
Consistency Is Key When Setting a New World Record for Running 10 Marathons in 10 Days

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Record Attempt
2.2. Measures
2.2.1. Body Composition
2.2.2. Pre-Attempt Laboratory Testing
2.2.3. Cardio-Respiratory, Heart Rate and Subjective Measures
2.2.4. Sleep and Lung Function
2.2.5. Haematology and Body Composition
2.2.6. Nutritional Intake
2.3. Other Recovery Measures
2.4. Statistical Analysis
3. Results
3.1. Laboratory Testing before the Attempt
3.2. Record Breaking Performance
3.3. Body Composition
3.4. Heart Rate
3.5. Sleep and Resting Heart Rate
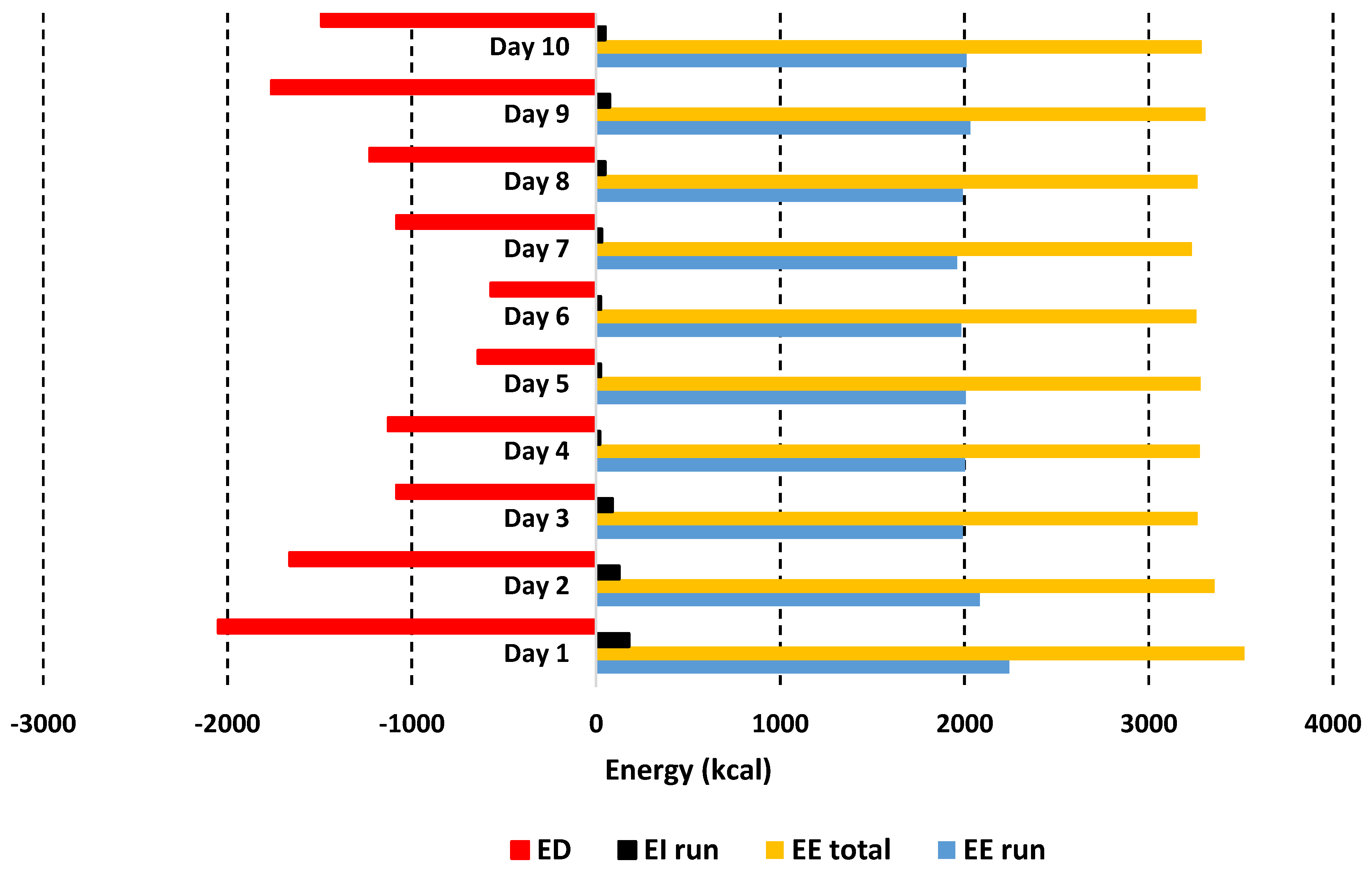
3.6. Lung Function, Oxygen Uptake and Energy Expenditure
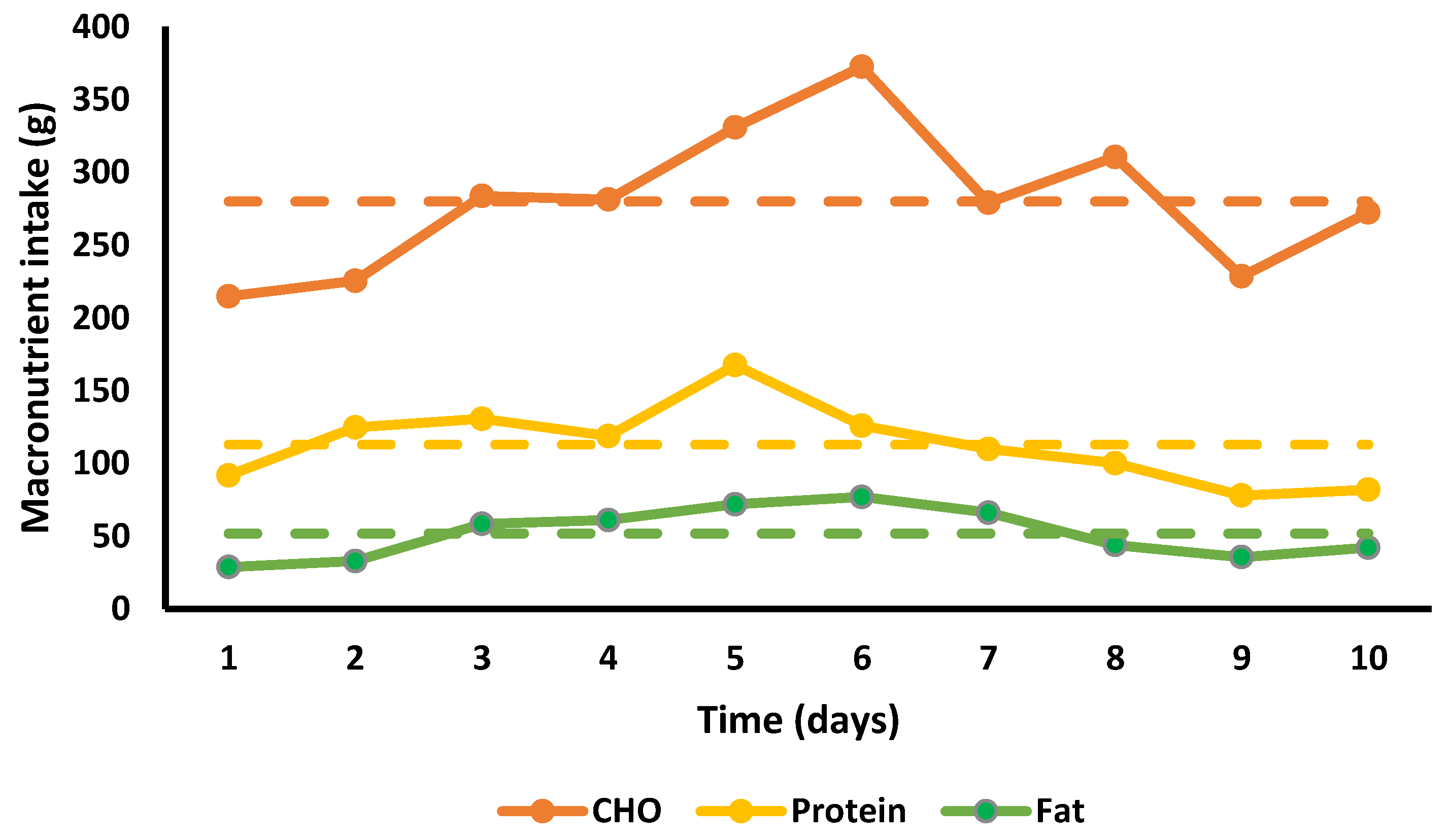
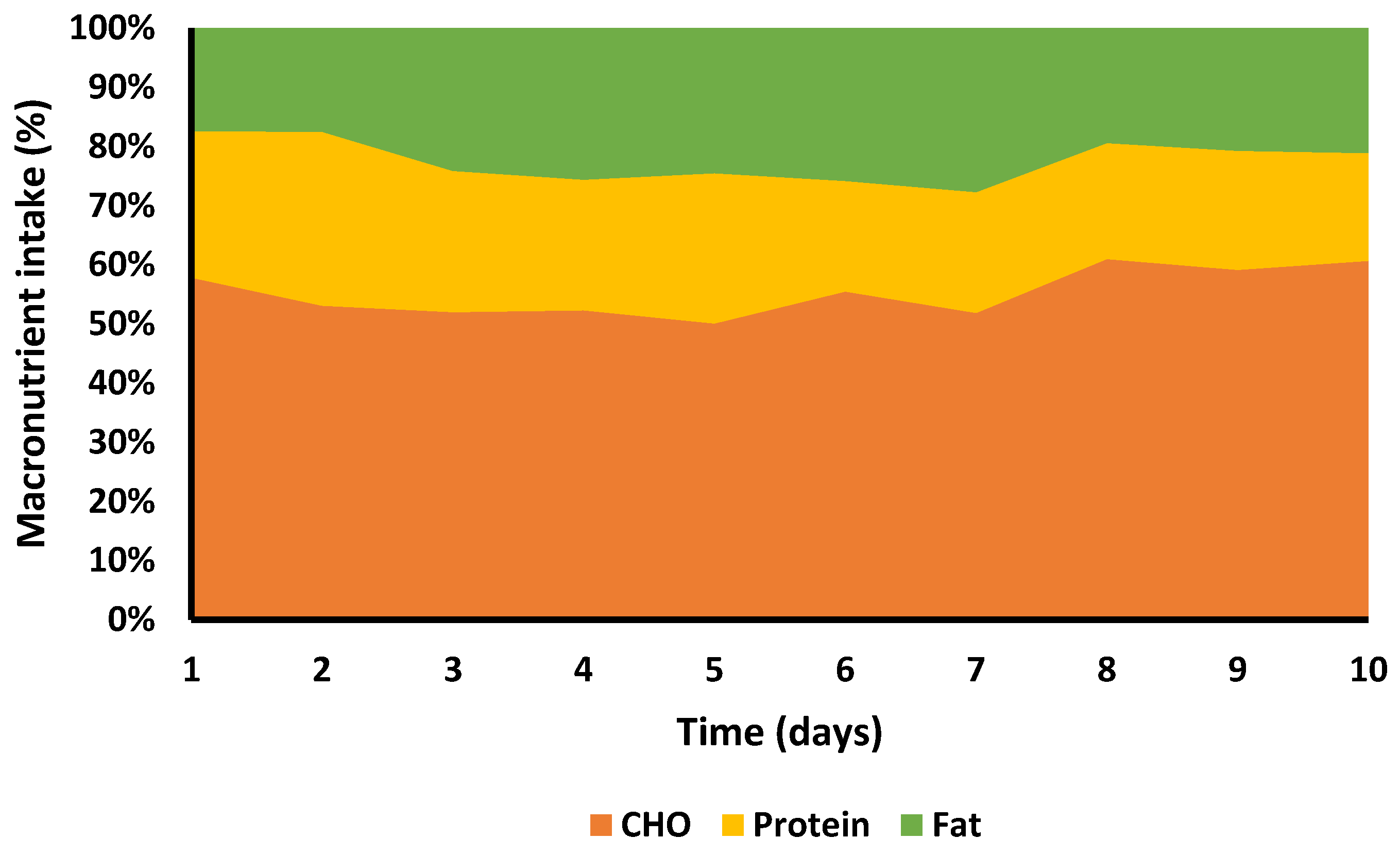
3.7. Nutritional Intake
3.8. Haematology
3.9. Subjective Measures
4. Discussion
4.1. Nutritional Intake and Energy Deficit
4.2. Body Composition Changes
4.3. Respiratory Variables
4.4. Recovery Strategies
4.5. Other Parameters
4.6. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christensen, D.L.; Nielsen, T.H.; Schwartz, A. Herodotos and Hemerodromoi: Pheidippides’ Run from Athens to Sparta in 490 BC from Historical and Physiological Perspectives. Hermes 2009, 137, 148–169. [Google Scholar]
- Matthews, V.J. The “Hemerodromoi”: Ultra Long-Distance Running in Antiquity. Class World 1974, 68, 161. [Google Scholar] [CrossRef]
- Knechtle, B.; Nikolaidis, P.T. Physiology and Pathophysiology in Ultra-Marathon Running. Front. Physiol. 2018, 9, 8–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knechtle, B.; Kohler, G. Running 338 Kilometres within Five Days has no Effect on Body Mass and Body Fat but Reduces Skeletal Muscle Mass—the Isarrun 2006. J. Sports Sci. Med. 2007, 6, 401–407. [Google Scholar]
- Bircher, S.; Enggist, A.; Jehle, T.; Knechtle, B. Effects of an extreme endurance race on energy balance and body composition—A case study. J. Sports Sci. Med. 2006, 5, 154–162. [Google Scholar] [PubMed]
