Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway

Abstract
:1. Introduction
2. Methods
2.1. Subjects and Groups
2.2. Modeling Depression in Mice
2.3. Behavioral Assessment
2.4. Aerobic Exercise Plan
2.5. Sample Preparation
2.6. Diagnostic Assay
2.6.1. Western Blot
2.6.2. Realtime-PCR
2.6.3. Elisa
2.7. Data Processing
3. Results
3.1. Weight Results
3.2. Behavioral Results
3.3. Western Blot Results
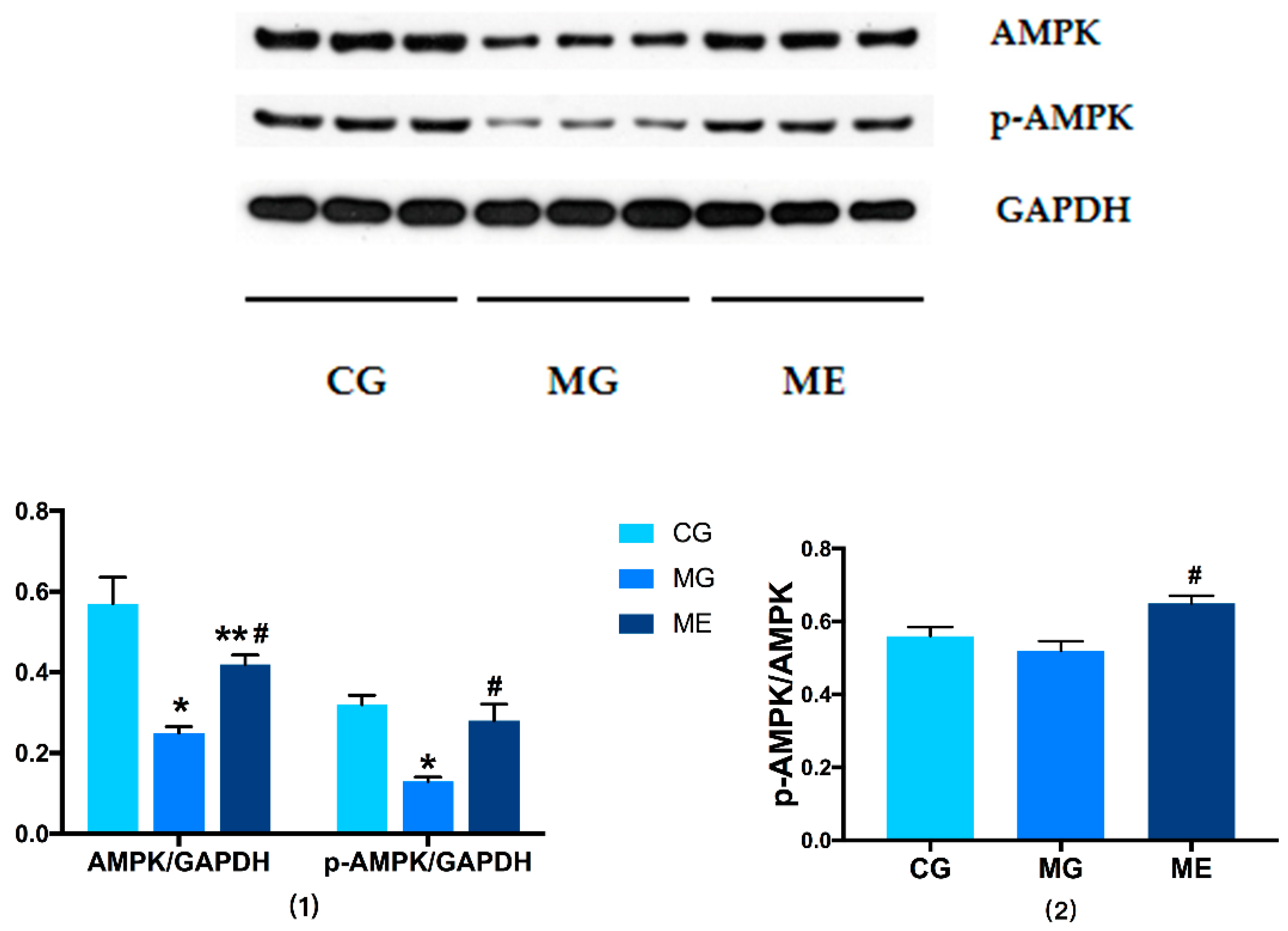
3.3.1. Changes in AMPK/p-AMPK Protein Expression
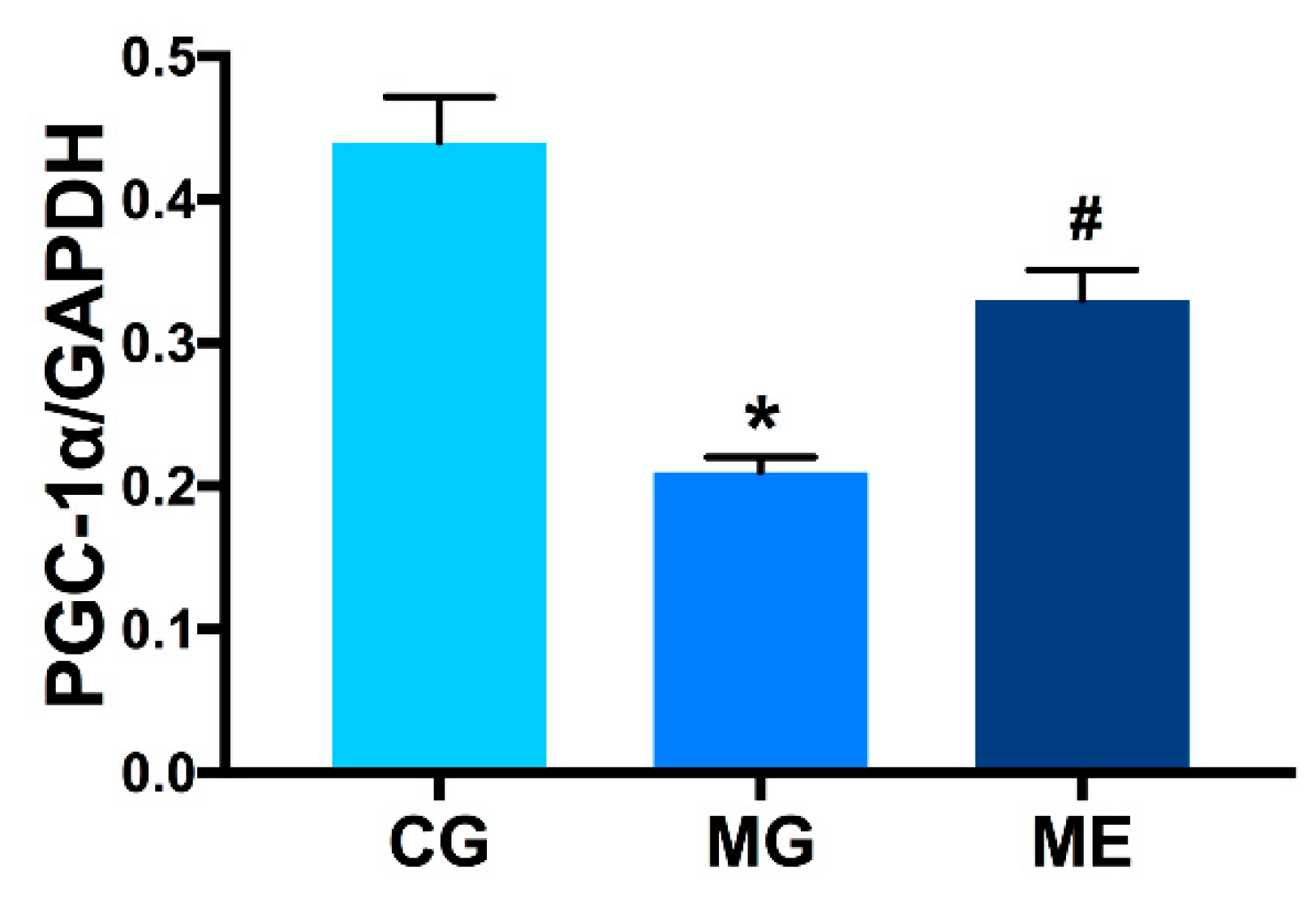
3.3.2. Changes in PGC-1α Protein Expression
3.4. RT-PCR on AMPK and PGC-1αmRNA Expression
AMPK and PGC-1αmRNA Expression
3.5. Elisa Results
4. Discussion
4.1. Discussion on Chronic Stress-Induced Depression Model in Mice
4.2. Impacts of Aerobic Exercise on the Behavior and Weight of Mice with CUMS-Induced Depression-Like Behaviors
4.3. Impacts of Aerobic Exercise on AMPK/PGC-1α of Mice with CUMS-Induced Depression-Like Behaviors
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hou, Z.; Qin, X.; Hu, Y.; Zhang, X.; Li, G.; Wu, J.; Li, J.; Sha, J.; Chen, J.; Xia, J.; et al. Longterm Exercise-Derived Exosomal miR-342-5p: A Novel Exerkine for Cardioprotection. Circ. Res. 2019, 124, 1386–1400. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, Z. Swimming training affects apoptosis-related microRNAs and reduces cardiac apoptosis in mice. Gen. Physiol. Biophys. 2016, 35, 443–450. [Google Scholar] [CrossRef]
- Shakil-Ur-Rehman, S.; Karimi, H.; Gillani, S.A. Effects of supervised structured aerobic exercise training program on fasting blood glucose level, plasma insulin level, glycemic control, and insulin resistance in type 2 diabetes mellitus. Pak. J. Med. Sci. 2017, 33, 576–580. [Google Scholar] [CrossRef]
- Du, J.; Wang, Y.; Zhi, N.; Geng, J.; Cao, W.; Yu, L.; Mi, J.; Zhou, Y.; Xu, Q.; Wen, W.; et al. Structural brain network measures are superior to vascular burden scores in predicting early cognitive impairment in post stroke patients with small vessel disease. Neuroimage Clin. 2019, 22, 101712. [Google Scholar] [CrossRef]
- Smith, K. Mental health: A world of depression. Nat. J. 2014, 515, 181. [Google Scholar] [CrossRef]
- Yoon, H.S.; Hattori, K.; Ogawa, S.; Sasayama, D.; Ota, M.; Teraishi, T.; Kunugi, H. Relationships of Cerebrospinal Fluid Monoamine Metabolite Levels with Clinical Variables in Major Depressive Disorder. J. Clin. Psychiatry 2017, 78, e947–e956. [Google Scholar] [CrossRef]
