Inexpensive Home Infrared Living/Environment Sensor with Regional Thermal Information for Infant Physical and Psychological Development


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
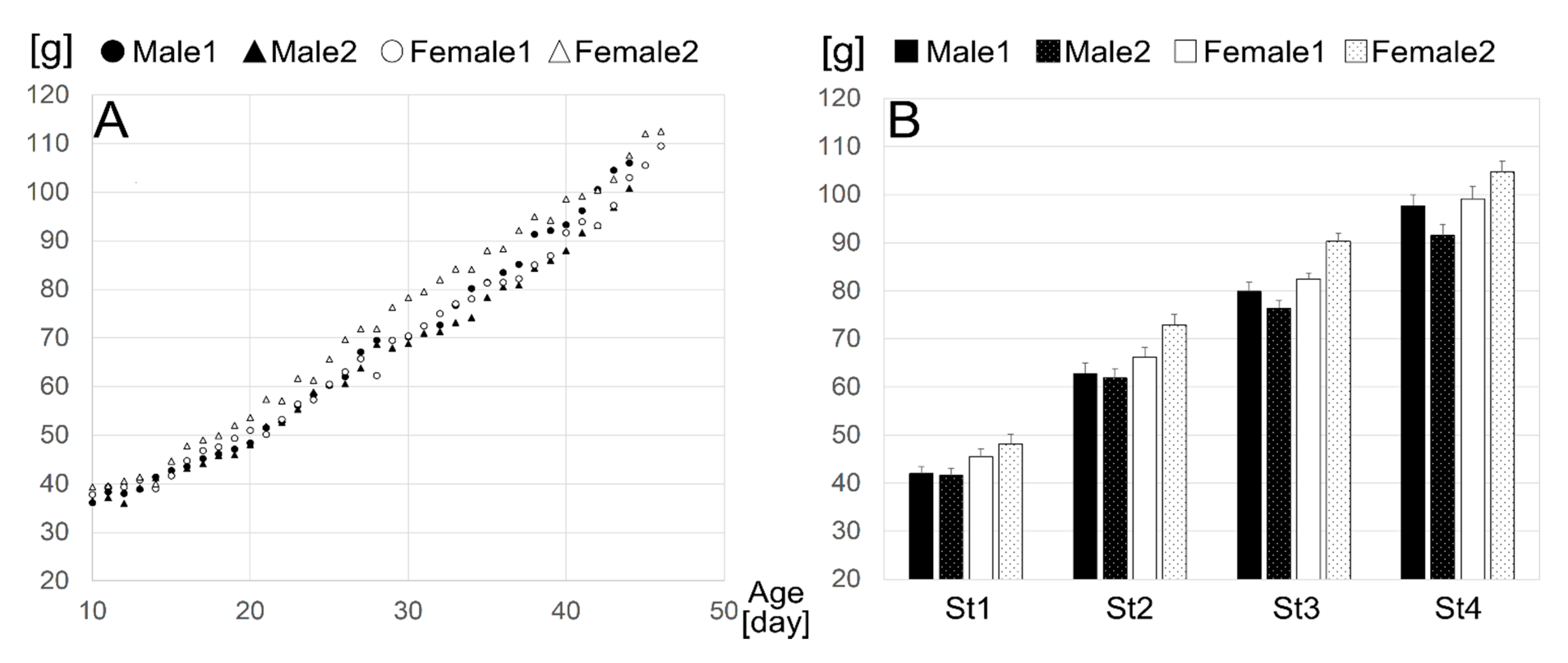
2.1. Animals
2.2. Image Processing and Data Acquisition
- S1 < Sbase because the area of body size and the detected surface temperature increased independently of the actual surface temperature when the marmoset approached the camera.
- NS1 > 0.6 to discriminate a marmoset from other objects based on their shapes, because the shape of a marmoset in the image was like a circle or ellipse (Figure 1), whereas those of other heat-generating elements were diagonally slender.
- Rmax > 0.3 because the correlation coefficient became low when the pattern of the background was flat.
- VHratio > 1/3 and <3 to distinguish a marmoset from a slender object, such as heat-generating elements.
- MAX > 1.5 because when the temperature difference between a marmoset and the surroundings was too small, a false position was incorrectly detected as the marmoset’s position.
- SD < 1.5 because when a marmoset approached the thermal camera too closely, the standard deviation became high due to the detection of increased temperature of the marmoset compared to the surrounding.
- Correction of BV. For BV, the difference between the distance in pixels and the actual distance depends on the distance between the camera and the marmoset. To normalize BV for changing height size, BV determined in the smaller height (up to P31) was multiplied by 320/700 (the height (mm) of the IR camera).
- Correction of BT. We found preliminarily that the raw BT (BTraw) detected by the IR thermal camera fluctuated depending on the temperature inside the cage (IT). To remove the increment or decrement of the IT-dependent BTraw signals, the model formula below was used, and BTraw was normalized individually:BT: the corrected body-surface temperature, ITave: the mean IT during the whole measurement period, alpha: the slope from the regression of BTraw by IT.BT = BTraw − α * (IT − ITave)
2.3. Accuracy of the Developed Algorithm
2.4. Acquisition of Environmental Variables
2.5. Definition of Biological Door Preference (BD)
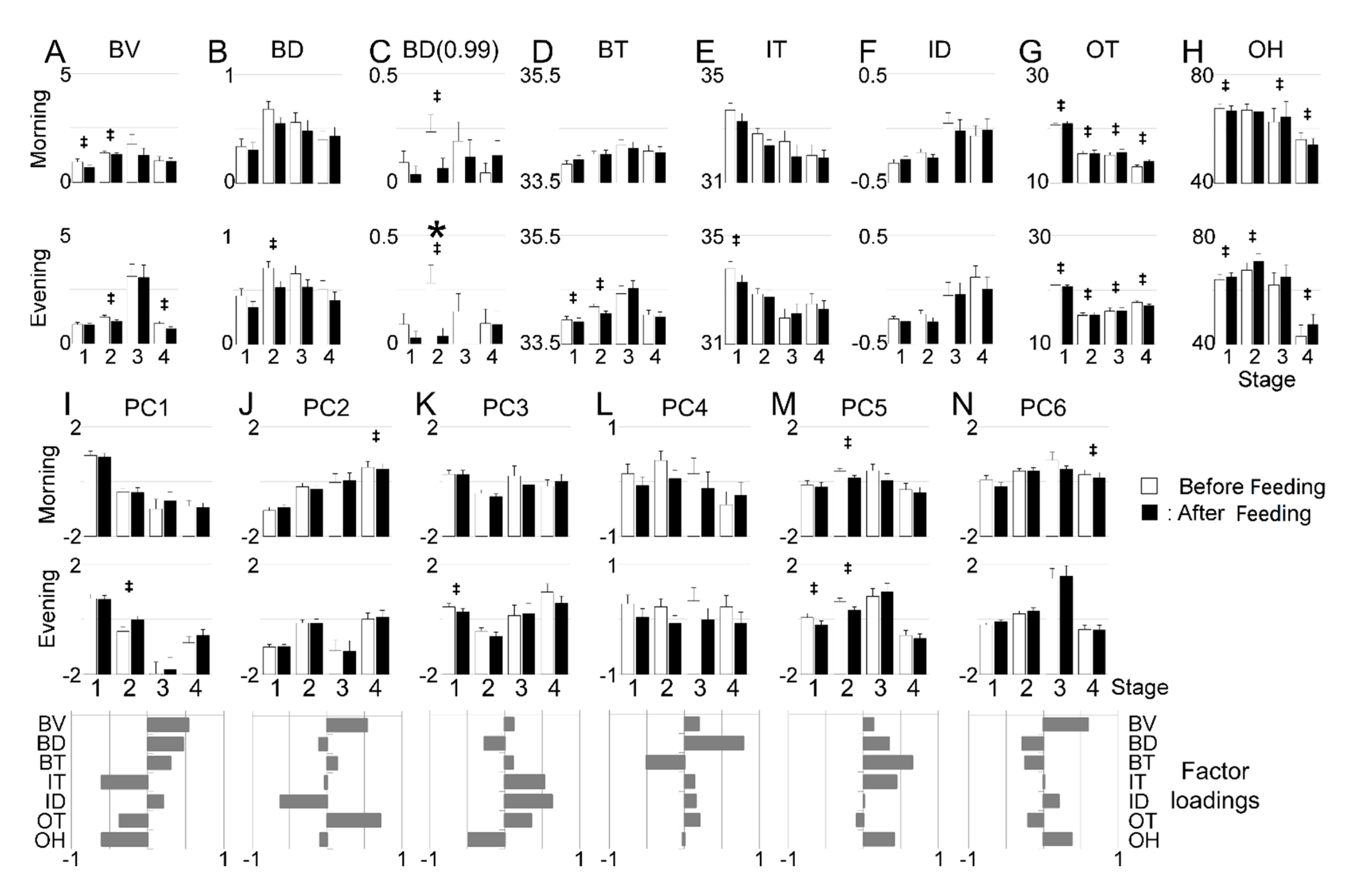
2.6. Data Extraction for ‘Feeding’-Related Analysis
2.7. Statistical Analyses
2.7.1. Comparison between Light/Dark Periods and before/after Feeding
2.7.2. Feeding-Dependent Explanatory/Response Variables
| lmer(R~E1+(E1|ID)) |
| lmer(R~E1+E2+(E1+E2|ID)) |
| lmer(R~E1+E2+E3+(E1+E2+E3|ID)) |
| lmer(R~E1+E2+E3+E4+(E1+E2+E3+E4|ID)) |
