
Alterations in Circulating MicroRNAs and the Relation of MicroRNAs to Maximal Oxygen Consumption and Intima–Media Thickness in Ultra-Marathon Runners

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. Blood Collection
2.3. RNA Preparation, Detection, and Quantification of miRNAs by Quantitative PCR
2.4. Statistical Analysis
3. Results
3.1. Participants
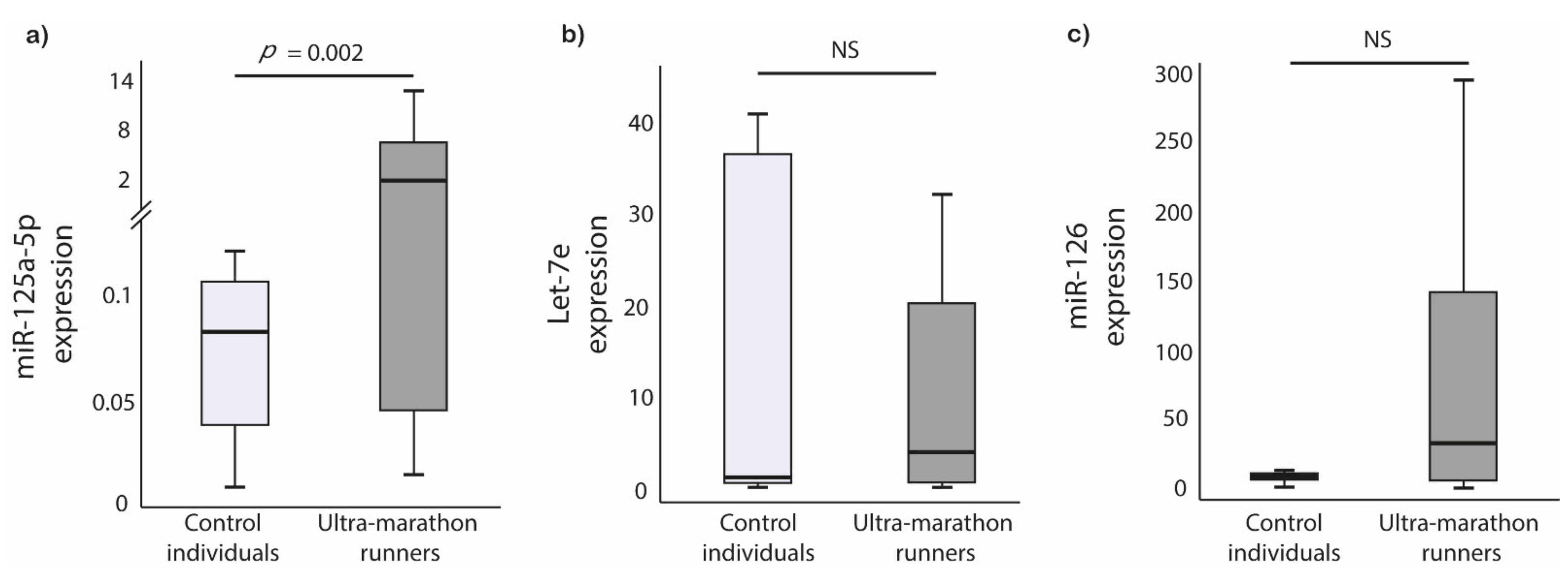
3.2. Circulating miRNAs
3.3. Correlations between miRNAs and Clinical Parameters
3.4. Carotid Intima–Media Thickness and miRNAs Expression
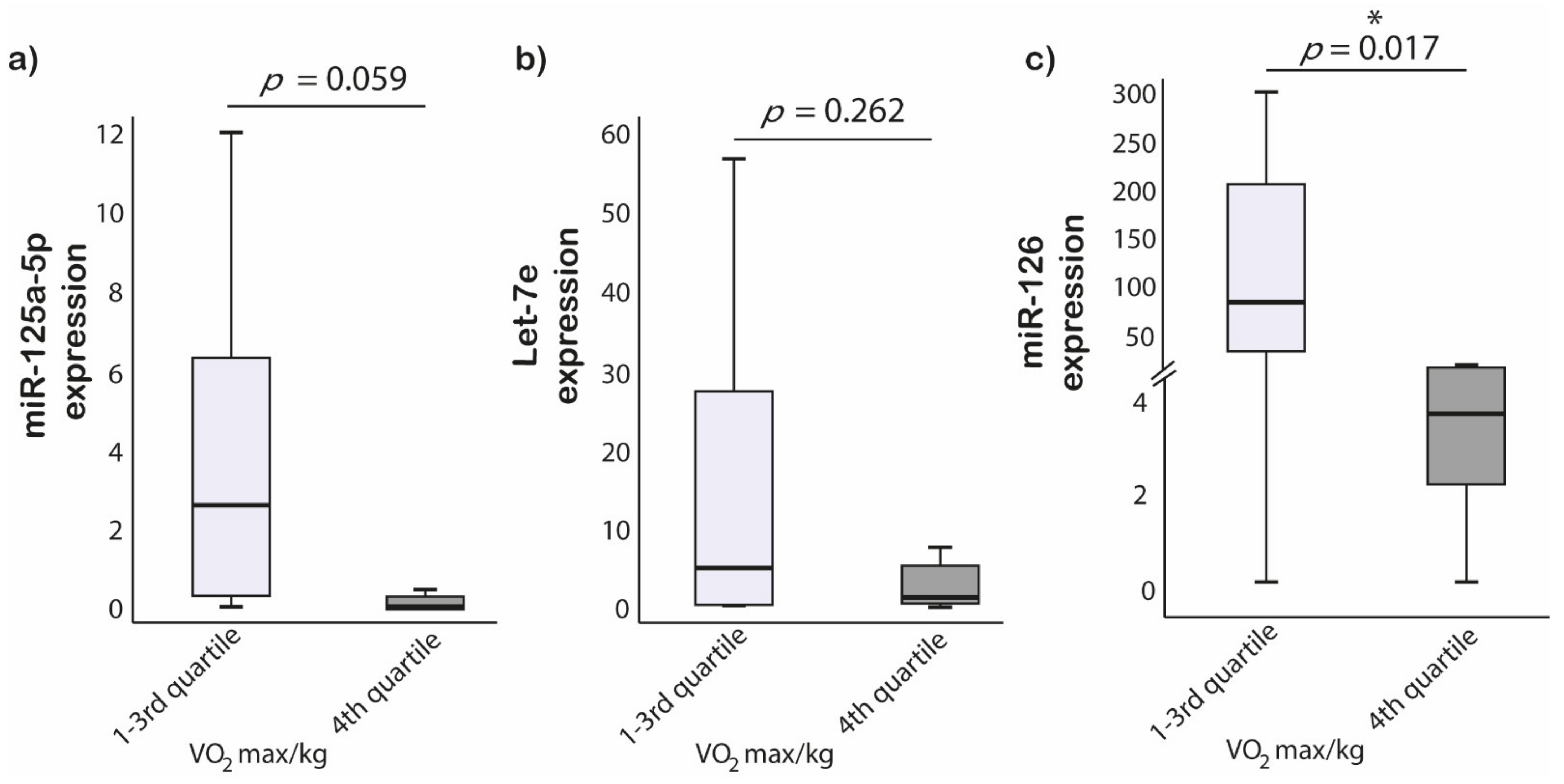
3.5. Cardiopulmonary Fitness and miRNAs Expression
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Helgerud, J.; Høydal, K.; Wang, E.; Karlsen, T.; Berg, P.; Bjerkaas, M.; Simonsen, T.; Helgesen, C.; Hjorth, N.; Bach, R.; et al. Aerobic High-Intensity Intervals Improve VO2max More than Moderate Training. Med. Sci. Sports Exerc. 2007, 39, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Seiler, S.; Tønnessen, E. Intervals, Thresholds and Long Slow Distance: The Role of Intensity and Duration in Endurance Training. Sportscience 2009, 13, 32–53. [Google Scholar]
- Pelliccia, A.; Caselli, S. Structural and Functional Adaptations in the Athlete’s Heart. In The ESC Textbook of Sports Cardiology; Section 1.2.1; Oxford University Press: Oxford, UK, 2019; pp. 9–20. [Google Scholar] [CrossRef]
- MacInnis, M.J.; Gibala, M.J. Physiological Adaptations to Interval Training and the Role of Exercise Intensity. J. Physiol. 2017, 595, 2915–2930. [Google Scholar] [CrossRef] [Green Version]
- Małek, Ł.A.; Barczuk-Falęcka, M.; Werys, K.; Czajkowska, A.; Mróz, A.; Witek, K.; Burrage, M.; Bakalarski, W.; Nowicki, D.; Roik, D.; et al. Cardiovascular Magnetic Resonance with Parametric Mapping in Long-Term Ultra-Marathon Runners. Eur. J. Radiol. 2019, 117, 89–94. [Google Scholar] [CrossRef]
- Konwerski, M.; Postuła, M.; Barczuk-Falęcka, M.; Czajkowska, A.; Mróz, A.; Witek, K.; Bakalarski, W.; Gąsecka, A.; Małek, Ł.A.; Mazurek, T. Epicardial Adipose Tissue and Cardiovascular Risk Assessment in Ultra-Marathon Runners: A Pilot Study. Int. J. Environ. Res. Public Health 2021, 18, 3136. [Google Scholar] [CrossRef] [PubMed]
- Hellsten, Y.; Nyberg, M. Cardiovascular Adaptations to Exercise Training. Compr. Physiol. 2015, 6, 1–32. [Google Scholar] [CrossRef]
- Williams, P.T. Physical Fitness and Activity as Separate Heart Disease Risk Factors: A Meta-Analysis. Med. Sci. Sports Exerc. 2001, 33, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Clanton, T.L. Hypoxia-Induced Reactive Oxygen Species Formation in Skeletal Muscle. J. Appl. Physiol. 2007, 102, 2379–2388. [Google Scholar] [CrossRef] [Green Version]
