Isolation and Identification of Chromium Reducing Bacillus Cereus Species from Chromium-Contaminated Soil for the Biological Detoxification of Chromium

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Isolation of the Strains
2.3. Morphological Observation of Strains
2.4. Phylogenetic Analysis Using 16S rDNA
2.5. Determining Cr(Ⅵ) in the Culture Medium
2.6. Conditions for Bacterial Growth
2.7. Immobilization of the Bacteria
2.8. Statistical Analysis
3. Results and Discussion
3.1. Isolation and Identification of Strains
3.2. Chromium Tolerance of the Isolated Strains
3.3. The Effect of pH on the Growth of the Isolated Strains
3.4. The Effect of Temperature on the Growth and Chromium Reduction of the Isolated Strains
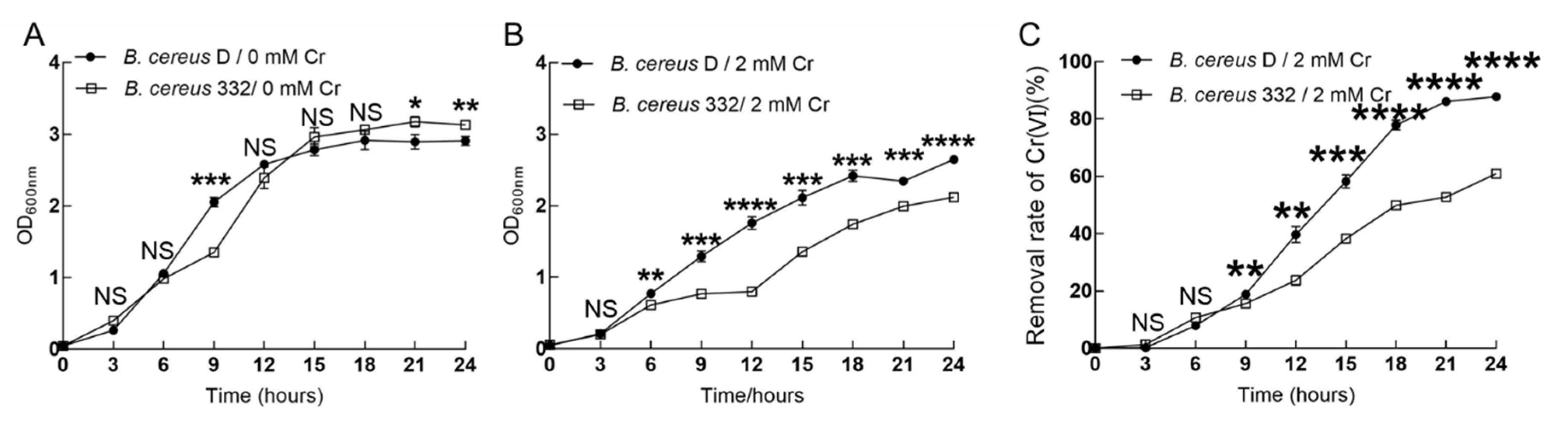
3.5. The Reducing Ability of the Isolated Strains
3.6. The Effect of Chemical Factors on the Ability of B. Cereus D and 332 in Reducing Chromium
3.7. Chromium Reduction of the Immobilized Strains
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kourtev, P.S.; Nakatsu, C.H.; Konopka, A. Inhibition of nitrate reduction by chromium (VI) in anaerobic soil microcosms. Appl. Environ. Microbiol. 2009, 75, 6249–6257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Hao, P.; Cheng, Y.; Ahmed, I.M.; Cao, F. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 162, 71–76. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, J.; Chen, R.; Yu, P.; Guo, S.; Wang, X. Adsorption and reduction of chromium(VI) from aqueous solution using polypyrrole/calcium rectorite composite adsorbent. Water Res. 2019, 160, 148–157. [Google Scholar] [CrossRef]
- Wang, Z.; Lin, F.; Huang, L.; Chang, Z.; Yang, B.; Liu, S.; Zheng, M.; Lu, Y.; Chen, J. Cyclodextrin functionalized 3D-graphene for the removal of Cr(VI) with the easy and rapid separation strategy. Environ. Pollut. 2019, 254, 112854. [Google Scholar] [CrossRef] [PubMed]
- Goswami, L.; Mukhopadhyay, R.; Bhattacharya, S.S.; Das, P.; Goswami, R. Detoxification of chromium-rich tannery industry sludge by Eudrillus eugeniae: Insight on compost quality fortification and microbial enrichment. Bioresour. Technol. 2018, 266, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Bibi, I.; Niazi, N.K.; Choppala, G.; Burton, E.D. Chromium(VI) removal by siderite (FeCO3) in anoxic aqueous solutions: An X-ray absorption spectroscopy investigation. Sci. Total Environ. 2018, 640–641, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Oruko Ongon’g, R.; Edokpayi, J.N.; Msagati, T.A.M.; Tavengwa, N.T.; Ijoma, G.N.; Odiyo, J.O. The potential health risk associated with edible vegetables grown on Cr(VI) polluted soils. Int. J. Environ. Res. Public Health 2020, 17, 470. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, H.; Wu, X.; Zhang, T.; Shen, K.; Li, L.; Peng, Y.; Mehmood, K.; Zhou, D. Metabonomic analysis of the hepatic injury suffer from hexavalent chromium poisoning in broilers. Environ. Sci. Pollut. Res. 2019, 26, 18181–18190. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Zhu, Y.; Zhang, Y. Interaction of chromium(III) or chromium(VI) with catalase and its effect on the structure and function of catalase: An In Vitro study. Food Chem. 2018, 244, 378–385. [Google Scholar] [CrossRef]
- Ancona, V.; Campanale, C.; Tumolo, M.; De Paola, D.; Ardito, C.; Volpe, A.; Uricchio, V.F. Enhancement of chromium (VI) reduction in microcosms amended with lactate or yeast extract: A laboratory-scale Study. Int. J. Environ. Res. Public Health 2020, 17, 704. [Google Scholar] [CrossRef] [Green Version]
