Maximal Time Spent at VO2max from Sprint to the Marathon



Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of Maximal Oxygen Uptake and Velocity Associated with VO2max—The UMTT
2.2. Determination of The Time Limit at VO2max (Tlim at VO2max)
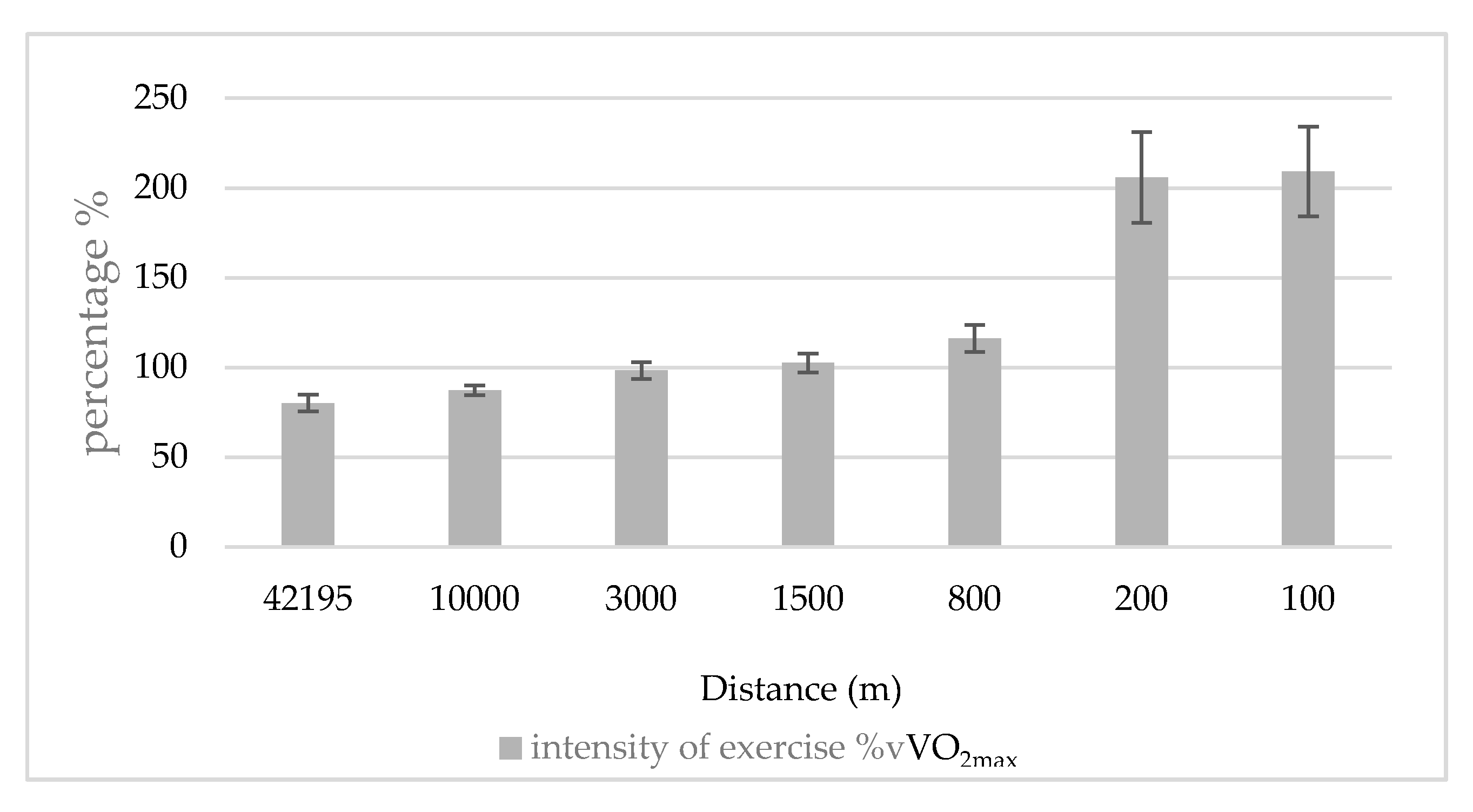
2.3. Calculation of the Intensity of Race in the Percentage of Vvo2max (Intensity of Exercise %Vvo2max)
2.4. Statistical Analysis
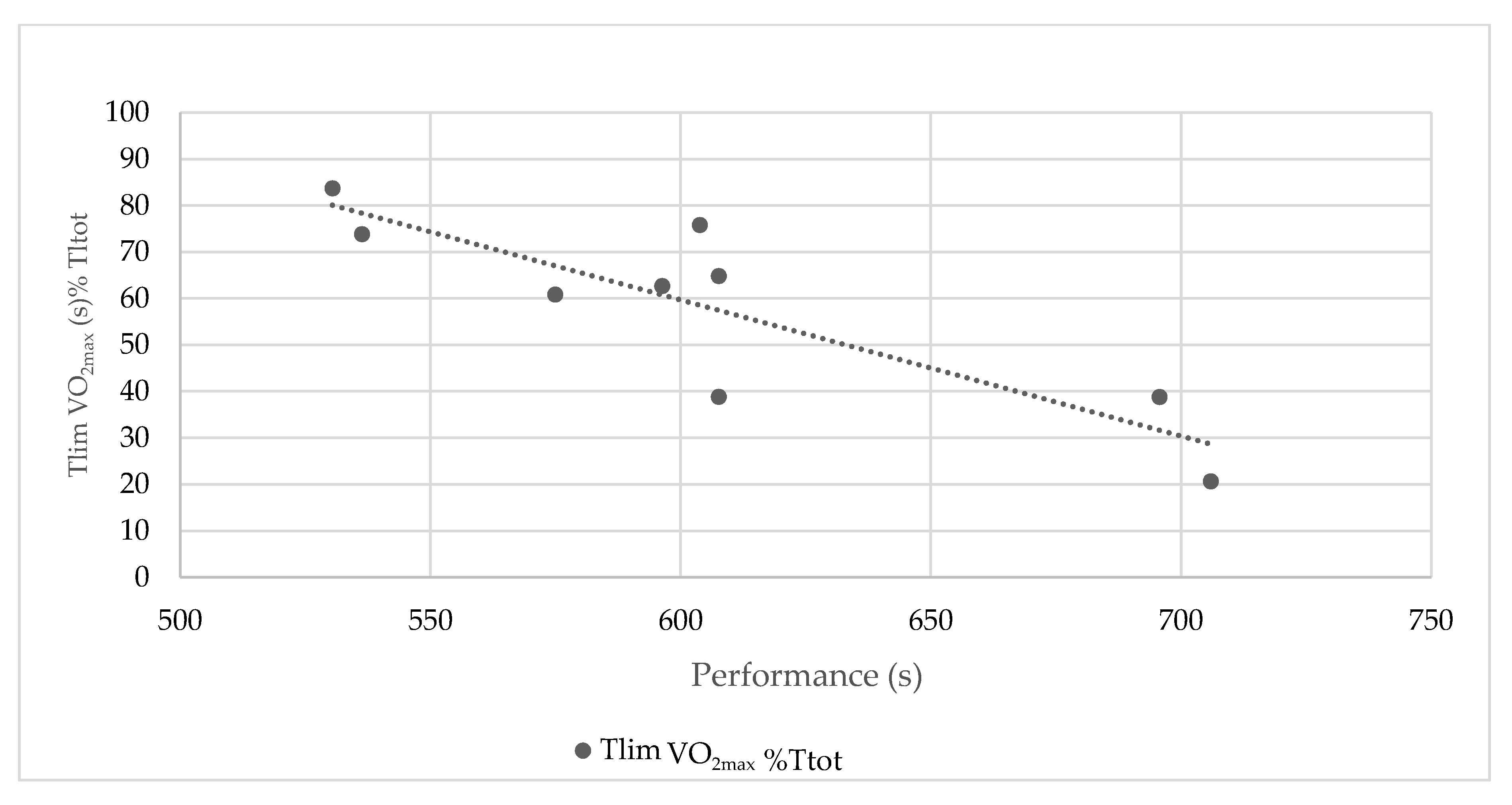
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Billat, V.; Hamard, L.; Koralsztein, J.P.; Morton, R.H. Differential modeling of anaerobic and aerobic metabolism in the 800-m and 1500-m run. J. Appl. Physiol. 2009, 107, 478–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, R.H.; Billat, V. Maximal endurance time at VO2max. Med. Sci. Sports Exerc. 2000, 32, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V.; Lupton, H. Muscular Exercise, Lactic Acid, and the Supply and Utilization of Oxygen. QJM Int. J. Med. 1923, 62, 135–171. [Google Scholar] [CrossRef]
- Saks, V.A.; Kongas, O.; Vendelin, M.; Kay, L. Role of the creatine/phosphocreatine system in the regulation of mitochondrial respiration. Acta Physiol. Scand. 2000, 168, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B.; Zoladz, J.A. Possible mechanisms underlying slow component of Vo2 on-kinetics in skeletal muscle. J. Appl. Physiol. 2015, 118, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Di Prampero, P.E. The energy cost of human locomotion on land and in water. Int. J. Sports Med. 1986, 7, 55–72. [Google Scholar] [CrossRef]
- Zinner, C. Training Aspects of Marathon Running. In Marathon Running: Physiology, Psychology, Nutrition and Training Aspects; Zinner, C., Sperlich, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 153–171. [Google Scholar] [CrossRef]
- Billat, V.; Bernard, O.; Pinoteau, J.; Petit, B.; Koralsztein, J.P. Time to exhaustion at VO2max and lactate steady state velocity in sub elite long-distance runners. Arch. Int. Physiol. Biochim. Biophys. 1994, 102, 215–219. [Google Scholar] [CrossRef]
- Whipp, B.J. The slow component of O2 uptake kinetics during heavy exercise. Med. Sci. Sports Exerc. 1994, 26, 1319–1326. [Google Scholar] [CrossRef]
- Maron, M.B.; Horvath, S.M.; Wilkerson, J.E.; Gliner, J.A. Oxygen uptake measurements during competitive marathon running. J. Appl. Physiol. 1976, 40, 836–838. [Google Scholar] [CrossRef]
