1. Introduction
Skeletal muscle contractions consist of dynamic (e.g., concentric and eccentric contractions) and static muscle actions (e.g., isometric contraction). Based on the load type applied on the muscle, the isometric muscle actions can occur during two different tasks: contracting muscles against an immovable object (force task), and holding still against an external load (position task). Interestingly, even with the similar mechanical torque requirements for both isometric tasks, it is well-established that the fatiguing isometric tasks show differences in task failure time [
1], myoelectric activities, perceived effort [
2,
3], and spinal reflex activities [
4]. In addition, the variation of fatigue responses in both tasks was also dependent on contraction intensities and limb positions [
5,
6,
7]. Thus, a variety of factors such as the modulations of the motor unit (MU) activities (discharging and recruitment) [
8] can contribute to the task-related differences in skeletal muscle fatigue [
9].
With the difficulty to identify a single major contributing factor for the task-related differences in muscle fatigue, an alternative way is to study motor control strategies for these tasks directly, during the non-fatiguing condition. For example, by using surface electromyography (EMG) in both sexes, Garner et al. [
10] found no difference in the soleus muscle EMG amplitude between the force and position tasks during submaximal plantarflexion across a range of contraction intensities (20–50% of maximal ground reaction force). However, Buchanan and Lloyd [
11] reported differential coactivation patterns of EMG amplitude for the synergistic muscles for both non-fatiguing elbow flexion and extension force and position tasks, regardless of sex, suggesting two distinct motor control strategies being used in these tasks. In addition to the global EMG activity, examining MU firing properties may shed more information on understanding the underlying mechanisms of the task-related differences. For example, Mottram et al. [
8] examined single MU behavior from biceps brachii in fifteen men using a fine-wire bipolar electrode during the force and position isometric sustained fatiguing contractions at low target force. The results showed that overall MU behaviors were similar regarding MU mean firing rates (MFR) and recruitment threshold (RT) between two tasks. However, the number of recruited MUs was greater during the position task than the force task, suggesting greater excitatory input from the central nervous system (CNS) to motoneuron pool for the position task when compared to the force task [
8]. A more recent study compared single MU activity during fatiguing contraction between these two tasks in both sexes, and found that the magnitude of the decline in the MFR was greater in the position task when compared to the force task. In addition, the decline in the RT of detected MUs was present during the position task, but not during the force task. The authors suggest that these changes may involve differential synaptic mechanisms [
12]. Thus, despite similar mechanical requirements between two isometric tasks during non-fatiguing contractions, these results from previous studies suggested that two distinct mechanisms of MU control strategies, such as differential MU activities and/or excitatory and inhibitory input to the CNS, may exist in these tasks.
For more than four decades, the development in surface EMG decomposition algorithms has allowed researchers to decompose the surface EMG signals into MU action potential trains [
13,
14,
15,
16]. With these techniques, previous studies examined MU activities during submaximal, maximal, or fatiguing contractions using different muscles. Specifically, they found that a negative linear relationship between RT and MFR of the active MUs during a voluntary contraction, which was described as the “onion-skin” scheme [
12]. This indicates that the low-threshold MUs tend to have higher firing rates than those of the high-threshold MUs, during the voluntary contractions [
5,
6,
7]. This relationship also suggests an important neural strategy on how the CNS controls the motoneuron pool: the slope of the linear regression line indicates the overall motor control scheme described as the “operating point” of the motoneuron pools [
17]. Specifically, when the force output (contraction intensity) increases, the slope and y-intercept of the linear regression line between MU RT and MFR gradually become less negative and higher, respectively, suggesting the recruitment of the additional high-threshold MUs and the increased firing rates of the active MUs to meet the force production demand. On the other hand, the relationship between MU RT and derecruitment threshold (DT) is positive, suggesting that higher threshold MUs generally tend to be derecruited at higher force levels than where they were recruited, but lower threshold MUs tend to be derecruited at lower force level [
17,
18]. This MU RT vs. DT relationship has been used in a previous study to examine the fatiguing effect on the motor control strategy [
19]. Thus, the EMG decomposition algorithm, which examines the active MU action potential trains during voluntary isometric contractions, can serve as a useful tool to investigate MU control mechanisms for non-fatiguing isometric tasks.