- Aranda-García, E.; Cristiani-Urbina, E. Effect of pH on hexavalent and total chromium removal from aqueous solutions by avocado shell using batch and continuous systems. Environ. Sci. Pollut. Res. 2019, 26, 3157–3173. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chai, L.; Yang, W.; Wang, H.; Zhang, L. Two-dimensional titanium carbides (Ti3C2Tx) functionalized by poly(m-phenylenediamine) for efficient adsorption and reduction of hexavalent chromium. Int. J. Environ. Res. Public Health 2019, 17, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlosser, P.M.; Sasso, A.F. A revised model of ex-vivo reduction of hexavalent chromium in human and rodent gastric juices. Toxicol. Appl. Pharmacol. 2014, 280, 352–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modenes, A.N.; de Oliveira, A.P.; Espinoza-Quinones, F.R.; Trigueros, D.E.G.; Kroumov, A.D.; Bergamasco, R. Study of the involved sorption mechanisms of Cr(VI) and Cr(III) species onto dried Salvinia auriculata biomass. Chemosphere 2017, 172, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Murugavelh, S.; Mohanty, K. Isolation, identification and characterization of Cr(VI) reducing Bacillus cereus from chromium contaminated soil. Chem. Eng. J. 2013, 230, 1–9. [Google Scholar] [CrossRef]
- Poonkuzhali, K.; Rajeswari, V.; Saravanakumar, T.; Viswanathamurthi, P.; Park, S.M.; Govarthanan, M.; Sathishkumar, P.; Palvannan, T. Reduction of hexavalent chromium using Aerva lanata L.: Elucidation of reduction mechanism and identification of active principles. J. Hazard. Mater. 2014, 272, 89–95. [Google Scholar] [CrossRef]
- Liang, M.; Xu, S.; Zhu, Y.; Chen, X.; Deng, Z.; Yan, L.; He, H. Preparation and characterization of Fe-Mn binary oxide/mulberry stem biochar composite adsorbent and adsorption of Cr(VI) from aqueous solution. Int. J. Environ. Res. Public Health 2020, 17, 676. [Google Scholar] [CrossRef] [Green Version]
- Raptis, S.; Gasparatos, D.; Economou-Eliopoulos, M.; Petridis, A. Chromium uptake by lettuce as affected by the application of organic matter and Cr(VI)-irrigation water: Implications to the land use and water management. Chemosphere 2018, 210, 597–606. [Google Scholar] [CrossRef]
- Xia, Q.; Huang, B.; Yuan, X.; Wang, H.; Wu, Z.; Jiang, L.; Xiong, T.; Zhang, J.; Zeng, G.; Wang, H. Modified stannous sulfide nanoparticles with metal-organic framework: Toward efficient and enhanced photocatalytic reduction of chromium (VI) under visible light. J. Colloid Interface Sci. 2018, 530, 481–492. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, Y.; Zhang, X.; Liu, P.; Ren, J.; Wu, G.; Zhang, Y.; Chen, Y.; Li, X. A Bacillus subtilis strain can reduce hexavalent chromium to trivalent and an nfrA gene is involved. Int. Biodeterior. Biodegrad. 2015, 97, 90–96. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, Z.; Cheng, Y.; Pan, D.; Pan, X.; Yu, M.; Pan, Z.; Lin, Z.; Guan, X.; Wu, Z. Cr(VI) uptake mechanism of Bacillus cereus. Chemosphere 2012, 87, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, B.; Diao, M.; Shi, J.; Jiang, Y.; Cheng, Y.; Liu, H. Enhancement of synchronous bio-reductions of vanadium (V) and chromium (VI) by mixed anaerobic culture. Environ. Pollut. 2018, 242, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wu, K.; Khan, A.; Jiang, Y.; Ling, Z.; Liu, P.; Chen, Y.; Tao, X.; Li, X. A novel Pseudomonas gessardii strain LZ-E simultaneously degrades naphthalene and reduces hexavalent chromium. Bioresour. Technol. 2016, 207, 370–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- More, A.G.; Gupta, S.K. Evaluation of chromium removal efficiency at varying operating conditions of a novel bioelectrochemical system. Bioprocess. Biosyst. Eng. 2018, 41, 1547–1554. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, Q.; Chen, W.; Teng, B. Removal of hexavalent chromium in tannery wastewater by Bacillus cereus. Can. J. Microbiol. 2012, 58, 23–28. [Google Scholar] [CrossRef]