- Billat, V.; Dalmay, F.; Antonini, M.T.; Chassain, A.P. A method for determining the maximal steady state of blood lactate concentration from two levels of submaximal exercise. Eur. J. Appl. Physiol. 1994, 69, 196–202. [Google Scholar] [CrossRef]
- Billat, V.L.; Morton, R.H.; Blondel, N.; Berthoin, S.; Bocquet, V.; Koralsztein, J.P.; Barstow, T.J. Oxygen kinetics and modelling of time to exhaustion whilst running at various velocities at maximal oxygen uptake. Eur. J. Appl. Physiol. 2000, 82, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.; Petot, H.; Karp, J.R.; Sarre, G.; Morton, R.H.; Mille-Hamard, L. The sustainability of VO2max: Effect of decreasing the workload. Eur. J. Appl. Physiol. 2013, 113, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Bergh, U.; Ekblom, B.; Astrand, P.O. Maximal oxygen uptake ‘classical’ versus ‘contemporary’ viewpoints. Med. Sci. Sports Exerc. 2000, 32, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Ekblom, B. Counterpoint: Maximal oxygen uptake is not limited by a central nervous system governor. J. Appl. Physiol. 2009, 106, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.; Vitiello, D.; Palacin, F.; Correa, M.; Pycke, J.R. Race Analysis of the World’s Best Female and Male Marathon Runners. Int. J. Environ. Res. Public Health 2020, 17, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Prampero, P.E.; Botter, A.; Osgnach, C. The energy cost of sprint running and the role of metabolic power in setting top performances. Eur. J. Appl. Physiol. 2015, 115, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Perez-Suarez, I.; Martin-Rincon, M.; Gonzalez-Henriquez, J.J.; Fezzardi, C.; Perez-Regalado, S.; Galvan-Alvarez, V.; Juan-Habib, J.W.; Morales-Alamo, D.; Calbet, J.A. Accuracy and Precision of the COSMED K5 Portable Analyser. Front. Physiol 2018, 9, 1764. [Google Scholar] [CrossRef]
- Borg, G. Borg’s Perceived Exertion and Pain Scales; Human Kinetics: Champaign, IL, USA, 1998. [Google Scholar]
- Hogg, J.S.; Hopker, J.G.; Mauger, A.R. The Self-Paced VO2max Test to Assess Maximal Oxygen Uptake in Highly Trained Runners. Int. J. Sports Physiol. Perform. 2015, 10, 172–177. [Google Scholar] [CrossRef]
- Gaesser, G.A.; Poole, D.C. The Slow Component of Oxygen Uptake Kinetics in Humans. Exerc. Sport Sci. Rev. 1996, 24, 35. [Google Scholar] [CrossRef]