3. Results and Discussion
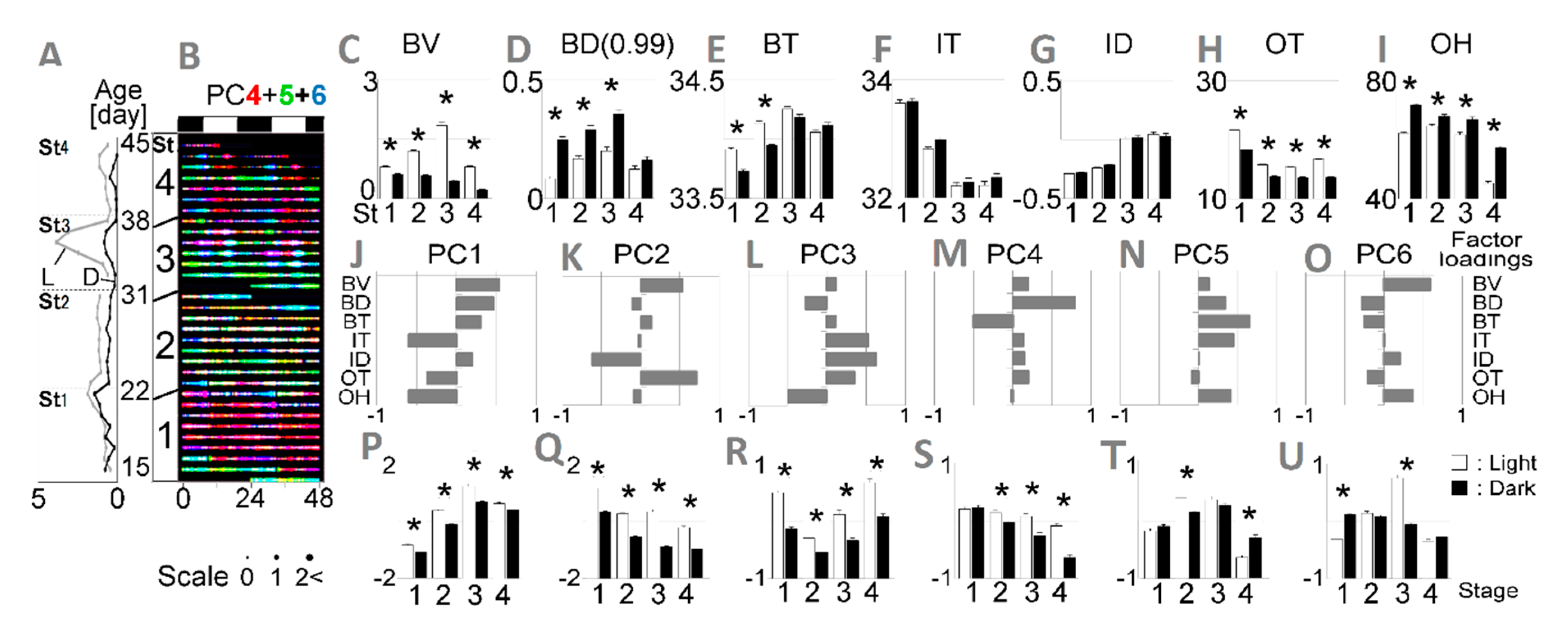
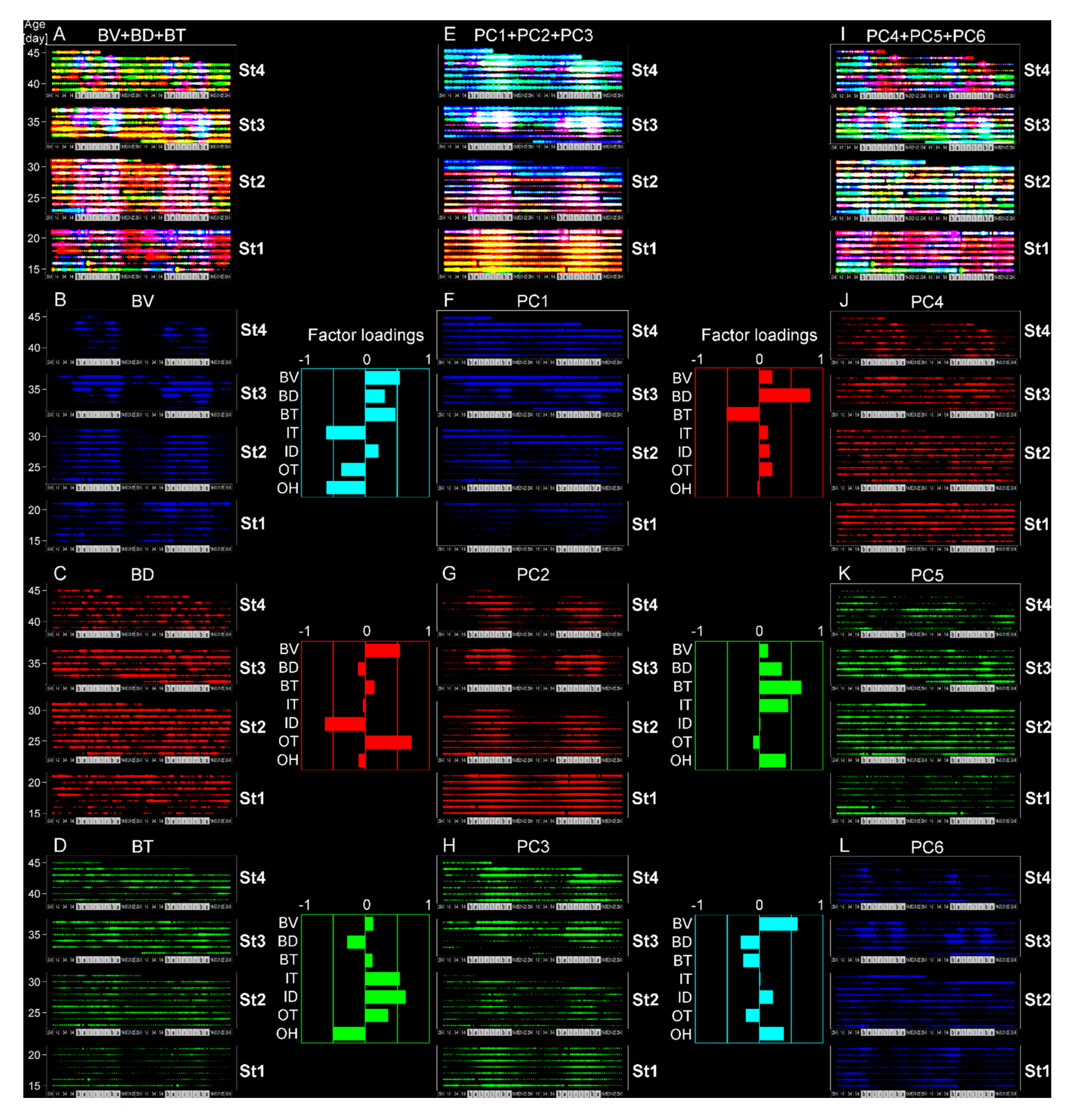
3.1. Definition of Age Stages
3.2. Testing of the Developed Algorithm through Evaluation of Age-Dependent Shifts in Circadian Rhythms
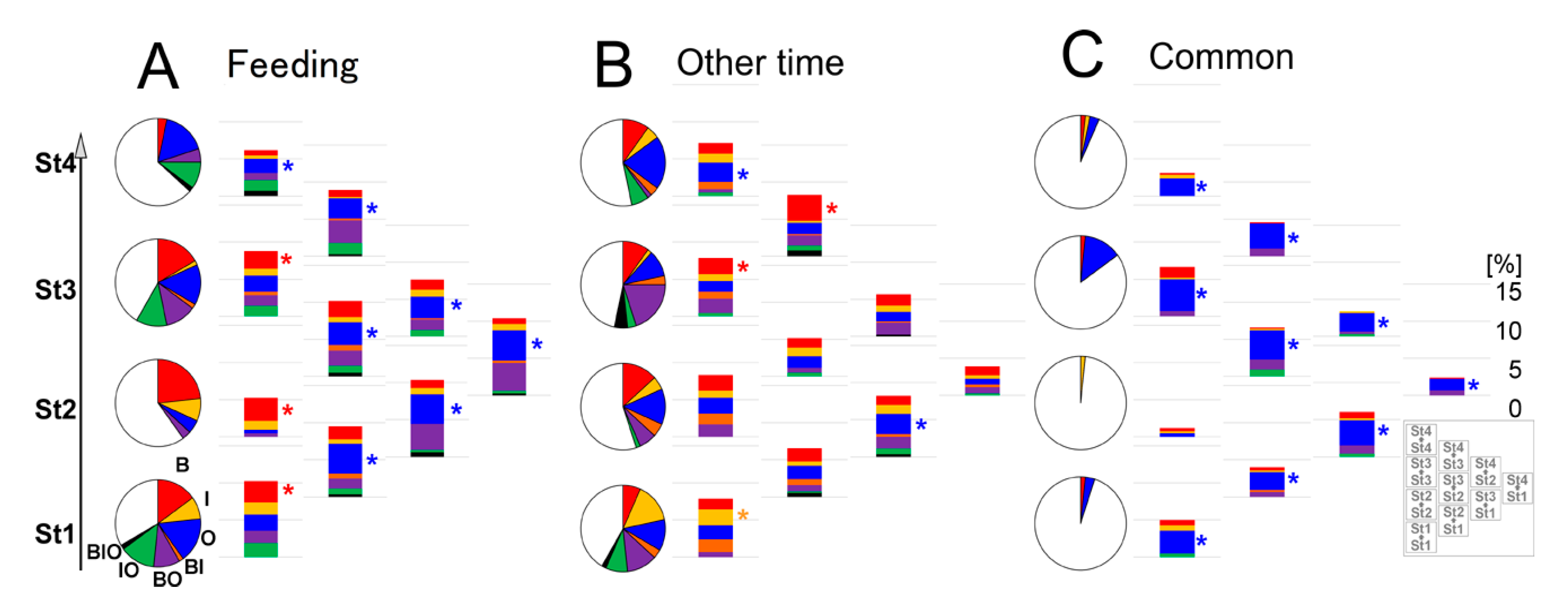
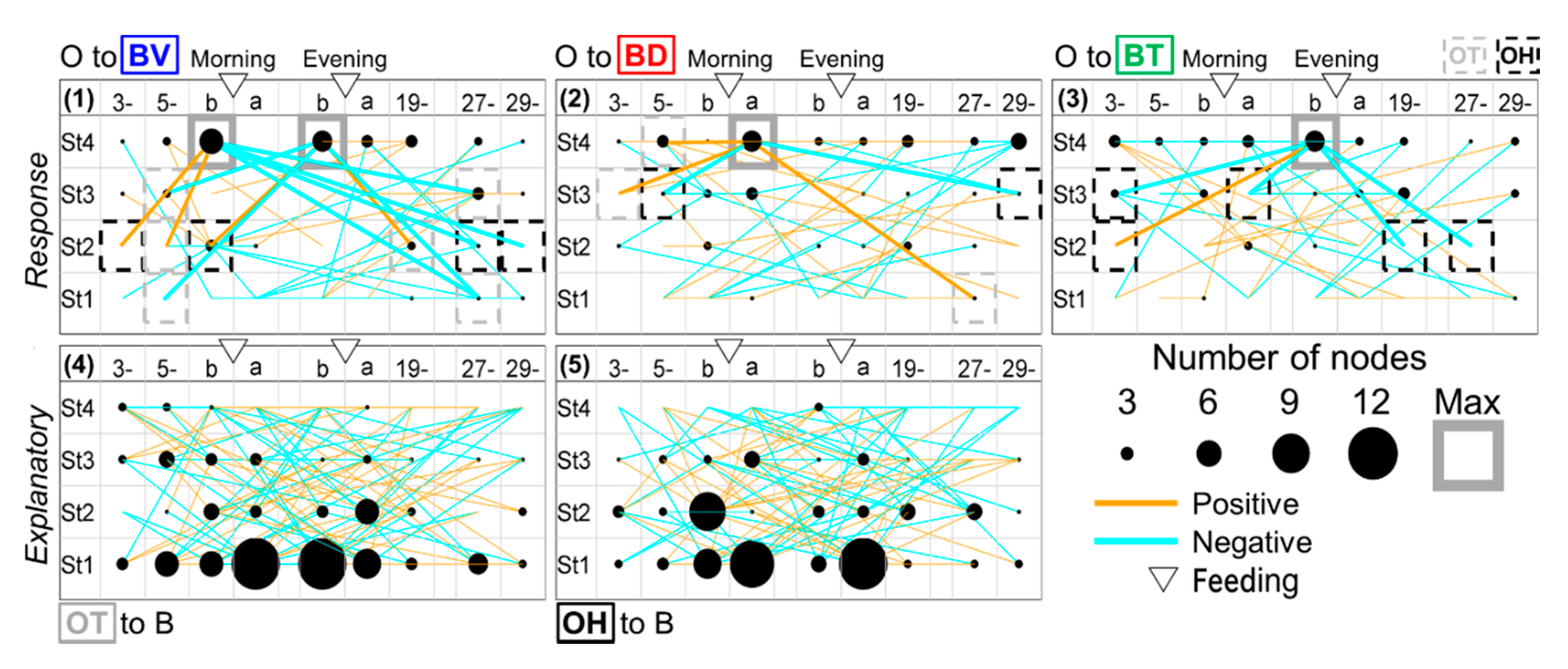
3.3. Estimation of Developmental Cause and Effect Based on Socially Interactive Feeding
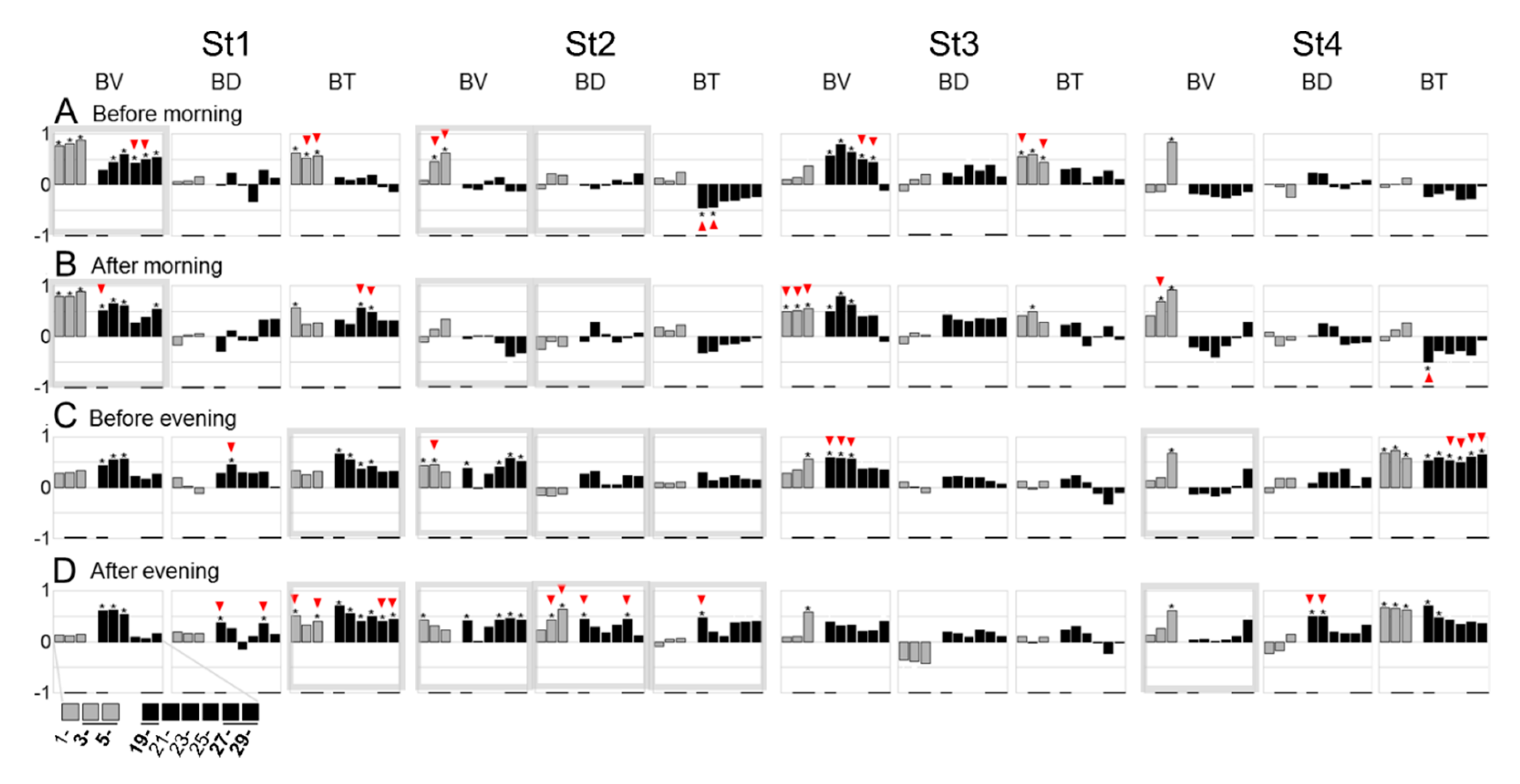
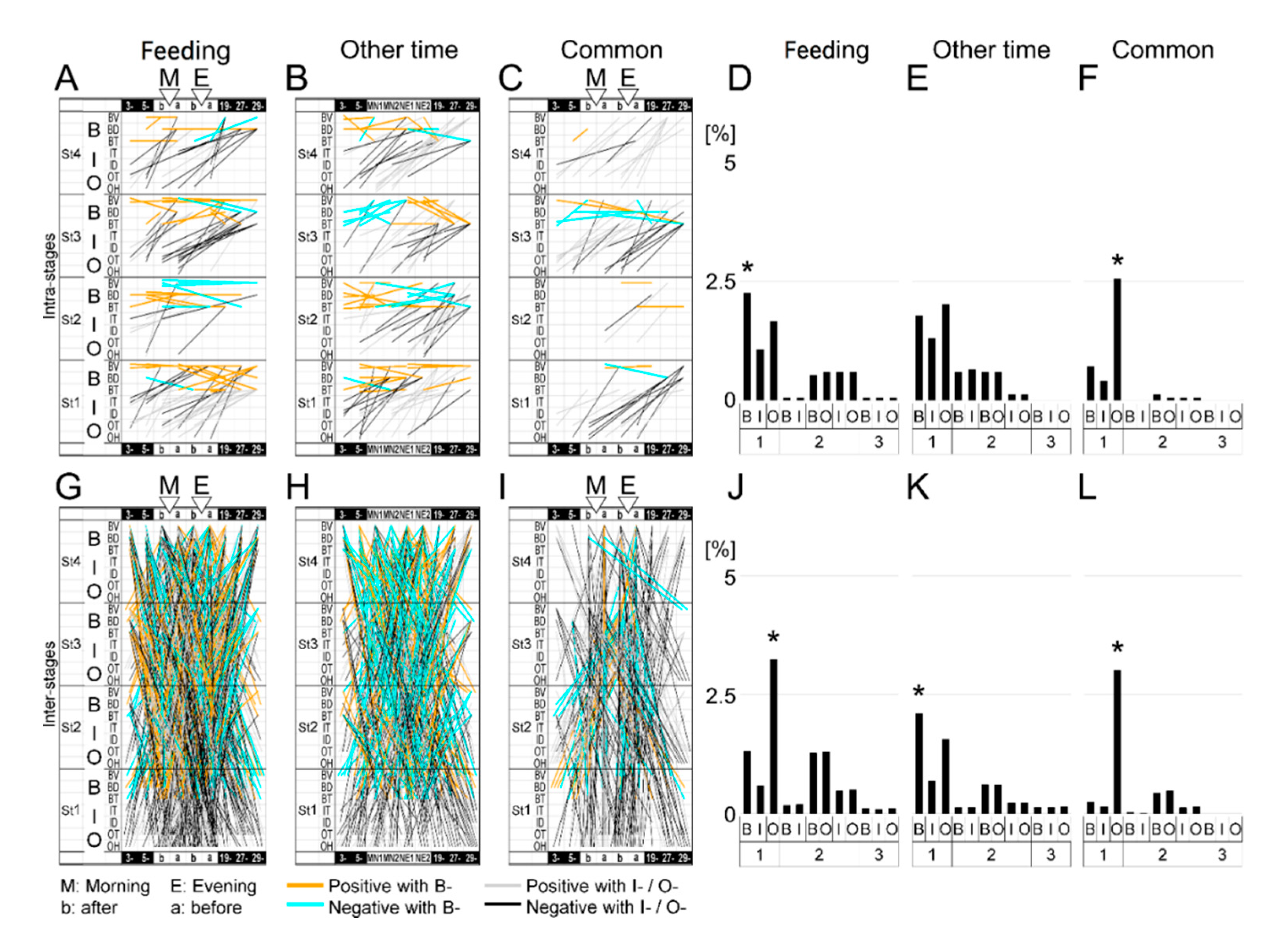
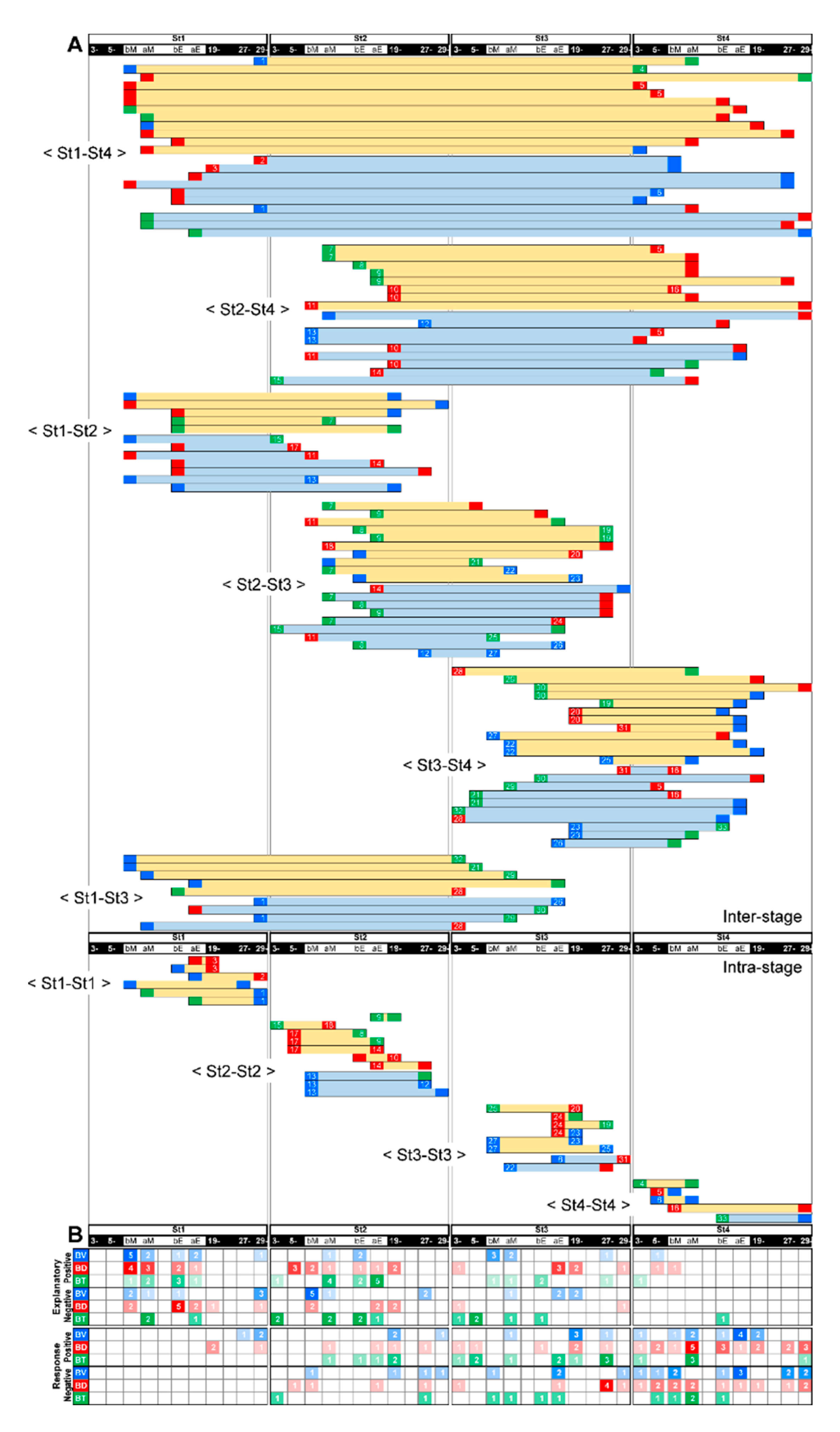
3.4. Feeding-Dependent B- Developmental Pathways of Intra- and Inter-Stages with Autonomic B- or Climatic O- Explanatory Variables
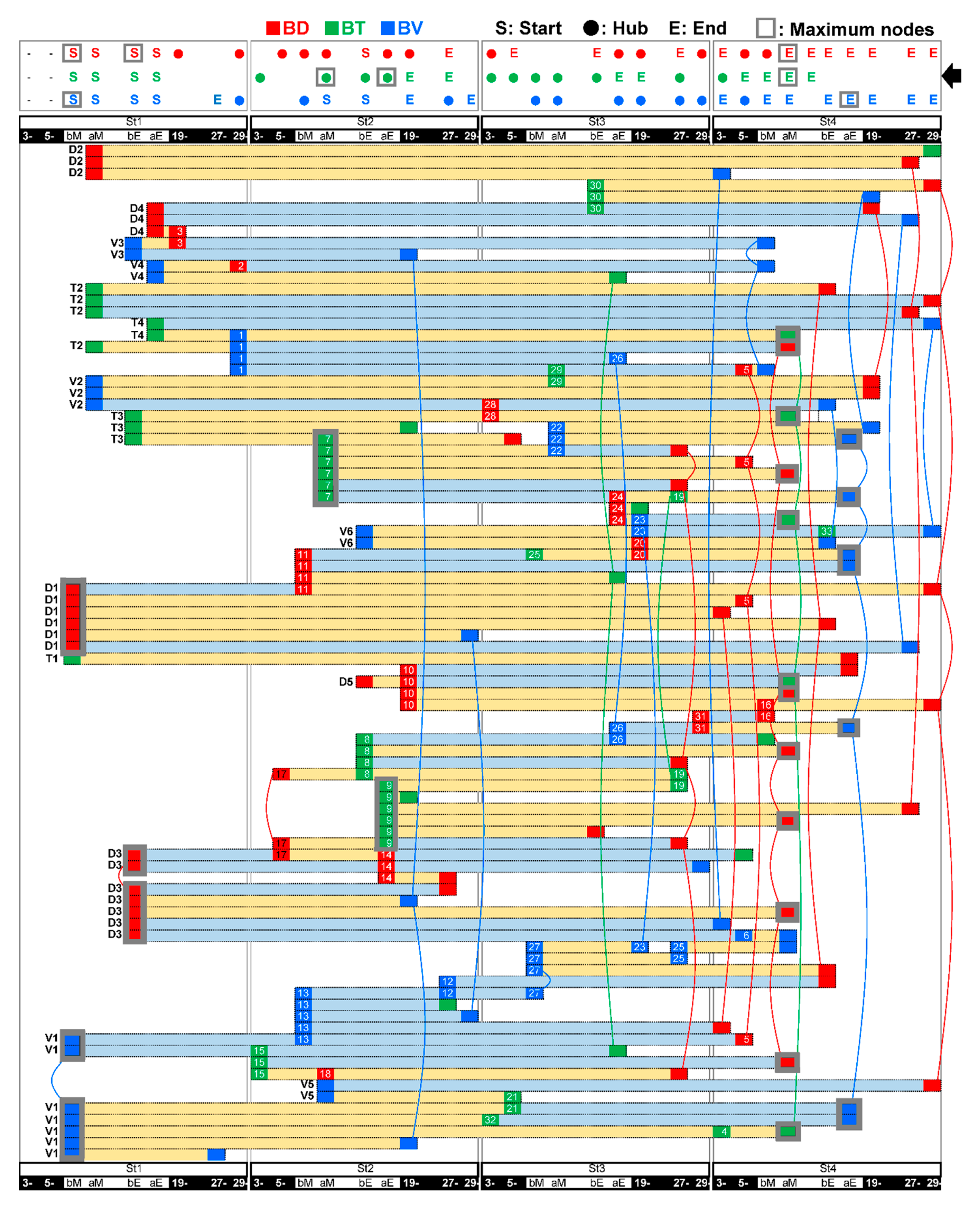
3.5. A Trigger Candidate of the Feeding-Dependent B-Developmental Pathway Contributes to Circadian Biological Door Preference-Body Surface Temperature (BD-BT) Correlation Switching
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jeon, E.S.; Choi, J.-S.; Lee, J.H.; Shin, K.Y.; Kim, Y.G.; Le, T.T.; Park, K.R. Human detection based on the generation of a background image by using a far-infrared light camera. Sensors 2015, 15, 6763–6788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzubaidi, A.; Leonhardt, S. Intelligent neonatal monitoring based on a virtual thermal sensor. BMC Med. Imaging 2014, 14, 9. [Google Scholar] [CrossRef]
- Shimizu, T.; Saga, T. The thermal imaging sensor with element of about 2000 pixels and applications. The Papers of Technical Meeting on Physical Sensor; IEE: Tokyo, Japan PHS 09-25. , 2009; pp. 45–49. Available online: https://ci.nii.ac.jp/naid/10027802584/ (accessed on 1 September 2020).
- Kido, S.; Miyasaka, T.; Tanaka, T.; Shimizu, T.; Saga, T. Fall detection in toilet rooms using thermal imaging sensors. In Proceedings of the 2009 IEEE/SICE International Symposium on System Integration, Tokyo, Japan, 29–29 January 2009. [Google Scholar] [CrossRef]
- Koshiba, M.; Seno, A.; Karino, G.; Shirakawa, Y.; Mimura, K.; Sagawa, T.; Tsugawa, W.; Sode, K.; Nakamura, S. Blood glucose dependence on emotional behaviors and body surface temperatures in common marmoset’s socio-psychological learning with peers—For ‘development of human-environment interface by sensing and multivariate analysis of bio-ecosystem’. ECS Trans. 2013, 50, 9–14. [Google Scholar] [CrossRef]
- Koshiba, M.; Nakamura, S.; Mimura, K.; Senoo, A.; Karino, G.; Amemiya, S.; Miyaji, T.; Kunikata, T.; Yamanouchi, H. Socio-emotional development evaluated by Behaviour Output analysis for Quantitative Emotional State Translation (BOUQUET): Towards early diagnosis of individuals with developmental disorders. OA Autism 2013, 1, 18. Available online: http://www.oapublishinglondon.com/images/article/pdf/1381009937.pdf (accessed on 1 July 2020). [CrossRef]