Considering previous studies [
8,
12] have found distinct MU control strategies between the force and position tasks during sustained fatiguing contractions regardless of sex, it is possible to observe two different mechanisms during non-fatiguing force and position tasks. Specifically, the direct comparison of MU control strategies between the non-fatiguing force and position isometric tasks has not been examined, and varying the MU control strategies will potentially influence MU activities and the mechanisms involved. The potential presence of two distinct mechanisms in the two different tasks may provide a better understanding of the underlying mechanism(s) leading to the task differences. However, the knowledge regarding these two tasks per se remains to be poor. Therefore, the primary purpose of this study was to determine the MU control strategies using linear regression analysis during the force and position tasks at 40% and 70% contraction intensities in human biceps brachii muscle. Based on a previous study [
20] that reported biceps brachii muscle replying more on the strategy of recruiting MU at high force level (around up to 88%), 40% and 70% contraction intensities were selected to examine MU control strategies between two different isometric tasks. In addition, the surface EMG amplitudes of biceps and triceps brachii were also monitored. Since it has been shown that the EMG amplitude did not differ between two tasks [
10], we expected to see a similar finding regarding the EMG amplitude between two tasks. However, considering the different excitatory inputs to the motoneuron pool between two different tasks [
21,
22], the relationship between MU RT and MFR, as well as the relationship between MU RT and DT between two tasks may differ.
3. Results
3.1. Test–Retest Reliability
The intraclass correlation coefficient model (3, 1) (ICC
3,1) [
27] for the elbow flexors’ maximal isometric strength during three visits (familiarization visit vs. force-task visit vs. position-task visit) was reliable (r = 0.97), with no significant difference among the three visits (F = 0.25,
p = 0.782).
3.2. Isometric Force and EMG Amplitude during Submaximal Isometric Trapezoid Contractions
The paired sample t-test indicated no significant difference for maximal isometric strength between two experimental visits (force-task visit vs. position-task visit = 299.3 ± 133.7 N vs. 299.4 ± 132.4 N, p = 0.987, d = 0.001). For the force error CoV, the 3-way ANOVA only showed a 2-way phase × intensity interaction (F = 5.45, p = 0.032, partial = 0.280), and a main effect for phase (F = 78.176, p < 0.001, partial = 0.848). In addition, based on the result of the 2-way ANOVA, the force steadiness (force CoVs) during the mid-6-s of the plateau regions of the two isometric tasks were also not significantly different (F = 2.79, p = 0.117, partial = 0.166).
For the biceps EMG amplitude, the results from the 2-way ANOVA showed no interaction (F = 0.25, p = 0.621, partial = 0.014) or main effect for condition (F = 1.43, p = 0.248, partial = 0.073), but there was a main effect for intensity (F = 75.22, p < 0.001, partial = 0.807), indicating greater EMG amplitude during 70% MVIC, when compared to 40% MVIC (tasks merged, 40% MVIC vs. 70% MVIC = 57.5 ± 19.8% vs. 105.1 ± 30.2%, p < 0.0001, d = 1.86). For the normalized triceps brachii EMG amplitude, the 2-way ANOVA showed no interaction (F = 1.19, p = 0.290, partial = 0.062) or main effect for condition (F = 0.22, p = 0.644, partial = 0.012), but there was a main effect for intensity (F = 31.94, p < 0.001, partial = 0.640). After collapsing the across condition, 70% MVIC contraction intensity generally showed a greater EMG amplitude (both tasks merged) than 40% MVIC (40% MVIC vs. 70% MVIC = 12.4 ± 8.6% vs. 23.5 ± 16.6%, p < 0.0001, d = 1.88).