- Morton, R.H. A three component model of human bioenergetics. J. Math. Biol. 1986, 24, 451–466. [Google Scholar] [CrossRef]
- Barstow, T.J.; Mole, P.A. Linear and nonlinear characteristics of oxygen uptake kinetics during heavy exercise. J. Appl. Physiol. 1991, 71, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Glaister, M. Multiple Sprint Work. Sports Med. 2005, 35, 757–777. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, H.B.; Ward, S.A.; Kowalchuk, J.M.; Howe, F.A.; Griffiths, J.R.; Whipp, B.J. Dynamic asymmetry of phosphocreatine concentration and O(2) uptake between the on- and off-transients of moderate- and high-intensity exercise in humans. J. Physiol. 2002, 541, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Volkov, N.I.; Shirkovets, E.A.; Borilkevich, V.E. Assessment of aerobic and anaerobic capacity of athletes in treadmill running tests. Eur. J. Appl. Physiol. 1975, 34, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Bogdanis, G.C.; Nevill, M.E.; Boobis, L.H.; Lakomy, H.K. Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J. Appl. Physiol. 1996, 80, 876–884. [Google Scholar] [CrossRef] [PubMed]
- McGawley, K.; Bishop, D.J. Oxygen uptake during repeated-sprint exercise. J. Sci. Med. Sport 2015, 18, 214–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchheit, M.; Ufland, P. Effect of endurance training on performance and muscle reoxygenation rate during repeated-sprint running. Eur. J. Appl. Physiol. 2011, 111, 293–301. [Google Scholar] [CrossRef]
- Belfry, G.R.; Paterson, D.H.; Murias, J.M.; Thomas, S.G. The effects of short recovery duration on VO2 and muscle deoxygenation during intermittent exercise. Eur. J. Appl. Physiol. 2012, 112, 1907–1915. [Google Scholar] [CrossRef]
- Weyand, P.; Curcton, K.; Conley, D.; Sloniger, M. Percentage Anaerobic Energy Utilized During Track Running Events. Med. Sci. Sports Exerc. 1993, 25, S105. [Google Scholar] [CrossRef]
- Billat, V.L.; Demarle, A.; Slawinski, J.; Paiva, M.; Koralsztein, J.P. Physical and training characteristics of top-class marathon runners. Med. Sci. Sports Exerc. 2001, 33, 2089–2097. [Google Scholar] [CrossRef]
