallu, R.; Iwamoto, K.S.; McBain, C.J.; et al. Young dentate granule cells mediate pattern separation, whereas old granule cells facilitate pattern completion. Cell 2012, 149, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Wu, C.W.; Tain, Y.L.; Fu, M.H.; Hung, C.Y.; Chen, I.C.; Chen, L.W.; Wu, K.L.H. Oral pioglitazone ameliorates fructose-induced peripheral insulin resistance and hippocampal gliosis but not restores inhibited hippocampal adult neurogenesis. Biochim. Biophys. Acta 2018, 1864, 274–285. [Google Scholar] [CrossRef]
- Yang, X.J.; Seto, E. HATs and HDACs: From structure, function and regulation to novel strategies for therapy and prevention. Oncogene 2007, 26, 5310–5318. [Google Scholar] [CrossRef]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef]
- Wang, W.H.; Cheng, L.C.; Pan, F.Y.; Xue, B.; Wang, D.Y.; Chen, Z.; Li, C.J. Intracellular trafficking of histone deacetylase 4 regulates long-term memory formation. Anat. Rec. (Hoboken) 2011, 294, 1025–1034. [Google Scholar] [CrossRef]
- Scharfman, H.; Goodman, J.; Macleod, A.; Phani, S.; Antonelli, C.; Croll, S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp. Neurol. 2005, 192, 348–356. [Google Scholar] [CrossRef]
- Yao, B.; Christian, K.M.; He, C.; Jin, P.; Ming, G.L.; Song, H. Epigenetic mechanisms in neurogenesis. Nat. Rev. Neurosci. 2016, 17, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Speisman, R.B.; Kumar, A.; Rani, A.; Pastoriza, J.M.; Severance, J.E.; Foster, T.C.; Ormerod, B.K. Environmental enrichment restores neurogenesis and rapid acquisition in aged rats. Neurobiol. Aging 2013, 34, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H. More hippocampal neurons in adult mice living in an enriched environment. Nature 1997, 386, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Novkovic, T.; Mittmann, T.; Manahan-Vaughan, D. BDNF contributes to the facilitation of hippocampal synaptic plasticity and learning enabled by environmental enrichment. Hippocampus 2015, 25, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rampon, C.; Tang, Y.P.; Goodhouse, J.; Shimizu, E.; Kyin, M.; Tsien, J.Z. Enrichment induces structural changes and recovery from nonspatial memory deficits in CA1 NMDAR1-knockout mice. Nat. Neurosci. 2000, 3, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.P.; Wang, H.; Feng, R.; Kyin, M.; Tsien, J.Z. Differential effects of enrichment on learning and memory function in NR2B transgenic mice. Neuropharmacology 2001, 41, 779–790. [Google Scholar] [CrossRef]
- Lee, E.H.; Hsu, W.L.; Ma, Y.L.; Lee, P.J.; Chao, C.C. Enrichment enhances the expression of sgk, a glucocorticoid-induced gene, and facilitates spatial learning through glutamate AMPA receptor mediation. Eur. J. Neurosci. 2003, 18, 2842–2852. [Google Scholar] [CrossRef]
- Fischer, A.; Sananbenesi, F.; Wang, X.; Dobbin, M.; Tsai, L.H. Recovery of learning and memory is associated with chromatin remodelling. Nature 2007, 447, 178–182. [Google Scholar] [CrossRef]
- Koppel, I.; Timmusk, T. Differential regulation of Bdnf expression in cortical neurons by class-selective histone deacetylase inhibitors. Neuropharmacology 2013, 75, 106–115. [Google Scholar] [CrossRef]
- Tozuka, Y.; Kumon, M.; Wada, E.; Onodera, M.; Mochizuki, H.; Wada, K. Maternal obesity impairs hippocampal BDNF production and spatial learning performance in young mouse offspring. Neurochem. Int. 2010, 57, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Deng, S.; Li, W.G.; Yu, Y.; Li, F.; Mao, M. Maternal obesity caused by overnutrition exposure leads to reversal learning deficits and striatal disturbance in rats. PLoS ONE 2013, 8, e78876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, K.C.; Jones, E.K.; Anday, E.K. Maternal and postweaning high-fat diets disturb hippocampal gene expression, learning, and memory function. American journal of physiology. Regul. Integr. Comp. Physiol. 2014, 306, R527–R537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeijmakers, L.; Lucassen, P.J.; Korosi, A. The interplay of early-life stress, nutrition, and immune activation programs adult hippocampal structure and function. Front. Mol. Neurosci. 2014, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Alejandre-Gomez, M.; Garcia-Segura, L.M.; Gonzalez-Burgos, I. Administration of an inhibitor of estrogen biosynthesis facilitates working memory acquisition in male rats. Neurosci. Res. 2007, 58, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Lopez, M.; Mendez, M.; Lopez, L.; Arias, J.L. Spatial working memory in Wistar rats: Brain sex differences in metabolic activity. Brain. Res. Bull. 2009, 79, 187–192. [Google Scholar] [CrossRef]