- Pukhalsky, A.L.; Shmarina, G.V.; Alioshkin, V.A.; Sabelnikov, A. HPA axis exhaustion and regulatory T cell accumulation in patients with a functional somatic syndrome: Recent view on the problem of Gulf War veterans. J. Neuroimmunol. 2008, 196, 133–138. [Google Scholar] [CrossRef]
- Morgan, J.A.; Singhal, G.; Corrigan, F.; Jaehne, E.J.; Jawahar, M.C.; Baune, B.T. Exercise related anxiety-like behaviours are mediated by TNF receptor signaling, but not depression-like behaviours. Brain Res. 2018, 1695, 10–17. [Google Scholar] [CrossRef]
- Ye, F.; Zhan, Q.; Xiao, W.; Tang, X.; Li, J.; Dong, H.; Sha, W.; Zhang, X. Altered serum levels of vascular endothelial growth factor in first-episode drug-naive and chronic medicated schizophrenia. Psychiatry Res. 2018, 264, 361–365. [Google Scholar] [CrossRef]
- Jemili, H.; Mejri, M.A.; Bouhlel, E.; Amri, M. Biochemical status, oxidative and antioxidant responses after 3-month specific training in elite karate athletes. Physiol. Int. 2017, 104, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Karabatsiakis, A.; Bock, C.; Salinas-Manrique, J.; Kolassa, S.; Calzia, E.; Dietrich, D.E.; Kolassa, I.T. Mitochondrial respiration in peripheral blood mononuclear cells correlates with depressive subsymptoms and severity of major depression. Transl. Psychiatry. 2014, 4, e397. [Google Scholar] [CrossRef] [Green Version]
- Fattal, O.; Budur, K.; Vaughan, A.J.; Franco, K. Review of the literature on major mental disorders in adult patients with mitochondrial diseases. Psychosomatics 2006, 47, 1–7. [Google Scholar] [CrossRef]
- Carling, D. AMPK signalling in health and disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.; Sun, R.; Tao, X.; Wang, X.; Kang, Q.; Wang, H.; Zhang, L.; Liu, P.; Zhang, J.; et al. SIRT5 deficiency suppresses mitochondrial ATP production and promotes AMPK activation in response to energy stress. PLoS ONE 2019, 14, e0211796. [Google Scholar] [CrossRef]
- Canto, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Auwerx, P.P.J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Vella, C.A.; Nelson, O.L.; Jansen, H.T.; Robbins, C.T.; Jensen, A.E.; Constantinescu, S.; Abbott, M.J.; Turcotte, L.P. Regulation of metabolism during hibernation in brown bears (Ursus arctos): Involvement of cortisol, PGC-1alpha and AMPK in adipose tissue and skeletal muscle. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 240, 110591. [Google Scholar] [CrossRef]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharm. Acology 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Caichuan, W. Study of chronic stress stimulation on behavior of depression model mice. J. Med. Sci. Yan Bian Univ. 2017, 40, 92–96. [Google Scholar]