- Nikolaidis, P.T.; Knechtle, B. Do Fast Older Runners Pace Differently From Fast Younger Runners in the “New York City Marathon”? J. Strength Cond. Res. 2019, 33, 3423–3430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundle, M.W.; Hoyt, R.W.; Weyand, P.G. High-speed running performance: A new approach to assessment and prediction. J. Appl. Physiol. 2003, 95, 1955–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, M.; Bishop, D.; Dawson, B.; Goodman, C. Physiological and Metabolic Responses of Repeated-Sprint Activities. Sports Med. 2005, 35, 1025–1044. [Google Scholar] [CrossRef] [PubMed]
- Hirvonen, J.; Rehunen, S.; Rusko, H.; Härkönen, M. Breakdown of high-energy phosphate compounds and lactate accumulation during short supramaximal exercise. Eur. J. Appl. Physiol. 1987, 56, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B.; Zoladz, J.A. Factors determining the oxygen consumption rate (V.o2) on-kinetics in skeletal muscles. Biochem. J. 2004, 379, 703–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heubert, R.A.P.; Billat, V.L.; Chassaing, P.; Bocquet, V.; Morton, R.H.; Koralsztein, J.P.; Di Prampero, P.E. Effect of a previous sprint on the parameters of the work-time to exhaustion relationship in high intensity cycling. Int. J. Sports Med. 2005, 26, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Wilkerson, D.P.; Vanhatalo, A.; Burnley, M. Influence of pacing strategy on O2 uptake and exercise tolerance. Scand. J. Med. Sci. Sports 2008, 18, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Sandals, L.E.; Wood, D.M.; Draper, S.B.; James, D.V. Influence of Pacing Strategy on Oxygen Uptake During Treadmill Middle-Distance Running. Int. J. Sports Med. 2006, 27, 37–42. [Google Scholar] [CrossRef]
- Sahlin, K. High-Energy Phosphates and Muscle Energetics. Princ. Exerc. Biochem. 2004, 46, 87–107. [Google Scholar] [CrossRef]
- Hanon, C.; Thomas, C. Effects of optimal pacing strategies for 400-, 800-, and 1500-m races on the [Vdot]O2 response. J. Sports Sci. 2011, 29, 905–912. [Google Scholar] [CrossRef]
- Billat, V.; Carbillet, T.; Correa, M.; Pycke, J.R. Detecting the marathon asymmetry with a statistical signature. Phys. Stat. Mech. Appl. 2019, 515, 240–247. [Google Scholar] [CrossRef]