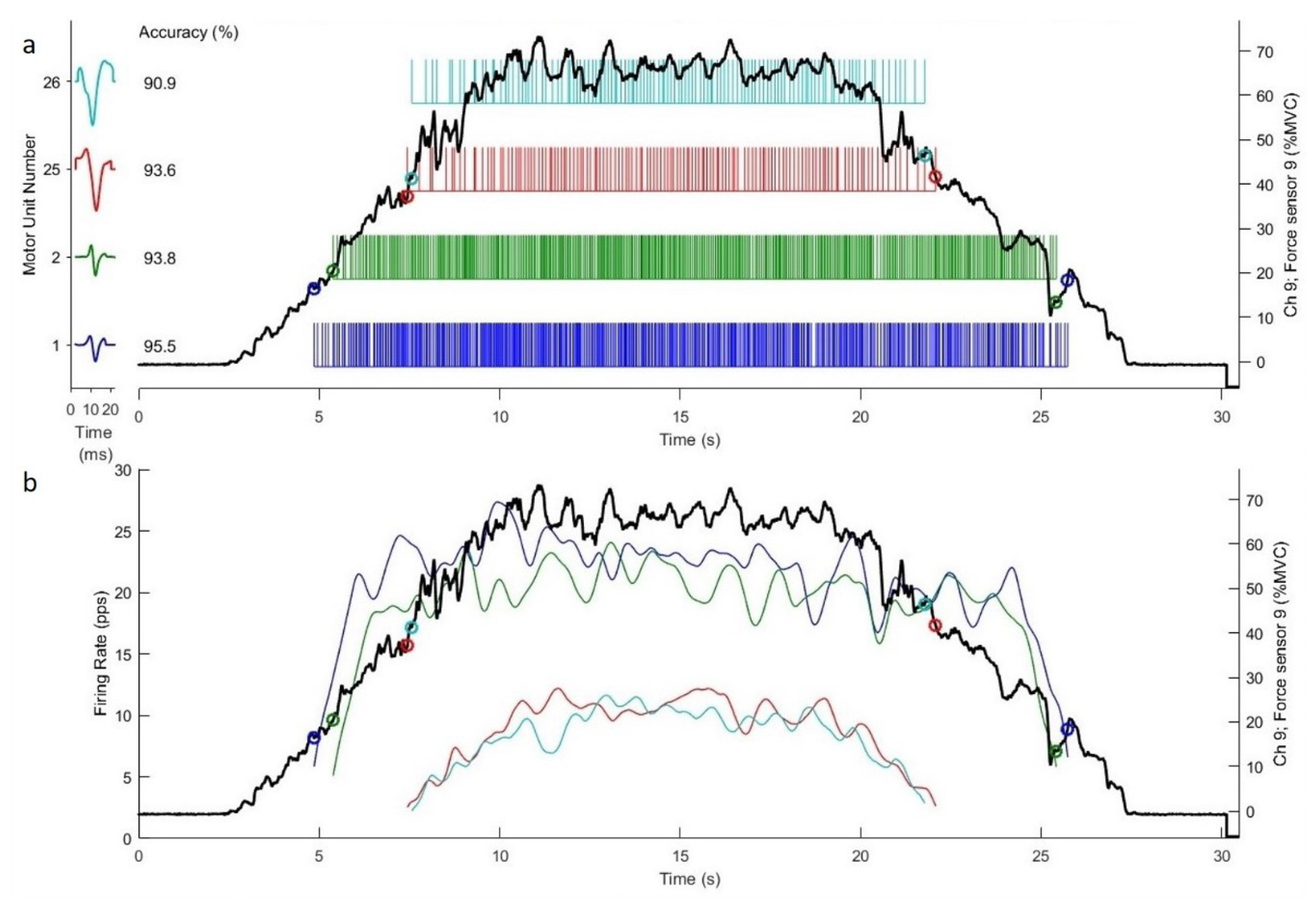
3.3. Motor Unit Firing Properties
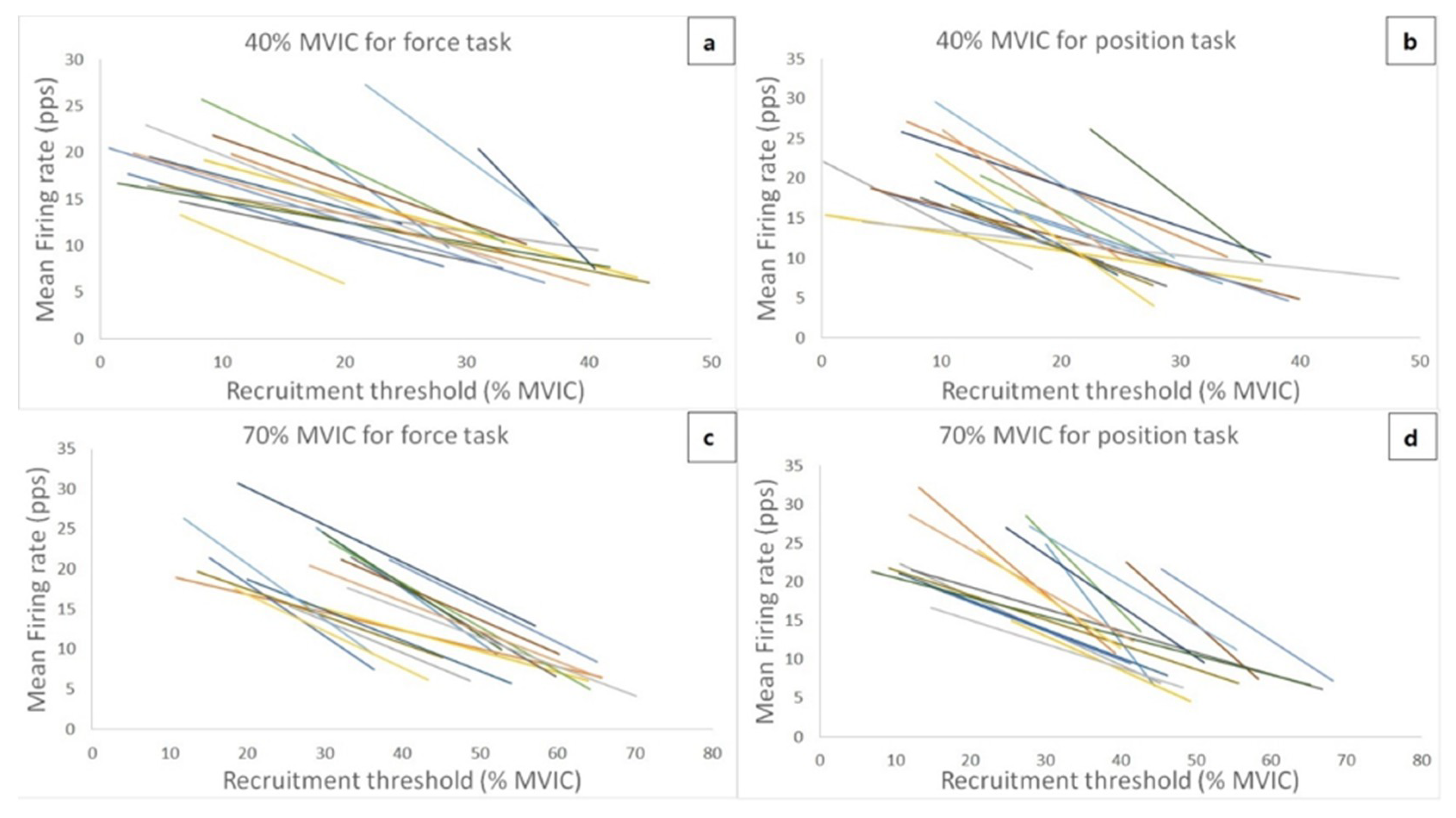
Figure 3 shows the theoretical regression lines for the relationship between the MU RT and MFR for all 17 subjects during 40% and 70% MVIC force task and position task isometric contraction. Due to the low-quality data and exclusion criterion, two subjects were excluded for linear regression analysis. Thus, the linear regression for the relationship between MU RT and MFR was analyzed from seventeen subjects. The ranges of recruitment thresholds of recorded MUs were similar between the two different tasks at 40% (force task vs. position task = 8.37 ± 7.92% to 34.91 ± 6.86% vs. 8.70 ± 5.51% to 31.63 ± 7.45%) and 70% MVIC contraction intensities (force task vs. position task = 24.41 ± 8.48% to 55.25 ± 10.47% vs. 20.40 ± 11.26% to 50.51 ± 9.56%). For the linear slope coefficients, the 2-way ANOVA showed no significant interaction (F = 0.05,
p = 0.826, partial
= 0.003) or main effect for intensity (F = 1.71,
p = 0.209, partial
= 0.097), but there was a significant main effect for condition (F = 6.73,
p = 0.020, partial
= 0.296). The follow-up pairwise comparison indicated that the slope coefficient was significantly more negative (intensity merged) during the position task, when compared to the force task (force task vs. position task = -0.48 ± 0.24 pps/%MVIC vs. −0.63 ± 0.27 pps/%MVIC,
p = 0.010,
d = 0.588). For the y-intercepts, the 2-way ANOVA did not show a significant interaction (F = 1.08,
p = 0.314, partial
= 0.063) or main effect for condition (F = 1.90,
p = 0.187, partial
= 0.106). However, there was a significant main effect for intensity (F = 13.56,
p = 0.002, partial
= 0.459), indicating that the y-intercepts (condition merged) were higher during the 70% than 40% MVIC contraction intensity (40% MVIC vs. 70% MVIC = 26.75 ± 10.83 pps vs. 34.75 ± 11.03 pps,
p < 0.001,
d = 0.734).