- Karino, G.; Shukuya, M.; Nakamura, S.; Murakoshi, T.; Kunikata, T.; Yamanouchi, H.; Koshiba, M. Common marmosets develop age-specific peer social experiences that may affect their adult body weight adaptation to climate. Stress Brain Behav. 2015, 3, 1–8. Available online: http://isbsjapan.kir.jp/wp-content/uploads/2019/08/e018001.pdf (accessed on 1 July 2020).
- Karino, G.; Murakoshi, T.; Nakamura, S.; Kunikata, T.; Yamanouchi, H.; Koshiba, M. Timing of changes from a primitive reflex to a voluntary behavior in infancy as a potential predictor of socio-psychological and physical development during juvenile stages among common marmosets. J. King Saud Univ. Sci. 2015, 27, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, Y.; Mimura, K.; Senoo, A.; Fujii, K.; Shimizu, T.; Saga, T.; Tanaka, I.; Honda, Y.; Tokuno, H.; Usui, S.; et al. Multivariate correlation analysis suggested high ubiquinol and low ubiquinone in plasma promoted primate’s social motivation and ir detected lower body temperature. J. Clin. Toxicol. 2013, 3, 1000160. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.T.; Freiwald, W.A.; Leopold, D.A.; Mitchell, J.F.; Silva, A.C.; Wang, X. Marmosets: A neuroscientific model of human social behavior. Neuron 2016, 90, 219–233. [Google Scholar] [CrossRef]
- Abbott, D.H.; Barnett, D.K.; Colman, R.J.; Yamamoto, M.E.; Schultz-Darken, N.J. Aspects of common marmoset basic biology and life history important for biomedical research. Comp. Med. 2003, 53, 339–350. [Google Scholar]
- Tardif, S.D.; Smucny, D.A.; Abbott, D.H.; Mansfield, K.; Schultz-Darken, N.; Yamamoto, M.E. Reproduction in captive common marmosets (Callithrix jacchus). Comp. Med. 2003, 53, 364–368. [Google Scholar]
- Colman, R.J. Non-human primates as a model for aging. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2733–2741. [Google Scholar] [CrossRef] [PubMed]
- Schiel, N.; Souto, A. The common marmoset: An overview of its natural history, ecology and behavior. Dev. Neurobiol. 2017, 77, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.C.; Yen, C.C.; Ciuchta, J.L.; Papoti, D.; Bock, N.A.; Leopold, D.A.; Silva, A.C. Functional mapping of face-selective regions in the extrastriate visual cortex of the marmoset. J. Neurosci. 2015, 35, 1160–1172. [Google Scholar] [CrossRef] [PubMed]
- Voelkl, B.; Huber, L. Imitation as faithful copying of a novel technique in marmoset monkeys. PLoS ONE 2007, 2, e611. [Google Scholar] [CrossRef] [Green Version]