- Bedford, T.G.; Tipton, C.M.; Wilson, N.C.; Oppliger, R.A.; Gisolfi, C.V. Maximum oxygen consumption of rats and its changes with various experimental procedures. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1979, 47, 1278–1283. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nat. J. 2008, 7215, 894–902. [Google Scholar] [CrossRef]
- Gu, Z.; Pan, J.; Chen, L. MiR-124 suppression in the prefrontal cortex reduces depression-like behavior in mice. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, B.; Pochwat, B.; Muszynska, B.; Opoka, W.; Krakowska, A.; Rafalo-Ulinska, A.; Friedland, K.; Nowak, G. Antidepressant-like activity of hyperforin and changes in BDNF and zinc levels in mice exposed to chronic unpredictable mild stress. Behav. Brain Res. 2019, 372, 112045. [Google Scholar] [CrossRef]
- Shehu, A.; Magaji, M.G.; Yau, J.; Ahmed, A. Methanol stem bark extract of Adansonia digitata ameliorates chronic unpredictable mild stress-induced depression-like behavior: Involvement of the HPA axis, BDNF, and stress biomarkers pathways. J. Basic Clin. Physiol. Pharmacol. 2019, 30. [Google Scholar] [CrossRef]
- Katz, R.J.; Roth, K.A.; Carroll, B.J. Acute and chronic stress effects on open field activity in the rat: Implications for a model of depression. Neurosci. Biobehav. Rev. 1981, 5, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Ren-ling, W.; Qi, P. Neural immune change in depression model mice. Acta Anat. Sin. 2018, 49, 281–287. [Google Scholar]
- Qiao-ling, H.; Hua-li, W. Comparison between two animal models of depression induced by corticosterone repeated injection and chronic unpredictable mild stress. Acta Anat. Sin. 2017, 48, 273–281. [Google Scholar]
- Shen, Z.; Xu, Y.; Jiang, X.; Wang, Z.; Guo, Y.; Pan, W.; Hou, J. Avicularin Relieves Depressive-Like Behaviors Induced by Chronic Unpredictable Mild Stress in Mice. Med. Sci. Monit. 2019, 25, 2777–2784. [Google Scholar] [CrossRef]
- Ning, J.; Lin-Xi, F. Antidepressant effects of the extract of Dendrobium nobile Lindl on chronic unpredictable mild stress-induced depressive mice. Acta Physiol. Sin. J. 2017, 69, 159–166. [Google Scholar]