- Blondel, N.; Berthoin, S.; Billat, V.; Lensel, G. Relationship between Run Times to Exhaustion at 90, 100, 120, and 140 % of vV˙O2max and Velocity Expressed Relatively to Critical Velocity and Maximal Velocity. Int. J. Sports Med. 2001, 22, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.L.; Blondel, N.; Berthoin, S. Determination of the velocity associated with the longest time to exhaustion at maximal oxygen uptake. Eur. J. Appl. Physiol. 1999, 80, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Burnley, M.; Vanhatalo, A.; Rossiter, H.B.; Jones, A.M. Critical Power: An Important Fatigue Threshold in Exercise Physiology. Med. Sci. Sports Exerc. 2016, 48, 2320–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Vanhatalo, A.; Burnley, M.; Morton, R.H.; Poole, D.C. Critical power: Implications for determination of V˙O2max and exercise tolerance. Med. Sci. Sports Exerc. 2010, 42, 1876–1890. [Google Scholar] [CrossRef]
- Blondel, N.; Billat, V.; Berthoin, S. Relation entre le temps limite de course et l’intensité relative de l’exercice, exprimée en fonction de la vitesse critique et de la vitesse maximale. Sci. Sports 2000, 15, 242–244. [Google Scholar] [CrossRef]
- Faina, M.; Billat, V.; Squadrone, R.; De Angelis, M.; Koralsztein, J.P.; Dal Monte, A. Anaerobic contribution to the time to exhaustion at the minimal exercise intensity at which maximal oxygen uptake occurs in elite cyclists, kayakists and swimmers. Eur. J. Appl. Physiol. 1997, 76, 13–20. [Google Scholar] [CrossRef]
- Billat, V.L.; Slawinski, J.; Bocquet, V.; Demarle, A.; Lafitte, L.; Chassaing, P.; Koralsztein, J.P. Intermittent runs at the velocity associated with maximal oxygen uptake enables subjects to remain at maximal oxygen uptake for a longer time than intense but submaximal runs. Eur. J. Appl. Physiol. 2000, 81, 188–196. [Google Scholar] [CrossRef]
- Billat, V.L.; Slawinksi, J.; Bocquet, V.; Chassaing, P.; Demarle, A.; Koralsztein, J.P. Very short (15 s–15 s) interval-training around the critical velocity allows middle-aged runners to maintain VO2 max for 14 minutes. Int. J. Sports Med. 2001, 22, 201–208. [Google Scholar] [CrossRef]
- Petot, H.; Meilland, R.; Le Moyec, L.; Mille-Hamard, L.; Billat, V.L. A new incremental test for VO2max accurate measurement by increasing VO2max plateau duration, allowing the investigation of its limiting factors. Eur. J. Appl. Physiol. 2012, 112, 2267–2276. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Runners | n | vVO2max (km∙h−1) | VO2max (mL∙kg−1∙min−1) | HRmax (Beat·min−1) | RPE Last Stage of UMTT |
|---|---|---|---|---|---|
| 100 m | 13 | 15.4 ± 1.6 | 53.1 ± 5.5 | 196.3 ± 4.5 | 19.5 ± 0.5 |