- Guyer, C.; Huber, R.; Fontijn, J.; Bucher, H.U.; Nicolai, H.; Werner, H.; Molinari, L.; Latal, B.; Jenni, O.G. Cycled light exposure reduces fussing and crying in very preterm infants. Pediatrics 2012, 130, e145–e151. [Google Scholar] [CrossRef] [Green Version]
- Wulff, K.; Gatti, S.; Wettstein, J.G.; Foster, R.G. Sleep and circadian rhythm disruption in psychiatric and neurodegenerative disease. Nat. Rev. Neurosci. 2010, 11, 589–599. [Google Scholar] [CrossRef]
- Mirmiran, M.; Bernardo, L.; Jenkins, S.L.; Ma, X.H.; Brenna, J.T.; Nathanielsz, P.W. Growth, neurobehavioral and circadian rhythm development in newborn baboons. Pediatr. Res. 2001, 49, 673–677. [Google Scholar] [CrossRef] [Green Version]
- Sumova, A.; Bendova, Z.; Sladek, M.; El-Hennamy, R.; Laurinova, K.; Jindrakova, Z.; Illnerova, H. Setting the biological time in central and peripheral clocks during ontogenesis. FEBS Lett. 2006, 580, 2836–2842. [Google Scholar] [CrossRef] [Green Version]
- Koshiba, M.; Senoo, A.; Mimura, K.; Shirakawa, Y.; Karino, G.; Obara, S.; Ozawa, S.; Sekihara, H.; Fukushima, Y.; Ueda, T.; et al. A cross-species socio-emotional behaviour development revealed by a multivariate analysis. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef]
- Koshiba, M.; Karino, G.; Senoo, A.; Mimura, K.; Shirakawa, Y.; Fukushima, Y.; Obara, S.; Sekihara, H.; Ozawa, S.; Ikegami, K.; et al. Peer attachment formation by systemic redox regulation with social training after a sensitive period. Sci. Rep. 2013, 3, 2503. [Google Scholar] [CrossRef]
- Hartley, C.A.; Lee, F.S. Sensitive periods in affective development: Nonlinear maturation of fear learning. Neuropsychopharmacology 2015, 40, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, T.B.; Russig, H.; Weiss, I.C.; Graff, J.; Linder, N.; Michalon, A.; Vizi, S.; Mansuy, I.M. Epigenetic transmission of the impact of early stress across generations. Biol. Psychiatry 2010, 68, 408–415. [Google Scholar] [CrossRef]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Levenson, J.M.; Sweatt, J.D. Epigenetic mechanisms in memory formation. Nat. Rev. Neurosci. 2005, 6, 108–118. [Google Scholar] [CrossRef]
- Bains, R.S.; Wells, S.; Sillito, R.R.; Armstrong, J.D.; Cater, H.L.; Banks, G.; Nolan, P.M. Assessing mouse behaviour throughout the light/dark cycle using automated in-cage analysis tools. J. Neurosci. Methods 2018, 300, 37–47. [Google Scholar] [CrossRef]