- Machefer, G.; Groussard, C.; Zouhal, H.; Vincent, S.; Youssef, H.; Faure, H.; Malardé, L.; Gratas-Delamarche, A. Nutritional and Plasmatic Antioxidant Vitamins Status of Ultra Endurance Athletes. J. Am. Coll. Nutr. 2007, 26, 311–316. [Google Scholar] [CrossRef]
- Enqvist, J.K.; Mattsson, C.M.; Johansson, P.H.; Brink-Elfegoun, T.; Bakkman, L.; Ekblom, B.T. Energy turnover during 24 h and 6 days of adventure racing. J. Sports Sci. 2010, 28, 947–955. [Google Scholar] [CrossRef]
- Hoogervorst, D.; van der Burg, N.; Versteegen, J.J.; Lambrechtse, K.J.; Redegeld, M.I.; Cornelissen, L.A.J.; Wardenaar, F.C. Gastrointestinal Complaints and Correlations with Self-Reported Macronutrient Intake in Independent Groups of (Ultra)Marathon Runners Competing at Different Distances. Sports 2019, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Knechtle, B.; Knechtle, P.; Rosemann, T.; Senn, O. What is associated with race performance in male 100-km ultra-marathoners—anthropometry, training or marathon best time? J. Sports Sci. 2011, 29, 571–577. [Google Scholar] [CrossRef]
- Schütz, U.H.; Billich, C.; König, K.; Würslin, C.; Wiedelbach, H.; Brambs, H.-J.; Machann, J. Characteristics, changes and influence of body composition during a 4486 km transcontinental ultramarathon: Results from the Transeurope Footrace mobile whole body MRI-project. BMC Med. 2013, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Baur, D.A.; Bach, C.W.; Hyder, W.J.; Ormsbee, M.J. Fluid retention, muscle damage, and altered body composition at the Ultraman triathlon. Eur. J. Appl. Physiol. 2016, 116, 447–458. [Google Scholar] [CrossRef]
- Smith, K.A.; Kisiolek, J.N.; Willingham, B.D.; Morrissey, M.C.; Leyh, S.M.; Saracino, P.G.; Baur, D.A.; Cook, M.D.; Ormsbee, M.J. Ultra-endurance triathlon performance and markers of whole-body and gut-specific inflammation. Eur. J. Appl. Physiol. 2020, 120, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-H.; Lai, J.-I.; Wang, S.-H.; How, C.-K.; Li, L.-H.; Kao, W.-F.; Yang, C.-C.; Chen, R.-J. Early changes of the anemia phenomenon in male 100-km ultramarathoners. J. Chin. Med. Assoc. 