| 200 m | 13 | 15.4 ± 1.6 | 53.1 ± 5.5 | 196.3 ± 4.5 | 19.5 ± 0.5 |
| 800 m | 8 | 19.3 ± 0.7 ab | 64.6 ± 3.4 ab | 196.9 ± 6.4 | 19.7 ± 0.5 |
| 1500 m | 16 | 17.8 ± 2.2 ab | 59.0 ± 10.5 | 188.6 ± 12.6 | 19.8 ± 0.4 |
| 3000 m | 9 | 16.2 ± 1.0 abc | 51.1 ± 5.3 cd | 181.9 ± 11.7 abc | 19.9 ± 0.3 |
| 10,000 m | 7 | 19.1 ± 1.8 abe | 67.0 ± 6.5 abef | 183.4 ± 11.2 abc | 19.3 ± 0.5 de |
| 42,195 m | 12 | 17.0± 0.9 abc | 55.4 ± 4.7 c | 189.1 ± 8.2 abc | 19.5 ± 0.5 |
| Distance | n | IAAF Score | Race Time (hh:min:sec) | VO2max Reached (n, %) | Tlim at VO2max (s) | Tlim at VO2max%Ttot | Post-Run Lactate (mmol·L−1) |
|---|---|---|---|---|---|---|---|
| 100 m | 13 | 799.0 ± 143.5 | 11″ ± 0.5″ | 10 (76%) | 3 ± 2.1 | 25.6 ± 18.5 | 14.0 ± 2.8 |
| 200 m | 13 | 795.5 ± 135.5 | 23″ ± 1″1 | 11 (85%) | 6 ± 4.0 | 28.5 ± 17.7 | 14.9 ± 1.5 |
| 800 m | 8 | 563.0 ± 131.0 ab | 2′09″ ± 6″4 f | 8 (100%) | 28 ± 19.7 aef | 22.0 ± 15.8 | 15.9 ± 1.7 |
| 1500 m | 16 | 474.6 ± 191.8 ab | 4′40″ ± 24″7 acd | 15 (94%) | 129 ± 92.2 abe | 41.7 ± 28.6 | 12.4 ± 1.8 bc |
| 3000 m | 9 | 472.2 ± 218.8 ab | 10′07″ ± 1′9″ ab | 8 (89%) | 341 ± 103.3 abcd | 51.4 ± 18.3 abc | 11.7 ± 2.3 bc |
| 10,000 m | 7 | 522.4 ± 242.5 ab | 36′22″ ± 4′19″ ab | 7 (100%) | 680 ± 590.6 abcd | 30.6 ± 27.2 f | / |
| 42,195 m | 12 | 385.6 ± 190.7 ab | 3h7′17″ ± 18′41″ abcd | 10 (83%) | 479 ± 497.9 abc | 4.1 ± 4.0 abcde | 6.6 ± 2.1 abcde |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinari, C.A.; Edwards, J.; Billat, V. Maximal Time Spent at VO2max from Sprint to the Marathon. Int. J. Environ. Res. Public Health 2020, 17, 9250. https://doi.org/10.3390/ijerph17249250
Molinari CA, Edwards J, Billat V. Maximal Time Spent at VO2max from Sprint to the Marathon. International Journal of Environmental Research and Public Health. 2020; 17(24):9250. https://doi.org/10.3390/ijerph17249250
Chicago/Turabian StyleMolinari, Claire A., Johnathan Edwards, and Véronique Billat. 2020. "Maximal Time Spent at VO2max from Sprint to the Marathon" International Journal of Environmental Research and Public Health 17, no. 24: 9250. https://doi.org/10.3390/ijerph17249250
APA StyleMolinari, C. A., Edwards, J., & Billat, V. (2020). Maximal Time Spent at VO2max from Sprint to the Marathon. International Journal of Environmental Research and Public Health, 17(24), 9250. https://doi.org/10.3390/ijerph17249250