- Zeng, J.; Gou, M.; Tang, Y.Q.; Li, G.Y.; Sun, Z.Y.; Kida, K. Effective bioleaching of chromium in tannery sludge with an enriched sulfur-oxidizing bacterial community. Bioresour. Technol. 2016, 218, 859–866. [Google Scholar] [CrossRef]
- Jiang, B.; Zhu, D.; Song, Y.; Zhang, D.; Liu, Z.; Zhang, X.; Huang, W.E.; Li, G. Use of a whole-cell bioreporter, Acinetobacter baylyi, to estimate the genotoxicity and bioavailability of chromium(VI)-contaminated soils. Biotechnol. Lett. 2015, 37, 343–348. [Google Scholar] [CrossRef]
- Fonseca, B.; Pazos, M.; Tavares, T.; Sanroman, M.A. Removal of hexavalent chromium of contaminated soil by coupling electrokinetic remediation and permeable reactive biobarriers. Environ. Sci. Pollut. Res. 2012, 19, 1800–1808. [Google Scholar] [CrossRef] [Green Version]
- Goulhen, F.; Gloter, A.; Guyot, F.; Bruschi, M. Cr(VI) detoxification by Desulfovibrio vulgaris strain Hildenborough: Microbe-metal interactions studies. Appl. Microbiol. Biotechnol. 2006, 71, 892–897. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, N.; Nawani, N.N.; Jass, J.; Hossain, K.; Saud, Z.A.; Saha, A.K.; Ghosh, S.; Olsson, B.; Mandal, A. Bioremediation of hexavalent chromium (VI) by a soil-borne bacterium, Enterobacter cloacae B2-DHA. J. Environ. Sci. Health Part A Toxic/Hazard. Subst. Environ. Eng. 2015, 50, 1136–1147. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Ai, W.; Dong, X. High-quality draft genome sequence of Leucobacter sp. strain G161, a distinct and effective chromium reducer. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, N.K.; Donmez, G. Environmental conditions affecting exopolysaccharide production by Pseudomonas aeruginosa, Micrococcus sp., and Ochrobactrum sp. J. Hazard. Mater. 2008, 154, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Opperman, D.J.; Piater, L.A.; van Heerden, E. A novel chromate reductase from Thermus scotoductus SA-01 related to old yellow enzyme. J. Bacteriol. 2008, 190, 3076–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold spring harbor Laboratory Press: Cold spring harbor, NY, USA, 1982; pp. 895–909. [Google Scholar]
- Todd, C.D.; Reyes-Batlle, M.; Martin-Navarro, C.M.; Dorta-Gorrin, A.; Lopez-Arencibia, A.; Martinez-Carretero, E.; Pinero, J.E.; Valladares, B.; Lindo, J.F.; Lorenzo-Morales, J. Isolation and genotyping of Acanthamoeba strains from soil sources from Jamaica, West Indies. J. Eukaryot. Microbiol. 2015, 62, 416–421. [Google Scholar] [CrossRef]
- Cuesta, G.; Garcia-de-la-Fuente, R.; Abad, M.; Fornes, F. Isolation and identification of actinomycetes from a compost-amended soil with potential as biocontrol agents. J. Environ. Manag. 2012, 95, S280–S284. [Google Scholar] [CrossRef]
- Prakash, D.; Nawani, N.N. A rapid and improved technique for scanning electron microscopy of actinomycetes. J. Microbiol. Methods 2014, 99, 54–57. [Google Scholar] [CrossRef]
- Lee, J.T.; Chow, K.L. SEM sample preparation for cells on 3D scaffolds by freeze-drying and HMDS. Scanning 2012, 34, 12–25. [Google Scholar] [CrossRef]
- Fang, J.; Wang, C.; Li, Y.; Zhao, Z.; Mei, L. Comparison of bacterial adhesion to dental materials of polyethylene terephthalate (PET) and polymethyl methacrylate (PMMA) using atomic force microscopy and scanning electron microscopy. Scanning 2016, 38, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Josephine, A.M.; Paul, A.G.; Mizer, H.E. Laboratory Manual and Workbook in Microbiology; McGraw-Hill: New York, NY, USA, 2007; pp. 20–100. [Google Scholar]