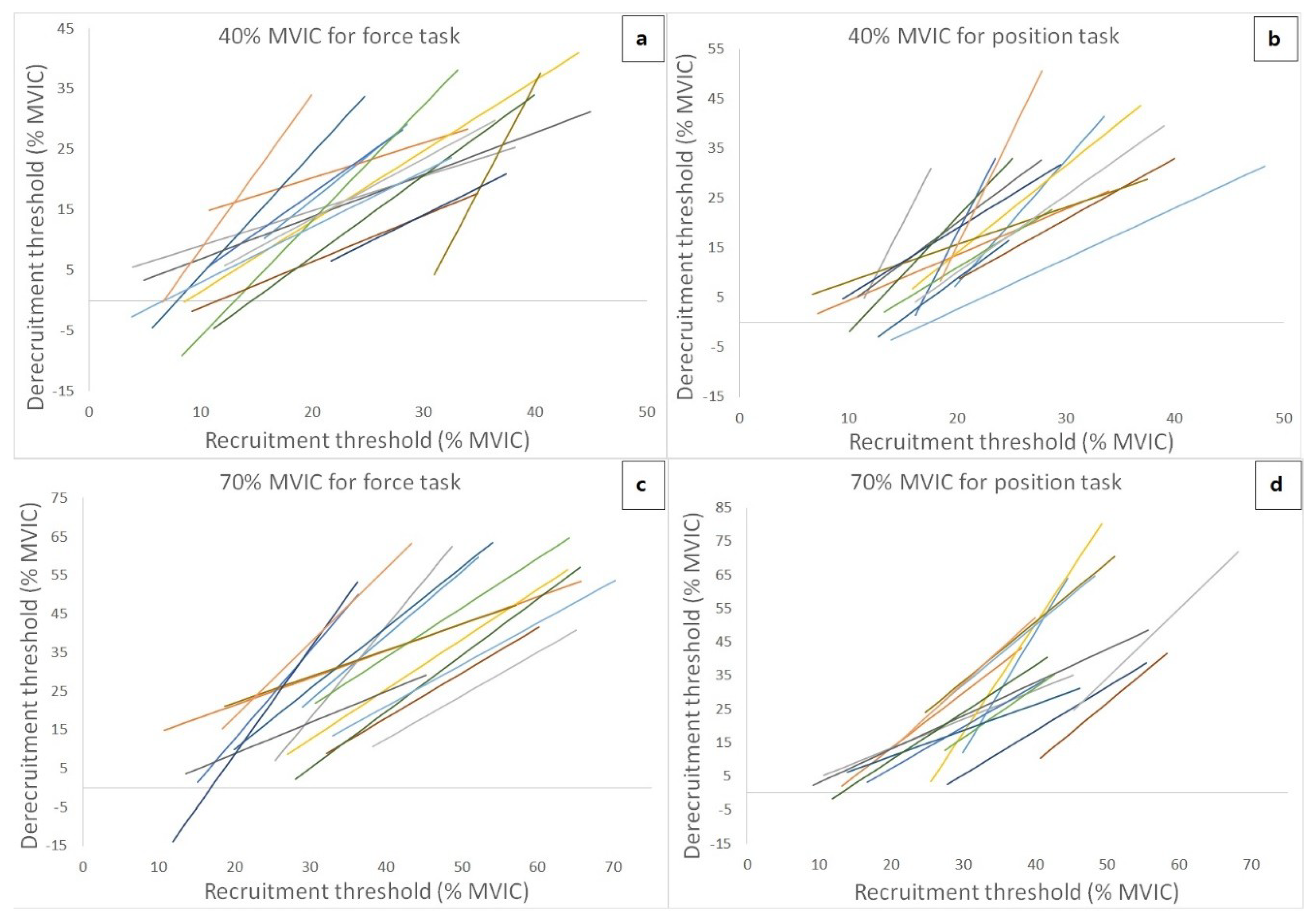
Figure 4 shows the theoretical regression lines for the relationship between the MU RT and DT for each of 15 subjects during 40% and 70% MVIC force task and position task isometric contraction. Four of the subjects were excluded for linear regression of 40% and 70% MVIC of trapezoid contracions due to MU data exclusion criteria. Thus, fifteen subjects were collected MU data for 40% and 70% MVIC trapezoid contractions. The number of MUs recorded, the number of MUS with RT < DT and RT > DT, and cross-over point (RT MVIC%) from the relationship between the MU RT and DT during both isometric tasks are presented in
Table 1 (40% MVIC contraction intensity) and
Table 2 (70% MVCI contraction intensity) for each subject. The cross-over point for force task and position task at 40% MVIC were varied with an average of 23.56% ± 6.97% and 19.60% ± 3.54%, respectively. In addition, the cross-over point varied with an average of 41.14% ± 14.34% and 39.25% ± 9.82% for force task and position task at 70% MVIC, respectively. The results from the 2-way ANOVA showed that there was no significant interaction (F = 0.34,
p = 0.571, partial
= 0.024) or main effect for intensity (F = 1.04,
p = 0.325, partial
= 0.069), but there was a significant main effect for condition (F = 4.81,
p = 0.046, partial
= 0.256) for the linear slope coefficient. The follow-up pairwise comparison indicated that the slope coefficient was significantly more positive (intensity merged) during the position task, when compared to the force task (force task vs. position task = 1.36 ± 0.67 pps/%MVIC vs. 1.70 ± 0.74 pps/%MVIC,
p = 0.023, d = 0.486). For the y-intercepts, the 2-way ANOVA showed that there was no significant interaction (F = 0.78,
p = 0.392, partial
= 0.053) or main effect for condition (F = 1.88,
p = 0.192, partial
= 0.118). However, there was a significant main effect for intensity (F = 5.16,
p = 0.039, partial
= 0.269), indicating that the y-intercepts (conditions merged) were lower during the 70% than 40% MVIC contraction intensity (40% MVIC vs. 70% MVIC = −15.45 ± 20.02 pps vs. −29.24 ± 23.45 pps,
p = 0.020,
d = 0.632).
4. Discussion