2015, 78, 108–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejka, C.; Knechtle, B.; Knechtle, P.; Rüst, C.A.; Rosemann, T. An increased fluid intake leads to feet swelling in 100-km ultra-marathoners—an observational field study. J. Int. Soc. Sport Nutr. 2012, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-H.; Tseng, Y.-F.; Lai, J.-I.; Chen, Y.-Q.; Wang, S.-H.; Kao, W.-F.; Li, L.-H.; Chiu, Y.-H.; How, C.-K.; Chang, W.-H. The changes of red blood cell viscoelasticity and sports anemia in male 24-h ultra-marathoners. J. Chin. Med. Assoc. 2018, 81, 475–481. [Google Scholar] [CrossRef]
- Bossi, A.H.; Matta, G.G.; Millet, G.Y.; Lima, P.; Pertence, L.C.; de Lima, J.P.; Hopker, J.G. Pacing Strategy During 24-h Ultramarathon-Distance Running. Int. J. Sports Physiol. Perform. 2017, 12, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.D.; Valentino, T.R.; Stuempfle, K.J.; Hassid, B.V. A Placebo-Controlled Trial of Riboflavin for Enhancement of Ultramarathon Recovery. Sports Med.-Open 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Kerksick, C.M.; Wilborn, C.D.; Roberts, M.D.; Smith-Ryan, A.; Kleiner, S.M.; Jäger, R.; Collins, R.; Cooke, M.; Davis, J.N.; Galvan, E.; et al. ISSN exercise & sports nutrition review update: Research & recommendations. J. Int. Soc. Sport Nutr. 2018, 15, 38. [Google Scholar] [CrossRef] [Green Version]
- Jayanama, K.; Putadechakun, S.; Srisuwarn, P.; Vallibhakara, S.A.-O.; Shantavasinkul, P.C.; Sritara, C.; Kantachuvesiri, S.; Komindr, S. Evaluation of Body Composition in Hemodialysis Thai Patients: Comparison between Two Models of Bioelectrical Impedance Analyzer and Dual-Energy X-Ray Absorptiometry. J. Nutr. Metab. 2018, 2018, 4537623. [Google Scholar] [CrossRef] [Green Version]
- Winter, E.M.; Jones, A.M.; Davison, R.C.R.; Bromley, P.D.; Mercer, T.H. Sport and Exercise Physiology Testing Guidelines: The British Association of Sport and Exercise Sciences Guide; Taylor & Francis: Abingdon-on-Thames, UK, 2016. [Google Scholar]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Borg, G. Borg’s Perceived Exertion and Pain Scales; Human Kinetics: Champaign, IL, USA, 1998; ISBN 9780880116237. [Google Scholar]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.S.