- Genewsky, A.; Heinz, D.E.; Kaplick, P.M.; Kilonzo, K.; Wotjak, C.T. A simplified microwave-based motion detector for home cage activity monitoring in mice. J. Biol. Eng. 2017, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.B.; Kunczik, J.; Zieglowski, L.; Tolba, R.; Abdelrahman, A.; Zechner, D.; Vollmar, B.; Janssen, H.; Thum, T.; Czaplik, M. Remote welfare monitoring of rodents using thermal imaging. Sensors 2018, 18, 3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, L.; Spruijt, B.; Riedel, G. Between and within laboratory reliability of mouse behaviour recorded in home-cage and open-field. J. Neurosci. Methods 2018, 300, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.; Coolen, A.; Schlumbohm, C.; Meerlo, P.; Fuchs, E. Remote long-term registrations of sleep-wake rhythms, core body temperature and activity in marmoset monkeys. Behav. Brain Res. 2012, 235, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Wolthuis, O.L.; Groen, B.; Philippens, I.H. A simple automated test to measure exploratory and motor activity of marmosets. Pharmacol. Biochem. Behav. 1994, 47, 879–881. [Google Scholar] [CrossRef]
- Senoo, A.; Okuya, T.; Sugiura, Y.; Mimura, K.; Honda, Y.; Tanaka, I.; Kodama, T.; Tokuno, H.; Yui, K.; Nakamura, S.; et al. Effects of constant daylight exposure during early development on marmoset psychosocial behavior. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Koshiba, M.; Mimura, K.; Sugiura, Y.; Okuya, T.; Senoo, A.; Ishibashi, H.; Nakamura, S. Reading marmoset behavior ‘semantics’ under particular social context by multi-parameters correlation analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1499–1504. [Google Scholar] [CrossRef]
- Braesicke, K.; Parkinson, J.A.; Reekie, Y.; Man, M.S.; Hopewell, L.; Pears, A.; Crofts, H.; Schnell, C.R.; Roberts, A.C. Autonomic arousal in an appetitive context in primates: A behavioural and neural analysis. Eur. J. Neurosci. 2005, 21, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Arenas, M.C.; Aguilar, M.A.; Montagud-Romero, S.; Mateos-García, A.; Navarro-Francés, C.I.; Miñarro, J.; Rodríguez-Arias, M. Influence of the Novelty-Seeking Endophenotype on the Rewarding Effects of Psychostimulant Drugs in Animal Models. Curr Neuropharmacol. 2016, 14, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentinuzzi, V.S.; Neto, S.P.; Carneiro, B.T.; Santana, K.S.; Araujo, J.F.; Ralph, M.R. Memory for time of training modulates performance on a place conditioning task in marmosets. Neurobiol. Learn. Mem. 2008, 89, 604–607. [Google Scholar] [CrossRef]
- Monclaro, A.V.; Sampaio, A.C.; Ribeiro, N.B.; Barros, M. Time-of-day effect on a food-induced conditioned place preference task in monkeys. Behav. Brain Res. 2014, 259, 336–341. [Google Scholar] [CrossRef]