- Lin, G.X.; Fang, Z.T. Effects of refined Xiaoyao Powder on ethology and expressions of P450scc in hippocampus of the depression rat. China J. Tradit. Chin. Med. Pharm. 2013, 28, 1253. [Google Scholar]
- Honglin, Q.; Jun, X.; Jiaqin, C. Aerobic Training Inhibits Hippocampal Inflammation by Activating the Hippocampus TLR4/miR223/NLRP3 Signaling Pathway Axis in Mice with CUMS Depression. China Sport Sci. J. 2019, 39, 39–50. [Google Scholar]
- Wen, L.; Jin, Y.; Li, L.; Sun, S.; Cheng, S.; Zhang, S.; Zhang, Y.; Svenningsson, P. Exercise prevents raphe nucleus mitochondrial overactivity in a rat depression model. Physiol. Behav. 2014, 132, 57–65. [Google Scholar] [CrossRef]
- Zhuang, P.C.; Tan, Z.N.; Jia, Z.Y.; Wang, B.; Grady, J.J.; Ma, X.M. Treadmill Exercise Reverses Depression Model-Induced Alteration of Dendritic Spines in the Brain Areas of Mood Circuit. Front. Behav. Neurosci. 2019, 13, 93. [Google Scholar] [CrossRef]
- Bangasser, D.A.; Curtis, A.; Reyes, B.A.; Bethea, T.T.; Parastatidis, I.; Ischiropoulos, H.; van Bockstaele, E.J.; Valentino, R.J. Sex differences in corticotropin-releasing factor receptor signaling and trafficking: Potential role in female vulnerability to stress-related psychopathology. Mol. Psychiatry 2010, 15, 877, 896–904. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, J.M.; Su, W.J.; Wang, B.; Jiang, C.L. Sex differences in depressive-like behaviour may relate to imbalance of microglia activation in the hippocampus. Brain Behav. Immun. 2019, 81, 188–197. [Google Scholar] [CrossRef]
- Crozet, P.; Margalha, L.; Confraria, A.; Rodrigues, A.; Martinho, C.; Adamo, M.; Elias, C.A.; Baena-González, E. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front Plant Sci. 2014, 5, 190. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. Sensing of energy and nutrients by AMP-activated protein kinase. Am. J. Clin. Nutr. J. 2011, 93, 891S–896S. [Google Scholar] [CrossRef] [Green Version]
- Oakhill, J.S.; Scott, J.W.; Kemp, B.E. AMPK functions as an adenylate charge-regulated protein kinase. Trends Endocrinol. Metab. 2012, 23, 125–132. [Google Scholar] [CrossRef]
- Rabinovitch, R.C.; Samborska, B.; Faubert, B.; Ma, E.H.; Gravel, S.P.; Andrzejewski, S.; Raissi, T.C.; Pause, A.; St-Pierre, J.; Jones, R.G. AMPK Maintains Cellular Metabolic Homeostasis through Regulation of Mitochondrial Reactive Oxygen Species. Cell Rep. 2017, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Jiang, Q.; Tuccitto, A.; Chan, D.; Alqawlaq, S.; Won, G.J.; Sivak, J.M. The AMPK-PGC-1alpha signaling axis regulates the astrocyte glutathione system to protect against oxidative and metabolic injury. Neurobiol. Dis. 2018, 113, 59–69. [Google Scholar] [CrossRef]