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sport Sci. 2011, 29, S17–S27. [Google Scholar] [CrossRef]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American College of Sports Medicine Joint Position Statement. Nutrition and Athletic Performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [CrossRef]
- Venkatraman, J.T.; Leddy, J.; Pendergast, D. Dietary fats and immune status in athletes: Clinical implications. Med. Sci. Sports Exerc. 2000, 32, S389–S395. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A. A Step Towards Personalized Sports Nutrition: Carbohydrate Intake During Exercise. Sports Med. 2014, 44, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Close, G.L.; Hamilton, D.L.; Philp, A.; Burke, L.M.; Morton, J.P. New strategies in sport nutrition to increase exercise performance. Free. Radic. Biol. Med. 2016, 98, 144–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, N.A.; Koehle, M.S. Optimizing recovery to support multi-evening cycling competition performance. Eur. J. Sport Sci. 2018, 19, 811–823. [Google Scholar] [CrossRef]
- Viribay, A.; Arribalzaga, S.; Mielgo-Ayuso, J.; Castañeda-Babarro, A.; Seco-Calvo, J.; Urdampilleta, A. Effects of 120 g/h of Carbohydrates Intake during a Mountain Marathon on Exercise-Induced Muscle Damage in Elite Runners. Nutrients 2020, 12, 1367. [Google Scholar] [CrossRef]
- Tiller, N.B.; Roberts, J.D.; Beasley, L.; Chapman, S.; Pinto, J.M.; Smith, L.; Wiffin, M.; Russell, M.; Sparks, S.A.; Duckworth, L.; et al. International Society of Sports Nutrition Position Stand: Nutritional considerations for single-stage ultra-marathon training and racing. J. Int. Soc. Sports Nutr. 2019, 16, 50. [Google Scholar] [CrossRef] [Green Version]
- Best, R.; Barwick, B.; Best, A.; Berger, N.; Harrison, C.; Wright, M.; Sparrow, J. Changes in Pain and Nutritional Intake Modulate Ultra-Running Performance: A Case Report. Sports 2018, 6, 111. [Google Scholar] [CrossRef] [Green Version]