- Möhlenkamp, S.; Lehmann, N.; Breuckmann, F.; Bröcker-Preuss, M.; Nassenstein, K.; Halle, M.; Budde, T.; Mann, K.; Barkhausen, J.; Heusch, G.; et al. Running: The Risk of Coronary Events: Prevalence and Prognostic Relevance of Coronary Atherosclerosis in Marathon Runners. Eur. Heart J. 2008, 29, 1903–1910. [Google Scholar] [CrossRef] [Green Version]
- Soplinska, A.; Zareba, L.; Wicik, Z.; Eyileten, C.; Jakubik, D.; Siller-Matula, J.M.; De Rosa, S.; Malek, L.A.; Postula, M. MicroRNAs as Biomarkers of Systemic Changes in Response to Endurance Exercise—A Comprehensive Review. Diagnostics 2020, 10, 813. [Google Scholar] [CrossRef]
- Aengevaeren, V.L.; Mosterd, A.; Braber, T.L.; Prakken, N.H.J.; Doevendans, P.A.; Grobbee, D.E.; Thompson, P.D.; Eijsvogels, T.M.H.; Velthuis, B.K. Relationship Between Lifelong Exercise Volume and Coronary Atherosclerosis in Athletes. Circulation 2017, 136, 138–148. [Google Scholar] [CrossRef]
- Merghani, A.; Maestrini, V.; Rosmini, S.; Cox, A.T.; Dhutia, H.; Bastiaenan, R.; David, S.; Yeo, T.J.; Narain, R.; Malhotra, A.; et al. Prevalence of Subclinical Coronary Artery Disease in Masters Endurance Athletes With a Low Atherosclerotic Risk Profile. Circulation 2017, 136, 126–137. [Google Scholar] [CrossRef]
- Davis, P.H.; Dawson, J.D.; Riley, W.A.; Lauer, R.M. Carotid Intimal-Medial Thickness Is Related to Cardiovascular Risk Factors Measured from Childhood through Middle Age: The Muscatine Study. Circulation 2001, 104, 2815–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scuteri, A.; Franco, O.H.; Majiid, A.; Jolita, B.; Sergey, B.; Cheng, H.-M.; Chen, C.-H.; Choi, S.-W.; Francesco, C.; De Buyzere, M.L.; et al. The Relationship between the Metabolic Syndrome and Arterial Wall Thickness: A Mosaic Still to Be Interpreted. Atherosclerosis 2016, 255, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, S.; Papacosta, O.; Whincup, P.; Wannamethee, G.; Walker, M.; Nicolaides, A.N.; Dhanjil, S.; Griffin, M.; Belcaro, G.; Rumley, A.; et al. Carotid Plaque, Intima Media Thickness, Cardiovascular Risk Factors, and Prevalent Cardiovascular Disease in Men and Women: The British Regional Heart Study. Stroke 1999, 30, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, S.H.; Mathiesen, E.B. Carotid Plaque Compared with Intima-Media Thickness as a Predictor of Coronary and Cerebrovascular Disease. Curr. Cardiol. Rep. 2009, 11, 21–27. [Google Scholar] [CrossRef]
- Tatsukawa, M.; Sawayama, Y.; Maeda, N.; Okada, K.; Furusyo, N.; Kashiwagi, S.; Hayashi, J. Carotid Atherosclerosis and Cardiovascular Risk Factors: A Comparison of Residents of a Rural Area of Okinawa with Residents of a Typical Suburban Area of Fukuoka, Japan. Atherosclerosis 2004, 172, 337–343. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in Cardiovascular Biology and Disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Jakubik, D.; Fitas, A.; Eyileten, C.; Jarosz-Popek, J.; Nowak, A.; Czajka, P.; Wicik, Z.; Sourij, H.; Siller-Matula, J.M.; De Rosa, S.; et al. MicroRNAs and Long Non-Coding RNAs in the Pathophysiological Processes of Diabetic Cardiomyopathy: Emerging Biomarkers and Potential Therapeutics. Cardiovasc. Diabetol. 2021, 20, 55. [Google Scholar] [CrossRef]