- Zhang, Q.; Liang, X.C. Effects of Mitochondrial Dysfunction via AMPK/PGC-1 alpha Signal Pathway on Pathogenic Mechanism of Diabetic Peripheral Neuropathy and the Protective Effects of Chinese Medicine. Chin. J. Integr. Med. 2019, 25, 386–394. [Google Scholar] [CrossRef]
- Yao, Y.; Rongjuan, G.; Huawei, S. Mitochondrial energy metabolism disorder in depression. J. Beijing Univ. Tradit. Chin. Med. 2019, 42, 602–606. [Google Scholar]
- Si-jun, Z.; Xiao-zhe, Z.; Huan, X. Regulation of seabuckthorn seed oil on metabolic pathway disturbances on rats exposed to chronic unpredictable mild stress. Chin. Tradit. Herbal Drugs J. 2017, 48, 2682–2690. [Google Scholar]
- Jou, S.H.; Chiu, N.Y.; Liu, C.S. Mitochondrial dysfunction and psychiatric disorders. Chang. Gung Med. J. 2009, 32, 370–379. [Google Scholar] [PubMed]
- Gardner, A.; Boles, R.G. Mitochondrial energy depletion in depression with somatization. Psychother. Psychosom. 2008, 77, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Zhang, J.; Hong, L.; Huang, W.; Dai, X.; Ye, Q.; Chen, X. Metformin ameliorates stress-induced depression-like behaviors via enhancing the expression of BDNF by activating AMPK/CREB-mediated histone acetylation. J. Affect. Disord. 2020, 260, 302–313. [Google Scholar] [CrossRef]
- Odaira, T.; Nakagawasai, O.; Takahashi, K.; Nemoto, W.; Sakuma, W.; Lin, J.R.; Tan-No, K. Mechanisms underpinning AMP-activated protein kinase-related effects on behavior and hippocampal neurogenesis in an animal model of depression. Neuropharmacology 2019, 150, 121–133. [Google Scholar] [CrossRef]
- Agudelo, L.Z.; Femenia, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal muscle PGC-1alpha1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Li, L.P.; Wang, Q.; Wu, Q.; Hu, H.H.; Zhang, M.; Fang, Y.Y.; Zhang, J.; Li, S.J.; Xiong, W.C.; et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [Google Scholar] [CrossRef]
- Kalkman, H.O. Novel Treatment Targets Based on Insights in the Etiology of Depression: Role of IL-6 Trans-Signaling and Stress-Induced Elevation of Glutamate and ATP. Pharmaceuticals 2019, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.Y.; Liu, J.; Zhou, J.; Lu, W.; Zhou, H.Y.; Long, L.H.; Hu, Z.L.; Ni, L.; Wang, Y.; Chen, J.G.; et al. AMPK Mediates Glucocorticoids Stress-Induced Downregulation of the Glucocorticoid Receptor in Cultured Rat Prefrontal Cortical Astrocytes. PLoS ONE 2016, 11, e0159513. [Google Scholar] [CrossRef]