- Lillycrop, K.A.; Burdge, G.C. Maternal diet as a modifier of offspring epigenetics. J. Dev. Orig. Health Dis. 2015, 6, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Huang, M.; Bushong, E.; Phan, S.; Uytiepo, M.; Beutter, E.; Boemer, D.; Tsui, K.; Ellisman, M.; Maximov, A. Class IIa HDACs regulate learning and memory through dynamic experience-dependent repression of transcription. Nat. Commun. 2019, 10, 3469. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.P.; Denicola, M.; Qin, X.; Zhao, Y.; Zhang, L.; Long, X.L.; Zhuang, S.; Liu, P.Y.; Zhao, T.C. HDAC inhibition promotes cardiogenesis and the survival of embryonic stem cells through proteasome-dependent pathway. J. Cell Biochem. 2011, 112, 3246–3255. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Qin, G.; Zhao, T.C. HDAC4: Mechanism of regulation and biological functions. Epigenomics 2014, 6, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Suh, H.; Consiglio, A.; Ray, J.; Sawai, T.; D’Amour, K.A.; Gage, F.H. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2+ neural stem cells in the adult hippocampus. Cell Stem. Cell 2007, 1, 515–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packard, A.I.; Lin, B.; Schwob, J.E. Sox2 and Pax6 Play Counteracting Roles in Regulating Neurogenesis within the Murine Olfactory Epithelium. PLoS ONE 2016, 11, e0155167. [Google Scholar] [CrossRef] [PubMed]
- Trazzi, S.; Fuchs, C.; Viggiano, R.; De Franceschi, M.; Valli, E.; Jedynak, P.; Hansen, F.K.; Perini, G.; Rimondini, R.; Kurz, T.; et al. HDAC4: A key factor underlying brain developmental alterations in CDKL5 disorder. Hum. Mol. Genet. 2016, 25, 3887–3907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruel-Jungerman, E.; Laroche, S.; Rampon, C. New neurons in the dentate gyrus are involved in the expression of enhanced long-term memory following environmental enrichment. Eur. J. Neurosci. 2005, 21, 513–521. [Google Scholar] [CrossRef]
- Falkenberg, T.; Mohammed, A.K.; Henriksson, B.; Persson, H.; Winblad, B.; Lindefors, N. Increased expression of brain-derived neurotrophic factor mRNA in rat hippocampus is associated with improved spatial memory and enriched environment. Neurosci. Lett. 1992, 138, 153–156. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g/kg | Regular Chow (ND) | 60% High-Fructose Diet (HFD) |
|---|---|---|
| Fructose | - | 600 |
| Lard | 50 | 50.0 |
| Casein | 232.3 | 207.0 |
| Cellulose | 51 | 79.81 |
| DL-Methionine | 6.7 | 3.0 |
| Mineral Mix Rogers-Harper (170760) | >7 | 50.0 |
| Zinc Carbonate | 0.04 | |
| Vitamin Mix Teklad (40060) | 10.0 | |
| Food Color (Green) | - | 0.15 |
| % kcal from | ||
| Carbohydrate | 57.996 | 66.8 |
| Fat | 13.496 | 13.0 |
| Protein | 28.507 | 20.2 |
| kcal/g | 3.35 | 3.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-C.; Wu, C.-W.; Hung, P.-L.; Chan, J.Y.H.; Tain, Y.-L.; Fu, M.-H.; Chen, L.-W.; Liang, C.-K.; Hung, C.-Y.; Yu, H.-R.; et al. Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4. Int. J. Environ. Res. Public Health 2020, 17, 3919. https://doi.org/10.3390/ijerph17113919
Liu W-C, Wu C-W, Hung P-L, Chan JYH, Tain Y-L, Fu M-H, Chen L-W, Liang C-K, Hung C-Y, Yu H-R, et al. Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4. International Journal of Environmental Research and Public Health. 2020; 17(11):3919. https://doi.org/10.3390/ijerph17113919
Chicago/Turabian StyleLiu, Wen-Chung, Chih-Wei Wu, Pi-Lien Hung, Julie Y. H. Chan, You-Lin Tain, Mu-Hui Fu, Lee-Wei Chen, Chih-Kuang Liang, Chun-Ying Hung, Hong-Ren Yu, and et al. 2020. "Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4" International Journal of Environmental Research and Public Health 17, no. 11: 3919. https://doi.org/10.3390/ijerph17113919
APA StyleLiu, W. -C., Wu, C. -W., Hung, P. -L., Chan, J. Y. H., Tain, Y. -L., Fu, M. -H., Chen, L. -W., Liang, C. -K., Hung, C. -Y., Yu, H. -R., Chen, I. -C., & Wu, K. L. H. (2020). Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4. International Journal of Environmental Research and Public Health, 17(11), 3919. https://doi.org/10.3390/ijerph17113919