- Zareba, L.; Fitas, A.; Wolska, M.; Junger, E.; Eyileten, C.; Wicik, Z.; De Rosa, S.; Siller-Matula, J.M.; Postula, M. MicroRNAs and Long Noncoding RNAs in Coronary Artery Disease: New and Potential Therapeutic Targets. Cardiol. Clin. 2020, 38, 601–617. [Google Scholar] [CrossRef]
- Uhlemann, M.; Möbius-Winkler, S.; Fikenzer, S.; Adam, J.; Redlich, M.; Möhlenkamp, S.; Hilberg, T.; Schuler, G.C.; Adams, V. Circulating microRNA-126 Increases after Different Forms of Endurance Exercise in Healthy Adults. Eur. J. Prev. Cardiol. 2014, 21, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Åkerström, T.; Rinnov, A.; Yfanti, C.; Scheele, C.; Pedersen, B.K.; Laye, M.J. The miRNA Plasma Signature in Response to Acute Aerobic Exercise and Endurance Training. PLoS ONE 2014, 9, e87308. [Google Scholar] [CrossRef]
- Baggish, A.L.; Hale, A.; Weiner, R.B.; Lewis, G.D.; Systrom, D.; Wang, F.; Wang, T.J.; Chan, S.Y. Dynamic Regulation of Circulating microRNA during Acute Exhaustive Exercise and Sustained Aerobic Exercise Training. J. Physiol. 2011, 589, 3983–3994. [Google Scholar] [CrossRef]
- Clauss, S.; Wakili, R.; Hildebrand, B.; Kääb, S.; Hoster, E.; Klier, I.; Martens, E.; Hanley, A.; Hanssen, H.; Halle, M.; et al. MicroRNAs as Biomarkers for Acute Atrial Remodeling in Marathon Runners (The miRathon Study—A Sub-Study of the Munich Marathon Study). PLoS ONE 2016, 11, e0148599. [Google Scholar] [CrossRef]
- Nielsen, S.; Scheele, C.; Yfanti, C.; Akerström, T.; Nielsen, A.R.; Pedersen, B.K.; Laye, M.J. Muscle Specific microRNAs Are Regulated by Endurance Exercise in Human Skeletal Muscle. J. Physiol. 2010, 588, 4029–4037. [Google Scholar] [CrossRef]
- Polakovičová, M.; Musil, P.; Laczo, E.; Hamar, D.; Kyselovič, J. Circulating MicroRNAs as Potential Biomarkers of Exercise Response. Int. J. Mol. Sci. 2016, 17, 1553. [Google Scholar] [CrossRef] [PubMed]
- Pordzik, J.; Pisarz, K.; De Rosa, S.; Jones, A.D.; Eyileten, C.; Indolfi, C.; Malek, L.; Postula, M. The Potential Role of Platelet-Related microRNAs in the Development of Cardiovascular Events in High-Risk Populations, Including Diabetic Patients: A Review. Front. Endocrinol. 2018, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The Endothelial-Specific microRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Zernecke, A.; Bidzhekov, K.; Noels, H.; Shagdarsuren, E.; Gan, L.; Denecke, B.; Hristov, M.; Köppel, T.; Jahantigh, M.N.; Lutgens, E.; et al. Delivery of microRNA-126 by Apoptotic Bodies Induces CXCL12-Dependent Vascular Protection. Sci. Signal. 