- Muros, J.J.; Sanchex-Munoz, C.; Hoyos, J.; Zabala, M. Nutritional intake and body composition changes in a UCI World Tour cycling team during the Tour of Spain. Eur. J. Sport Sci. 2018, 19, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Berger, N.; Cooley, D.; Graham, M.; Harrison, C.; Best, R. Physiological Responses and Nutritional Intake during a 7-Day Treadmill Running World Record. Int. J. Environ. Res. Public Health 2020, 17, 5962. [Google Scholar] [CrossRef] [PubMed]
- Wardenaar, F.C.; Dijkhuizen, R.; Ceelen, I.J.M.; Jonk, E.; Vries, J.H.M.D.; Witkamp, R.F.; Mensink, M. Nutrient Intake by Ultramarathon Runners: Can They Meet Recommendations? Int. J. Sport Nutr. Exerc. 2015, 25, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Bandegan, A.; Courtney-Martin, G.; Rafii, M.; Pencharz, P.B.; Lemon, P.W. Indicator Amino Acid-Derived Estimate of Dietary Protein Requirement for Male Bodybuilders on a Nontraining Day Is Several-Fold Greater than the Current Recommended Dietary Allowance. J. Nutr. 2017, 147, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Schoenfeld, B.J.; Aragon, A.A. How much protein can the body use in a single meal for muscle-building? Implications for daily protein distribution. J. Int. Soc. Sport Nutr. 2018, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Churchward-Venne, T.A.; Burd, N.A.; Breen, L.; Tarnopolsky, M.A.; Phillips, S.M. Myofibrillar protein synthesis following ingestion of soy protein isolate at rest and after resistance exercise in elderly men. Nutr. Metab. 2012, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Wall, B.T.; Gorissen, S.H.; Pennings, B.; Koopman, R.; Groen, B.B.L.; Verdijk, L.B.; van Loon, L.J.C. Aging Is Accompanied by a Blunted Muscle Protein Synthetic Response to Protein Ingestion. PLoS ONE 2015, 10, e0140903. [Google Scholar] [CrossRef]
- Moore, D.R.; Churchward-Venne, T.A.; Witard, O.; Breen, L.; Burd, N.A.; Tipton, K.D.; Phillips, S.M. Protein Ingestion to Stimulate Myofibrillar Protein Synthesis Requires Greater Relative Protein Intakes in Healthy Older Versus Younger Men. J. Gerontol. Ser. Biol. Sci. Med. Sci. 2014, 70, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennie, M.J.; Edwards, R.H.T.; Davies, C.T.M.; Krywawych, S.; Halliday, D.; Waterlow, J.C.; Millward, D.J. Protein and amino acid turnover during and after exercise. Biochem. Soc. Trans. 