- Fritz, M.; El Rawas, R.; Klement, S.; Kummer, K.; Mayr, M.J.; Eggart, V.; Salti, A.; Bardo, M.T.; Saria, A.; Zernig, G. Differential effects of accumbens core vs. shell lesions in a rat concurrent conditioned place preference paradigm for cocaine vs. social interaction. PLoS ONE 2011, 6, e26761. [Google Scholar] [CrossRef] [Green Version]
- El Rawas, R.; Klement, S.; Kummer, K.K.; Fritz, M.; Dechant, G.; Saria, A.; Zernig, G. Brain regions associated with the acquisition of conditioned place preference for cocaine vs. social interaction. Front. Behav. Neurosci. 2012, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Koshiba, M.; Shirakawa, Y.; Mimura, K.; Senoo, A.; Karino, G.; Nakamura, S. Familiarity perception call elicited under restricted sensory cues in peer-social interactions of the domestic chick. PLoS ONE 2013, 8, e58847. [Google Scholar] [CrossRef] [Green Version]
- Mimura, K.; Nakamura, S.; Koshiba, M. A flexion period for attachment formation in isolated chicks to unfamiliar peers visualized in a developmental trajectory space through behavioral multivariate correlation analysis. Neurosci. Lett. 2013, 547, 70–75. [Google Scholar] [CrossRef]
- Mimura, K.; Mochizuki, D.; Nakamura, S.; Koshiba, M. A sensitive period of peer-social learning. J. Clin. Toxicol. 2013, 3, 000158. [Google Scholar] [CrossRef]
- Koshiba, M.; Kakei, H.; Honda, M.; Karino, G.; Niitsu, M.; Miyaji, T.; Kishino, H.; Nakamura, S.; Kunikata, T.; Yamanouchi, H. Early-infant diagnostic predictors of the neuro-behavioral development after neonatal care. Behav. Brain Res. 2015, 276, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Mimura, K.; Kishino, H.; Karino, G.; Nitta, E.; Senoo, A.; Ikegami, K.; Kunikata, T.; Yamanouchi, H.; Nakamura, S.; Sato, K.; et al. Potential of a smartphone as a stress-free sensor of daily human behaviour. Behav. Brain Res. 2015, 276, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshiba, M.; Senoo, A.; Karino, G.; Ozawa, S.; Tanaka, I.; Honda, Y.; Usui, S.; Kodama, T.; Mimura, K.; Nakamura, S.; et al. Susceptible period of socio-emotional development affected by constant exposure to daylight. Neurosci. Res. 2015, 93, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fifer, W.P.; Byrd, D.L.; Kaku, M.; Eigsti, I.M.; Isler, J.R.; Grose-Fifer, J.; Tarullo, A.R.; Balsam, P.D. Newborn infants learn during sleep. Proc. Natl. Acad. Sci. USA 2010, 107, 10320–10323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Nelder, J.A.; Pawitan, Y. Generalized Linear Models with Random Effects Unified Analysis via H-likelihood; Chapman & Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]