2009, 2, ra81. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA Expression in Circulating Microvesicles Predicts Cardiovascular Events in Patients with Coronary Artery Disease. J. Am. Heart Assoc. 2014, 3, e001249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Shao, S.; Geng, H.; Yu, Y.; Wang, C.; Liu, Z.; Yu, C.; Jiang, X.; Deng, Y.; Gao, L.; et al. Expression Profiles of Six Circulating microRNAs Critical to Atherosclerosis in Patients with Subclinical Hypothyroidism: A Clinical Study. J. Clin. Endocrinol. Metab. 2014, 99, E766–E774. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Wang, X.-G.; Cheng, J.-D.; You, S.-Z.; Ma, S.-H.; Zhong, X.; Quan, L.; Luo, B. The up-Regulation of Endothelin-1 and down-Regulation of miRNA-125a-5p, -155, and -199a/b-3p in Human Atherosclerotic Coronary Artery. Cardiovasc. Pathol. 2014, 23, 217–223. [Google Scholar] [CrossRef]
- De Gonzalo-Calvo, D.; Dávalos, A.; Montero, A.; García-González, Á.; Tyshkovska, I.; González-Medina, A.; Soares, S.M.A.; Martínez-Camblor, P.; Casas-Agustench, P.; Rabadán, M.; et al. Circulating Inflammatory miRNA Signature in Response to Different Doses of Aerobic Exercise. J. Appl. Physiol. 2015, 119, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Backes, C.; Leidinger, P.; Keller, A.; Hart, M.; Meyer, T.; Meese, E.; Hecksteden, A. Blood Born miRNAs Signatures That Can Serve as Disease Specific Biomarkers Are Not Significantly Affected by Overall Fitness and Exercise. PLoS ONE 2014, 9, e102183. [Google Scholar] [CrossRef]
- Małek, Ł.A.; Mazurkiewicz, Ł.; Marszałek, M.; Barczuk-Falęcka, M.; Simon, J.E.; Grzybowski, J.; Miłosz-Wieczorek, B.; Postuła, M.; Marczak, M. Deformation Parameters of the Heart in Endurance Athletes and in Patients with Dilated Cardiomyopathy-A Cardiac Magnetic Resonance Study. Diagnostics 2021, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Malhotra, R.; Chiampas, G.; d’Hemecourt, P.; Troyanos, C.; Cianca, J.; Smith, R.N.; Wang, T.J.; Roberts, W.O.; Thompson, P.D.; et al. Cardiac Arrest during Long-Distance Running Races. N. Engl. J. Med. 2012, 366, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Knechtle, B.; Nikolaidis, P.T. Physiology and Pathophysiology in Ultra-Marathon Running. Front. Physiol. 2018, 9, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerche, A.L.; La Gerche, A.; Burns, A.T.; Mooney, D.J.; Inder, W.J.; Taylor, A.J.; Bogaert, J.; MacIsaac, A.I.; Heidbüchel, H.; Prior, D.L. Exercise-Induced Right Ventricular Dysfunction and Structural Remodelling in Endurance Athletes. Eur. Heart J. 2012, 33, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Małek, Ł.A.; Czajkowska, A.; Mróz, A.; Witek, K.; Nowicki, D.; Postuła, M. Factors Related to Cardiac Troponin T Increase after Participation in a 100 Km Ultra-Marathon. Diagnostics 2020, 10, 167. [Google Scholar] [CrossRef] [Green Version]