- Koltai, E.; Hart, N.; Taylor, A.W.; Goto, S.; Ngo, J.K.; Davies, K.J.; Radak, Z. Age-associated declines in mitochondrial biogenesis and protein quality control factors are minimized by exercise training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R127–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da-lei, W.; Tong, W. Research on the Mechanism of Long-term Aerobic Exercise Reducing Oxidative Stress in the Hippocampus of Aging Rats. J. Xi’an Phys. Educ. Univ. 2014, 31, 350–354, 363. [Google Scholar]
- Yangwenjie, W.; Zhaojie, L.; Xuan, H.; Ying, Z. Effects of Aerobic Exercise Training on apelin-mediated Expressing of UCP3 in Skeletal Muscles of Mice. Chin. J. Sports Med. 2018, 37, 772–778. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Base Sequence | |
|---|---|
| GAPDH | F ACAGCAACAGGGTGGTGGAC R TTTGAGGGTGCAGCGAACTT |
| PGC-1α | F TGATGTGAATGACTTGGATACAGACA R GCTCATTGTTGTACTGGTTGGATATG |
| AMPK | F AAACCCACAGAAATCCAAACAC R CCTTCCATTCATAGTCCAACTG |
| Group | Before Experiment | After Modelling | After Exercise Intervention |
|---|---|---|---|
| CG | 41.68 ± 1.3503 | 53.47 ± 2.8909 | 61.37 ± 2.5877 |
| MG | 41.54 ± 1.4773 | 49.86 ± 1.4607 | 55.56 ± 1.2855 |
| ME | 42.13 ± 1.2811 | 49.16 ± 1.0023 | 58.4 ± 1.5269 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Tang, C.; Chen, X.; Ren, Z.; Qu, H.; Chen, R.; Tong, Z. Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway. Int. J. Environ. Res. Public Health 2020, 17, 2042. https://doi.org/10.3390/ijerph17062042
Luo J, Tang C, Chen X, Ren Z, Qu H, Chen R, Tong Z. Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway. International Journal of Environmental Research and Public Health. 2020; 17(6):2042. https://doi.org/10.3390/ijerph17062042
Chicago/Turabian StyleLuo, Jia, Changfa Tang, Xiaobin Chen, Zhanbing Ren, Honglin Qu, Rong Chen, and Zhen Tong. 2020. "Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway" International Journal of Environmental Research and Public Health 17, no. 6: 2042. https://doi.org/10.3390/ijerph17062042
APA StyleLuo, J., Tang, C., Chen, X., Ren, Z., Qu, H., Chen, R., & Tong, Z. (2020). Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway. International Journal of Environmental Research and Public Health, 17(6), 2042. https://doi.org/10.3390/ijerph17062042