The main purpose of this study was to examine MU control strategies between a force and position task for the elbow flexors. The linear regression analysis was specifically used to examine the relationship between the MU RT and MFR, as well as the relationship between MU RT and DT at two contraction intensities (40% and 70% MVICs). In addition, global surface EMG amplitude was also examined for both agonist (biceps brachii) and antagonist (triceps brachii) muscles. The main findings of this study were: (1) The surface EMG amplitude did not differ between two tasks at both intensities for both agonist and antagonist muscles; and (2) The linear slope coefficients of the MU RT vs. MFR as well as the RT vs. DT relationships were significantly different between the two tasks (contraction intensities merged).
The elbow flexion isometric strength values were not different between the two experimental visits. The results of the agonist and antagonist EMG amplitude responses during the two tasks were generally in agreement with previous studies [
10,
11] that compared the EMG amplitude between these tasks. For example, Garner et al. [
10] reported no difference in the soleus muscle EMG amplitude between two tasks during non-fatiguing plantarflexions. In addition, Buchanan and Lloyd [
11] examined the EMG amplitude of biceps brachii, brachialis, brachioradialis, and triceps brachii during force and position elbow flexion tasks with relatively low loads (up to 40% of the maximal strength). It was reported that there was large individual variation (six out of nine individuals showed greater activation level during position task than force task) for the biceps brachii EMG amplitude between the two tasks, and no difference in antagonist muscle (triceps) EMG amplitude was shown [
11]. It is worth mentioning that in the current study, the condition factor imposed a medium effect on the biceps EMG responses (partial
= 0.073): 14 out of 19 and 13 out of 19 participants demonstrated greater EMG amplitude during 40% and 70% MVIC contraction intensities during position tasks, respectively, when compared to the force tasks. In addition, there were differences in MU firing properties (the relationship between RT vs. MFR and RT vs. DT) between the two tasks despite no significant difference in EMG amplitude.