- Spanidis, Y.; Stagos, D.; Orfanou, M.; Goutzourelas, N.; Bar-Or, D.; Spandidos, D.; Kouretas, D. Variations in Oxidative Stress Levels in 3 Days Follow-up in Ultramarathon Mountain Race Athletes. J. Strength Cond. Res. 2017, 31, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Mrakic-Sposta, S.; Gussoni, M.; Moretti, S.; Pratali, L.; Giardini, G.; Tacchini, P.; Dellanoce, C.; Tonacci, A.; Mastorci, F.; Borghini, A.; et al. Effects of Mountain Ultra-Marathon Running on ROS Production and Oxidative Damage by Micro-Invasive Analytic Techniques. PLoS ONE 2015, 10, e0141780. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.P.C.; Oliveira, G.P., Jr.; Madrid, B.; Almeida, J.A.; Franco, O.L.; Pereira, R.W. Circulating miR-1, miR-133a, and miR-206 Levels Are Increased after a Half-Marathon Run. Biomarkers 2014, 19, 585–589. [Google Scholar] [CrossRef]
- Harriss, D.J.; MacSween, A.; Atkinson, G. Ethical Standards in Sport and Exercise Science Research: 2020 Update. Int. J. Sports Med. 2019, 40, 813–817. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, S.; Eposito, F.; Carella, C.; Strangio, A.; Ammirati, G.; Sabatino, J.; Abbate, F.G.; Iaconetti, C.; Liguori, V.; Pergola, V.; et al. Transcoronary Concentration Gradients of Circulating microRNAs in Heart Failure. Eur. J. Heart Fail. 2018, 20, 1000–1010. [Google Scholar] [CrossRef]
- De Rosa, R.; De Rosa, S.; Leistner, D.; Boeckel, J.-N.; Keller, T.; Fichtlscherer, S.; Dimmeler, S.; Zeiher, A.M. Transcoronary Concentration Gradient of microRNA-133a and Outcome in Patients With Coronary Artery Disease. Am. J. Cardiol. 2017, 120, 15–24. [Google Scholar] [CrossRef]
- Gareri, C.; Iaconetti, C.; Sorrentino, S.; Covello, C.; De Rosa, S.; Indolfi, C. miR-125a-5p Modulates Phenotypic Switch of Vascular Smooth Muscle Cells by Targeting ETS-1. J. Mol. Biol. 2017, 429, 1817–1828. [Google Scholar] [CrossRef]
- Schmitz, B.; Breulmann, F.L.; Jubran, B.; Rolfes, F.; Thorwesten, L.; Krüger, M.; Klose, A.; Schnittler, H.-J.; Brand, S.-M. A Three-Step Approach Identifies Novel Shear Stress-Sensitive Endothelial microRNAs Involved in Vasculoprotective Effects of High-Intensity Interval Training (HIIT). Oncotarget 2019, 10, 3625–3640. [Google Scholar] [CrossRef] [PubMed]
- Bye, A.; Røsjø, H.; Aspenes, S.T.; Condorelli, G.; Omland, T.; Wisløff, U. Circulating MicroRNAs and Aerobic Fitness—The HUNT-Study. PLoS ONE 2013, 8, e57496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, K.S.; Gotlieb, A.I. The Role of Shear Stress in the Pathogenesis of Atherosclerosis. Lab. Investig. 2005, 85, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, P.; Wehmeier, U.F.; Jansen, F.J.; Kilian, Y.; Bloch, W.; Werner, N.; Mester, J.; Hilberg, T. Acute Effects of Different Exercise Protocols on the Circulating Vascular microRNAs -16, -21, and -126 in Trained Subjects. Front. Physiol. 2016, 7, 643. [Google Scholar] [CrossRef] [Green Version]