- Sheng-Mei, Y.E.; Gao, W. Improvement of Bacteria’s Spore Staining. J. Microbiol. 2011, 31, 106–109. [Google Scholar] [CrossRef] [Green Version]
- Suzuk Yildiz, S.; Kaskatepe, B.; Altinok, S.; Cetin, M.; Karagoz, A.; Savas, S. Comparison of MALDI-TOF and 16S rRNA methods in identification of viridans group streptococci. Mikrobiyol. Bul. 2017, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Duffy, G.; Maguire, I.; Heery, B.; Gers, P.; Ducree, J.; Regan, F. ChromiSense: A colourimetric lab-on-a-disc sensor for chromium speciation in water. Talanta 2018, 178, 392–399. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zheng, M.; Ma, T.; Ni, J. Nitrite interference and elimination in diphenylcarbazide (DPCI) spectrophotometric determination of hexavalent chromium. Water Sci. Technol. 2015, 72, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, S.M.; Shemirani, F. Selective and sensitive speciation analysis of Cr(VI) and Cr(III) in water samples by fiber optic-linear array detection spectrophotometry after ion pair based-surfactant assisted dispersive liquid-liquid microextraction. J. Hazard. Mater. 2013, 254–255, 134–140. [Google Scholar] [CrossRef]
- Lace, A.; Ryan, D.; Bowkett, M.; Cleary, J. Chromium monitoring in water by colorimetry using optimised 1,5-diphenylcarbazide method. Int. J. Environ. Res. Public Health 2019, 16, 1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.B.; Li, Y.Q.; Lin, J.Q.; Pang, X.; Liu, X.M.; Liu, B.Q.; Wang, R.; Zhang, C.J.; Wu, Y.; Lin, J.Q.; et al. The two-component system RsrS-RsrR regulates the tetrathionate intermediate pathway for thiosulfate oxidation in Acidithiobacillus caldus. Front. Microbiol. 2016, 7, 1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Zhang, Y.; Tu, B. Immobilization of ammonia-oxidizing bacteria by polyvinyl alcohol and sodium alginate. Braz. J. Microbiol. 2017, 48, 515–521. [Google Scholar] [CrossRef]
- Maqbool, F.; Wang, Z.; Xu, Y.; Zhao, J.; Gao, D.; Zhao, Y.G.; Bhatti, Z.A.; Xing, B. Rhizodegradation of petroleum hydrocarbons by Sesbania cannabina in bioaugmented soil with free and immobilized consortium. J. Hazard. Mater. 2012, 237–238, 262–269. [Google Scholar] [CrossRef]
- Ruan, B.; Wu, P.; Chen, M.; Lai, X.; Chen, L.; Yu, L.; Gong, B.; Kang, C.; Dang, Z.; Shi, Z.; et al. Immobilization of Sphingomonas sp. GY2B in polyvinyl alcohol-alginate-kaolin beads for efficient degradation of phenol against unfavorable environmental factors. Ecotoxicol. Environ. Saf. 2018, 162, 103–111. [Google Scholar] [CrossRef]
- Lin, H.; Chen, Z.; Megharaj, M.; Naidu, R. Biodegradation of TNT using Bacillus mycoides immobilized in PVA–sodium alginate–kaolin. Appl. Clay Sci. 2013, 83–84, 336–342. [Google Scholar] [CrossRef]
- Balows, A. Bergey’s Manual of Determinative Bacteriology. Eighth Edition. Am. J. Public Health 1975, 65, 315. [Google Scholar] [CrossRef] [Green Version]
- Nemecek, J.; Pokorny, P.; Lacinova, L.; Cernik, M.; Masopustova, Z.; Lhotsky, O.; Filipova, A.; CajthamL, T. Combined abiotic and biotic In-Situ reduction of hexavalent chromium in groundwater using nZVI and whey: A remedial pilot test. J. Hazard. Mater. 2015, 300, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Němeček, J.; Pokorný, P.; Lhotský, O.; Knytl, V.; Najmanová, P.; Steinová, J.; Černík, M.; Filipová, A.; Filip, J.; CajthamL, T. Combined nano-biotechnology for in-situ remediation of mixed contamination of groundwater by hexavalent chromium and chlorinated solvents. Sci. Total Environ. 2016, 563–564, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Pichtel, J.; Pichtel, T.M. Comparison of solvents for ex situ removal of chromium and lead from contaminated soil. Environ. Eng. Sci. 1997, 14, 97–104. [Google Scholar] [CrossRef]