1980, 8, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.J.; Fisher, E.C.; Hoerr, R.A.; Young, V.R. Protein Metabolism and Endurance Exercise. Physician Sportsmed. 1983, 11, 63–162. [Google Scholar] [CrossRef]
- Suida, A.R. Validity of Whole and Regional Body Composition Testing Devices; Northern Illinois University: DeKalb, IL, USA, 2016. [Google Scholar]
- Buckinx, F.; Reginster, J.-Y.; Dardenne, N.; Croisiser, J.-L.; Kaux, J.-F.; Beaudart, C.; Slomian, J.; Bruyère, O. Concordance between muscle mass assessed by bioelectrical impedance analysis and by dual energy X-ray absorptiometry: A cross-sectional study. BMC Musculoskelet Disord. 2015, 16, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venables, M.C.; Achten, J.; Jeukendrup, A.E. Determinants of fat oxidation during exercise in healthy men and women: A cross-sectional study. J. Appl. Physiol. 2005, 98, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Gibala, M.J. Protein Metabolism and Endurance Exercise. Sports Med. 2007, 37, 337–340. [Google Scholar] [CrossRef]
- Gualano, A.B.; Bozza, T.; Lopes, D. Branched-chain amino acids supplementation enhances exercise capacity and lipid oxidation during endurance exercise after muscle glycogen depletion. J. Sports Med. Phys. Fit. 2011, 51, 82–88. [Google Scholar]
- Knechtle, B.; Senn, O.; Imoberdorf, R.; Joleska, I.; Wirth, A.; Knechtle, P.; Rosemann, T. Maintained Total Body Water Content and Serum Sodium Concentrations Despite Body Mass Loss in Female Ultrarunners Drinking Ad Libitum during a 100 Km Race. Asia Pac. J. Clin. Nutr. 2010, 19, 83. [Google Scholar] [PubMed]
- Beis, L.Y.; Wright-Whyte, M.; Fudge, B.; Noakes, T.; Pitsiladis, Y.P. Drinking Behaviors of Elite Male Runners during Marathon Competition. Clin. J. Sport Med. 2012, 22, 254–261. [Google Scholar] [CrossRef]