- Tang, F.; Yang, T.-L. MicroRNA-126 Alleviates Endothelial Cells Injury in Atherosclerosis by Restoring Autophagic Flux via Inhibiting of PI3K/Akt/mTOR Pathway. Biochem. Biophys. Res. Commun. 2018, 495, 1482–1489. [Google Scholar] [CrossRef]
- Denham, J.; Prestes, P.R. Muscle-Enriched MicroRNAs Isolated from Whole Blood Are Regulated by Exercise and Are Potential Biomarkers of Cardiorespiratory Fitness. Front. Genet. 2016, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- Truettner, J.S.; Katyshev, V.; Esen-Bilgin, N.; Dietrich, W.D.; Dore-Duffy, P. Hypoxia Alters MicroRNA Expression in Rat Cortical Pericytes. Microrna 2013, 2, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, B.; Schelleckes, K.; Nedele, J.; Thorwesten, L.; Klose, A.; Lenders, M.; Krüger, M.; Brand, E.; Brand, S.-M. Dose-Response of High-Intensity Training (HIT) on Atheroprotective miRNA-126 Levels. Front. Physiol. 2017, 8, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thum, T. Noncoding RNAs and Myocardial Fibrosis. Nat. Rev. Cardiol. 2014, 11, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Liu, J.; He, F.; Xu, W.; Yao, K. Hypoxia-Induced Deregulation of miR-126 and Its Regulative Effect on VEGF and MMP-9 Expression. Int. J. Med. Sci. 2014, 11, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempsey, J.A.; Wagner, P.D. Exercise-Induced Arterial Hypoxemia. J. Appl. Physiol. 1999, 87, 1997–2006. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 Modulates Endothelial Cell Response to Hypoxia and Inhibits the Receptor Tyrosine Kinase Ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-J.; Ko, K.-J. Association between Resting Heart Rate, VOmax and Carotid Intima-Media Thickness in Middle-Aged Men. Int. J. Cardiol. Heart Vasc. 2019, 23, 100347. [Google Scholar] [CrossRef]
- Hao, X.-Z.; Fan, H.-M. Identification of miRNAs as Atherosclerosis Biomarkers and Functional Role of miR-126 in Atherosclerosis Progression through MAPK Signalling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2725–2733. [Google Scholar]
- Van Thienen, J.V.; Fledderus, J.O.; Dekker, R.J.; Rohlena, J.; van Ijzendoorn, G.A.; Kootstra, N.A.; Pannekoek, H.; Horrevoets, A.J.G. Shear Stress Sustains Atheroprotective Endothelial KLF2 Expression More Potently than Statins through mRNA Stabilization. Cardiovasc. Res. 2006, 72, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 Regulates Endothelial Expression of Vascular Cell Adhesion Molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Huang, Z.; Wang, L.; Wang, Y.; Wu, F.; Meng, S.; Wang, C. MicroRNA-125a-5p Partly Regulates the Inflammatory Response, Lipid Uptake, and ORP9 Expression in oxLDL-Stimulated Monocyte/macrophages. Cardiovasc. Res. 2009, 83, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, Q.; Yu, J.; Cao, X.; Xu, Z. miR-125a-5p Inhibits the Expression of NLRP3 by Targeting CCL4 in Human Vascular Smooth Muscle Cells Treated with Ox-LDL. Exp. Ther. Med. 2019, 18, 1645–1652. [Google Scholar] [CrossRef]