- Benjamin Saefken, T.K. A unifying approach to the estimation of the conditional Akaike information in generalized linear mixed models. Electron. J. Stat. 2014, 8, 201–225. [Google Scholar] [CrossRef]
- Rivkees, S.A. Developing circadian rhythmicity in infants. Pediatr. Endocrinol. Rev. 2003, 1, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Weinert, D. Ontogenetic development of the mammalian circadian system. Chronobiol. Int. 2005, 22, 179–205. [Google Scholar] [CrossRef]
- Weinert, D.; Waterhouse, J. The circadian rhythm of core temperature: Effects of physical activity and aging. Physiol. Behav. 2007, 90, 246–256. [Google Scholar] [CrossRef]
- Refinetti, R. Effects of food temperature and ambient temperature during a meal on food intake in the rat. Physiol. Behav. 1988, 43, 245–247. [Google Scholar] [CrossRef]
- Storch, K.F.; Weitz, C.J. Daily rhythms of food-anticipatory behavioral activity do not require the known circadian clock. Proc. Natl. Acad. Sci. USA 2009, 106, 6808–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.N.; Patton, D.F.; Michalik, M.; Opiol, H.; Mistlberger, R.E. Dopaminergic regulation of circadian food anticipatory activity rhythms in the rat. PLoS ONE 2013, 8, e82381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Ann. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, N.; Hioki, H.; Kaneko, T.; Nakamura, K. Psychological stress activates a dorsomedial hypothalamus-medullary raphe circuit driving brown adipose tissue thermogenesis and hyperthermia. Cell Metab. 2014, 20, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Tao, T.; Sakurai, H.; Kakei, H.; Morita, K.; Honda, M.; Kamei, Y.; Yamanouchi, H.; Kunikata, T.; Koshiba, M. Preterm infant vocal behavior and SpO2, pulse rate modulation in neonatal intensive care unit. Stress Brain Behav. 2019, 1, e019006. [Google Scholar] [CrossRef]
- Tao, T.; Sakurai, H.; Kakei, H.; Morita, K.; Honda, M.; Jiang, Z.; Yamanouchi, H.; Kunikata, T.; Koshiba, M. Longitudinal recording revealed preterm infants’ acoustic experiences per individual in clinical environments. Stress Brain Behav. 2019, 9, 5–8. [Google Scholar]
- Carneiro, B.T.S.; Fernandes, D.A.C.; Medeiros, C.F.P.; Diniz, N.L.; Araujo, J.F. Daily anticipatory rhythms of behavior and body temperature in response to glucose availability in rats. Psychol. Neurosci. 2012, 5, 191–197. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | All | St1 | St2 | St3 | St4 | |
|---|---|---|---|---|---|---|