- Sun, Y.; Guan, F.; Yang, W.; Wang, F. Removal of chromium from a contaminated soil using oxalic acid, citric acid, and hydrochloric acid: Dynamics, mechanisms, and concomitant removal of non-targeted metals. Int. J. Environ. Res. Public Health 2019, 16, 2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berar, I.M.; Micle, V.; Urs, A.M. Research on contaminated soils from baia mare area and their remediation by In Situ bio-leaching technique. Proenvironment/Promediu 2011, 4, 208–211. [Google Scholar]
- Camargo, F.A.; Okeke, B.C.; Bento, F.M.; Frankenberger, W.T. In Vitro reduction of hexavalent chromium by a cell-free extract of Bacillus sp. ES 29 stimulated by Cu2+. Appl. Microbiol. Biotechnol. 2003, 62, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, R.; Abhipsa, S.; Rohit, B.; Ligy, P.; Chandraraj, K. Reduction of Cr(VI) by a Bacillus sp. Biotechnol. Lett. 2006, 28, 247–252. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Results | Tests | Results | ||
|---|---|---|---|---|---|
| B. cereus D | B. cereus 332 | B. cereus D | B. cereus 332 | ||
| Morphological Tests | |||||
| Clone shape | Round | Round | Pigment | - | - |
| Margin | Irregular | Irregular | Gram-reaction | + | + |
| Surface | Rough | Rough | Spore | + | + |
| Opacity | Opaque | Opaque | Cell shape | Rods | Rods |
| Biochemical Tests | |||||
| MR-VP test | + | + | Motive test | + | + |
| Lysozyme | + | + | Nitrate test | + | + |
| Lecithin | + | + | Acid production | + | + |
| Hemolysis | + | + | Protein crystal | - | - |
| Root shape growth | + | + | Casein decomposition | + | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.-h.; Gao, X.-y.; Li, C.; Yang, C.-l.; Fu, C.-a.; Liu, J.; Wang, R.; Chen, L.-x.; Lin, J.-q.; Liu, X.-m.; et al. Isolation and Identification of Chromium Reducing Bacillus Cereus Species from Chromium-Contaminated Soil for the Biological Detoxification of Chromium. Int. J. Environ. Res. Public Health 2020, 17, 2118. https://doi.org/10.3390/ijerph17062118
Li M-h, Gao X-y, Li C, Yang C-l, Fu C-a, Liu J, Wang R, Chen L-x, Lin J-q, Liu X-m, et al. Isolation and Identification of Chromium Reducing Bacillus Cereus Species from Chromium-Contaminated Soil for the Biological Detoxification of Chromium. International Journal of Environmental Research and Public Health. 2020; 17(6):2118. https://doi.org/10.3390/ijerph17062118
Chicago/Turabian StyleLi, Ming-hao, Xue-yan Gao, Can Li, Chun-long Yang, Chang-ai Fu, Jie Liu, Rui Wang, Lin-xu Chen, Jian-qiang Lin, Xiang-mei Liu, and et al. 2020. "Isolation and Identification of Chromium Reducing Bacillus Cereus Species from Chromium-Contaminated Soil for the Biological Detoxification of Chromium" International Journal of Environmental Research and Public Health 17, no. 6: 2118. https://doi.org/10.3390/ijerph17062118
APA StyleLi, M. -h., Gao, X. -y., Li, C., Yang, C. -l., Fu, C. -a., Liu, J., Wang, R., Chen, L. -x., Lin, J. -q., Liu, X. -m., Lin, J. -q., & Pang, X. (2020). Isolation and Identification of Chromium Reducing Bacillus Cereus Species from Chromium-Contaminated Soil for the Biological Detoxification of Chromium. International Journal of Environmental Research and Public Health, 17(6), 2118. https://doi.org/10.3390/ijerph17062118