- Pan, Q.; Liao, X.; Liu, H.; Wang, Y.; Chen, Y.; Zhao, B.; Lazartigues, E.; Yang, Y.; Ma, X. MicroRNA-125a-5p Alleviates the Deleterious Effects of Ox-LDL on Multiple Functions of Human Brain Microvessel Endothelial Cells. Am. J. Physiol. Cell Physiol. 2017, 312, C119–C130. [Google Scholar] [CrossRef]
- Pan, Q.; Ma, C.; Wang, Y.; Wang, J.; Zheng, J.; Du, D.; Liao, X.; Chen, Y.; Chen, Y.; Bihl, J.; et al. Microvesicles-Mediated Communication between Endothelial Cells Modulates, Endothelial Survival, and Angiogenic Function via Transferring of miR-125a-5p. J. Cell. Biochem. 2019, 120, 3160–3172. [Google Scholar] [CrossRef]
- Khalyfa, A.; Kheirandish-Gozal, L.; Bhattacharjee, R.; Khalyfa, A.A.; Gozal, D. Circulating microRNAs as Potential Biomarkers of Endothelial Dysfunction in Obese Children. Chest 2016, 149, 786–800. [Google Scholar] [CrossRef] [Green Version]
- De Gonzalo-Calvo, D.; Dávalos, A.; Fernández-Sanjurjo, M.; Amado-Rodríguez, L.; Díaz-Coto, S.; Tomás-Zapico, C.; Montero, A.; García-González, Á.; Llorente-Cortés, V.; Heras, M.E.; et al. Circulating microRNAs as Emerging Cardiac Biomarkers Responsive to Acute Exercise. Int. J. Cardiol. 2018, 264, 130–136. [Google Scholar] [CrossRef]
- DeFina, L.F.; Radford, N.B.; Barlow, C.E.; Willis, B.L.; Leonard, D.; Haskell, W.L.; Farrell, S.W.; Pavlovic, A.; Abel, K.; Berry, J.D.; et al. Association of All-Cause and Cardiovascular Mortality With High Levels of Physical Activity and Concurrent Coronary Artery Calcification. JAMA Cardiol. 2019, 4, 174–181. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Ultra-Marathon Runners n = 30, Median (IQR) |
|---|---|
| Years of running (running history) | 9 (7–15) |
| Years of ultra-training | 6 (5–8) |
| Age at start of ultra-running | 34 (29–39) |
| Total covered distance (km) | 25,000 (20,000–40,000) |
| Weekly running distance (km) | 80 (70–90) |
| Number of ultra-races completed | 15 (10–27.5) |
| Number of ultra-races during previous year | 5.5 (4–9) |
| Number of completed ultra-races >100 km | 3.5 (2–7) |
| Best place achieved in an ultra-race | 5 (1–13) |
| Longest run (km) | 150 (106–246) |
| Most frequent ultra-race distance (km) | 100 (70–100) |
| Parameter | Ultra-Marathon Runners (n = 30) | Control Group (n = 9) | p Value |
|---|---|---|---|
| Age (years) | 40.93 ± 6.57 | 40.0 ± 8.32 | 0.76 |
| Height (cm) | 178 ± 5 | 179 ± 4 | 0.51 |
| Weight (kg) | 71.9 ± 4.7 | 83 ± 6.1 | 0.12 |