| 1 | The ratio of non-parsable images in the images in which there was no marmoset | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| 2 | The ratio of parsable images in the images in which there was a marmoset | 0.858 | 0.919 | 0.943 | 0.740 | 0.831 |
| 3 | The ratio of correctly parsable images in all the parsable images | 0.998 | 0.998 | 0.996 | 1.000 | 0.999 |
| All | St1 | St2 | St3 | St4 | ||
|---|---|---|---|---|---|---|
| alpha | X | 0.998 | 0.993 | 0.995 | 1.004 | 1.003 |
| Y | 1.020 | 1.027 | 1.033 | 1.010 | 1.011 | |
| BV | 1.038 | 1.052 | 1.073 | 0.966 | 0.965 | |
| BT | 0.999 | 0.999 | 0.999 | 1.001 | 0.999 | |
| R2 | X | 0.992 | 0.992 | 0.989 | 0.997 | 0.996 |
| Y | 0.987 | 0.990 | 0.976 | 0.991 | 0.995 | |
| BV | 0.964 | 0.965 | 0.961 | 0.968 | 0.968 | |
| BT | 0.935 | 0.919 | 0.911 | 0.962 | 0.966 |
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | |
|---|---|---|---|---|---|---|---|
| Contribution Rate | 0.213 | 0.174 | 0.163 | 0.143 | 0.132 | 0.104 | 0.072 |
| Cumulative Contribution Ratio | 0.213 | 0.386 | 0.550 | 0.692 | 0.824 | 0.928 | 1.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karino, G.; Senoo, A.; Kunikata, T.; Kamei, Y.; Yamanouchi, H.; Nakamura, S.; Shukuya, M.; Colman, R.J.; Koshiba, M. Inexpensive Home Infrared Living/Environment Sensor with Regional Thermal Information for Infant Physical and Psychological Development. Int. J. Environ. Res. Public Health 2020, 17, 6844. https://doi.org/10.3390/ijerph17186844
Karino G, Senoo A, Kunikata T, Kamei Y, Yamanouchi H, Nakamura S, Shukuya M, Colman RJ, Koshiba M. Inexpensive Home Infrared Living/Environment Sensor with Regional Thermal Information for Infant Physical and Psychological Development. International Journal of Environmental Research and Public Health. 2020; 17(18):6844. https://doi.org/10.3390/ijerph17186844
Chicago/Turabian StyleKarino, Genta, Aya Senoo, Tetsuya Kunikata, Yoshimasa Kamei, Hideo Yamanouchi, Shun Nakamura, Masanori Shukuya, Ricki J. Colman, and Mamiko Koshiba. 2020. "Inexpensive Home Infrared Living/Environment Sensor with Regional Thermal Information for Infant Physical and Psychological Development" International Journal of Environmental Research and Public Health 17, no. 18: 6844. https://doi.org/10.3390/ijerph17186844
APA StyleKarino, G., Senoo, A., Kunikata, T., Kamei, Y., Yamanouchi, H., Nakamura, S., Shukuya, M., Colman, R. J., & Koshiba, M. (2020). Inexpensive Home Infrared Living/Environment Sensor with Regional Thermal Information for Infant Physical and Psychological Development. International Journal of Environmental Research and Public Health, 17(18), 6844. https://doi.org/10.3390/ijerph17186844