- Kipps, C.; Sharma, S.; Pedoe, D.T. The incidence of exercise-associated hyponatraemia in the London marathon. Brit. J. Sport Med. 2011, 45, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, M.D.; Krishnan, E. Health and Exercise-Related Medical Issues among 1212 Ultramarathon Runners: Baseline Findings from the Ultrarunners Longitudinal TRAcking (ULTRA) Study. PLoS ONE 2014, 9, e83867. [Google Scholar] [CrossRef] [Green Version]
- Goedecke, J.H.; Gibson, A.S.C.; Grobler, L.; Collins, M.; Noakes, T.D.; Lambert, E.V. Determinants of the variability in respiratory exchange ratio at rest and during exercise in trained athletes. Am. J. Physiol.-Endocrinol. Metab. 2000, 279, E1325–E1334. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helge, J.; Fraser, A.; Kriketos, A.; Jenkins, A.; Calvert, G.; Ayre, K.; Storlien, L. Interrelationships between muscle fibre type, substrate oxidation and body fat. Int. J. Obes. 1999, 23, 986–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knufinke, M.; Nieuwenhuys, A.; Geurts, S.A.E.; Møst, E.I.S.; Maase, K.; Moen, M.H.; Coenen, A.M.L.; Kompier, M.A.J. Train hard, sleep well? Perceived training load, sleep quantity and sleep stage distribution in elite level athletes. J. Sci. Med. Sport 2018, 21, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Urdampilleta, A.; Arribalzaga, S.; Viribay, A.; Castañeda-Babarro, A.; Seco-Calvo, J.; Mielgo-Ayuso, J. Effects of 120 vs. 60 and 90 g/h Carbohydrate Intake during a Trail Marathon on Neuromuscular Function and High Intensity Run Capacity Recovery. Nutrients 2020, 12, 2094. [Google Scholar] [CrossRef] [PubMed]
- Brueckner, J.C.; Atchou, G.; Capelli, C.; Duvallet, A.; Barrault, D.; Jousselin, E.; Rieu, M.; Prampero, P.E.d. The energy cost of running increases with the distance covered. Eur. J. Appl. Physiol. 1991, 62, 385–389. [Google Scholar] [CrossRef]
- Daniels, J.; Daniels, N. Running economy of elite male and elite female runners. Med. Sci. Sports Exerc. 1992, 24, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J. Modeling: Optimal marathon performance on the basis of physiological factors. J. Appl. Physiol. 1991, 70, 683–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, C.; Lucia, A. Running economy: The forgotten factor in elite performance. Sports Med. 2007, 37, 316–319. [Google Scholar] [CrossRef]