| BMI (cm/m2) | 22.07 ± 1.54 | 26.1 ± 1.5 | 0.13 |
| Systolic BP (mmHg) | 128 ± 6 | 126 ± 7 | 0.78 |
| Diastolic BP (mmHg) | 78 ± 6 | 80 ± 5 | 0.35 |
| Hct (%/100) | 0.43 ± 0.03 | 0.45 ± 0.02 | 0.01 * |
| Resting HR (bpm) | 54.9 ± 9.2 | 69.6 ± 11.0 | 0.005 * |
| VO2 max (L/min) | 4.43 ± 0.44 | 3.63 ± 0.63 | 0.005 * |
| VO2 max (mL/min/kg) | 61.0 ± 4.9 | 40.2 ± 4.6 | <0.001 * |
| VE max (L) | 150.4 ± 16.9 | 127.9 ± 25.3 | 0.03* |
| RER max | 1.08 ± 0.05 | 1.10 ± 0.04 | 0.17 |
| LVEDVI (mL/m2) | 110 ± 15 | 78 ± 10 | <0.0001 * |
| LVESVI (mL/m2) | 39 ± 9 | 27 ± 4 | <0.0001 * |
| LVSVI (mL/m2) | 71 ± 9 | 51 ± 9 | <0.0001 * |
| LVEF (%) | 65 ± 5 | 66 ± 5 | 0.59 |
| LVMI (g/m2) | 83 ± 11 | 66 ± 11 | 0.001 * |
| RVEDVI (mL/m2) | 125 ± 20 | 88 ± 12 | <0.0001 * |
| RVESVI (mL/m2) | 52 ± 11 | 37 ± 6 | <0.0001 * |
| RVEF (%) | 59 ± 4 | 58 ± 5 | 0.61 |
| RVMI (g/m2) | 24 ± 3.5 | 22 ± 5 | 0.24 |
| IVSD (mm) | 11.5 ± 2 | 10.5 ± 1 | 0.32 |
| PWD (mm) | 10 ± 1 | 9 ± 1 | 0.17 |
| Left CIMT | 0.07 ± 0.02 | 0.07 ± 0.01 | 0.99 |
| Right CIMT | 0.08 ± 0.03 | 0.08 ± 0.01 | 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eyileten, C.; Fitas, A.; Jakubik, D.; Czajka, P.; Mróz, A.; Czajkowska, A.; Witek, K.; Bakalarski, W.; De Rosa, S.; Postuła, M.; et al. Alterations in Circulating MicroRNAs and the Relation of MicroRNAs to Maximal Oxygen Consumption and Intima–Media Thickness in Ultra-Marathon Runners. Int. J. Environ. Res. Public Health 2021, 18, 7234. https://doi.org/10.3390/ijerph18147234
Eyileten C, Fitas A, Jakubik D, Czajka P, Mróz A, Czajkowska A, Witek K, Bakalarski W, De Rosa S, Postuła M, et al. Alterations in Circulating MicroRNAs and the Relation of MicroRNAs to Maximal Oxygen Consumption and Intima–Media Thickness in Ultra-Marathon Runners. International Journal of Environmental Research and Public Health. 2021; 18(14):7234. https://doi.org/10.3390/ijerph18147234
Chicago/Turabian StyleEyileten, Ceren, Alex Fitas, Daniel Jakubik, Pamela Czajka, Anna Mróz, Anna Czajkowska, Katarzyna Witek, Wawrzyniec Bakalarski, Salvatore De Rosa, Marek Postuła, and et al. 2021. "Alterations in Circulating MicroRNAs and the Relation of MicroRNAs to Maximal Oxygen Consumption and Intima–Media Thickness in Ultra-Marathon Runners" International Journal of Environmental Research and Public Health 18, no. 14: 7234. https://doi.org/10.3390/ijerph18147234
APA StyleEyileten, C., Fitas, A., Jakubik, D., Czajka, P., Mróz, A., Czajkowska, A., Witek, K., Bakalarski, W., De Rosa, S., Postuła, M., & Małek, Ł. A. (2021). Alterations in Circulating MicroRNAs and the Relation of MicroRNAs to Maximal Oxygen Consumption and Intima–Media Thickness in Ultra-Marathon Runners. International Journal of Environmental Research and Public Health, 18(14), 7234. https://doi.org/10.3390/ijerph18147234