Our results of MU firing properties show an interesting finding: the slope coefficients of the MU RT vs. MFR and RT vs. DT relationships were more negative and more positive, respectively, during the position task when compared to these during the force task (both intensities merged). According to De Luca and Hostage [
17], the linear relationship between the MU RT and MFR describes an “operating point” of the motoneuron pool responding to different levels of excitation. With a greater level of contraction intensity (force output), the slope and y-intercept of this relationship become less negative and greater, respectively, suggesting the recruitment of the high-threshold MUs, as well as the increased firing rates of the active MUs. The current finding showed a more negative slope coefficient for the MU RT vs. MFR relationship under the position task, as compares to the force task (contraction intensities merged). This, therefore, confirms the different MU control strategies used between these tasks. It seems that compared to the force task contraction, the recruitment of the relatively high-threshold MUs might not be the main strategy to reach the same force output when performing the position task contraction. For the relationship between the MU RT and DT, it has been shown to be linear and positive in previous studies. [
17,
18]. Generally speaking, low-threshold and high-threshold MUs are usually derecruited at lower force and at higher force levels, respectively, than which they were recruited (De Luca and Hostage 2010; De Luca et al. 1982; Farina et al. 2009; Stock and Mota 2017), indicating that with the same absolute high force output, fewer active MUs are needed during derecruitment than during recruitment. This, however, may not always be true, because different muscles can add variation to this relationship [
17,
28]. In the current study, the more positive slope coefficient of the MU RT vs. DT relationship was found during the position task than that during the force task. This suggests that based on the theoretical slope regression line shown in the results, active MUs during the position task derecruited at higher force levels than where they were recruited. It also indicates that, when compared to the same absolute force levels between two isometric tasks, fewer MUs were required during derecruitment (force reducing phase) in the position task than that in the force task.
Our results of the y-intercepts of MU RT vs. MFR and RT vs. DT relationships were greater and lower at 70% MVIC than at 40% MVIC, respectively (both tasks merged). The increase of the force output from 40% to 70% MVIC is largely due to the greater neural drive to the motoneuron pools of the active muscles, inducing the recruitment of additional high-threshold MU, the increased firing rates of the active MUs, and the declined derecruitment threshold [
12,
17]. Thus, considering the y-intercepts of the MU RT vs. MFR and RT vs. DT relationships indicate the theoretical maximal firing rates and derecruitment of the active MU [
17], the results of the y-intercepts were within our expectation and in agreement with previous studies [
12,
17].
The current study found the task-related differences in the slopes of the linear relationships between the MU RT and MFR, and between the RT and DT during the non-fatiguing contractions. However, it is unclear what mechanism(s) could have contributed to these differences. There are several worth-mentioning limitations for this experiment. First, although the reflex-related mechanism may support the current findings, it is still speculative to link the current MU control results to this mechanism without any caution, because we were not able to examine the reflex responses for both tasks. Second, methodological-related limitations should also be identified to improve similar future experiments. For example, EMG was not normalized to the maximal compound muscle action potential or collected from synergist muscles. In addition, the execution of the submaximal trapezoid contraction during the position task could also be influenced by the force production from the research staff.
5. Conclusions
In conclusion, even though there were no differences in agonist and antagonist EMG amplitude responses between two different isometric tasks, the present study found task-related differences in MU control strategies between the force and position tasks. The recruitment of high-threshold MUs and the number of active MUs at derecruitment could have contributed to the task-related difference between two tasks. To reach and maintain the same force output, the position task may rely less on the recruitment of relatively high-threshold MUs, as compared to the force task. In addition, as the force output decreases, MUs tend to derecruit at higher force levels during the position task than during the force task. However, the exact mechanisms for the different MU control strategies between two tasks remain unclear. To further understand the underlying mechanism(s) for MU control strategies for the two isometric tasks, it is necessary to examine the changes of excitatory and inhibitory input that occur in the spinal or supraspinal level. The changes in heteronymous H-reflex can estimate modulation monosynaptic input to the spinal cord and is likely mediated by presynaptic inhibition of Ia afferent to the muscle. Thus, the potential difference in reflex responses, as well as the difference in MU activities between isometric tasks could be explored in future research.
Isometric muscle performance is a common measurement to evaluate neuromuscular functions and fitness levels in the field of sports science [
29]. In addition, isometric muscle action is an important means for patients during the rehabilitation process. Despite similar mechanical requirements for both isometric tasks, the current results suggest that force and position tasks seem to utilize different MU control strategies. This task-related difference can provide important information when designing experiments in the field of sports science, or designing exercise programs for patients in rehabilitation settings. Implementing different isometric tasks during the rehabilitation may yield distinct benefits for certain clinical populations.








{kind=link}
{kind=link}
{kind=link}
{kind=link}