- Barstow, T.J.; Mole, P.A. Linear and nonlinear characteristics of oxygen uptake kinetics during heavy exercise. J. Appl. Physiol. 1991, 71, 2099–2106. [Google Scholar] [CrossRef]
- Paterson, D.H.; Whipp, B.J. Asymmetries of oxygen uptake transients at the on- and offset of heavy exercise in humans. J. Physiol. 1991, 443, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Bearden, S.E.; Moffatt, R.J. VO(2) kinetics and the O2 deficit in heavy exercise. J. Appl. Physiol. 2000, 88, 1407–1412. [Google Scholar] [CrossRef] [Green Version]
- Roebuck, G.S.; Fitzgerald, P.B.; Urquhart, D.M.; Ng, S.-K.; Cicuttini, F.M.; Fitzgibbon, B.M. The psychology of ultra-marathon runners: A systematic review. Psychol. Sport Exerc. 2018, 37, 43–58. [Google Scholar] [CrossRef]
- Buffa, R.; Mereu, E.; Comandini, O.; Ibanez, M.E.; Marini, E. Bioelectrical impedance vector analysis (BIVA) for the assessment of two-compartment body composition. Eur. J. Clin. Nutr. 2014, 68, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Castizo-Olier, J.; Irurtia, A.; Jemni, M.; Carrasco-Marginet, M.; Fernandez-Garcia, R.; Rodriguez, F.A. Bioelectrical impedance vector analysis (BIVA) in sport and exercise: Systematic review and future perspectives. PLoS ONE 2018, 13, e0197957. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marathon | Completion Time |
|---|---|
| (h, min, s) | |
| 1 | 4 h 21 min 21 s |
| 2 | 4 h 21 min 39 s |
| 3 | 4 h 24 min 38 s |
| 4 | 4 h 24 min 06 s |
| 5 | 4 h 23 min 55 s |
| 6 | 4 h 24 min 22 s |
| 7 | 4 h 23 min 54 s |
| 8 | 4 h 23 min 21 s |
| 9 | 4 h 23 min 25 s |
| 10 | 4 h 21 min 21 s |
| Mean 1 | 4 h 23 min 9 s |
| Total | 43 h 51 min 39 s |
| Marathon | Heart Rate ± SD |
|---|---|
| (b.min−1) | |
| 1 | 139 ± 10 |
| 2 | 140 ± 5 |
| 3 | 136 ± 9 |
| 4 | 146 ± 5 |
| 5 | 148 ± 10 |
| 6 | 142 ± 4 |
| 7 | 141 ± 7 |
| 8 | 142 ± 9 |
| 9 | 150 ± 8 |
| 10 | 144 ± 6 |
| Mean 2 | 143 ± 4 |
| Hour of Marathon | Heart Rate ± SD |
|---|---|
| (b.min−1) | |
| 1 | 137 ± 6 |
| 2 | 137 ± 5 |
| 3 3 | 145 ± 6 |
| 4 | 151 ± 5 |
| Mean | 143 ± 4 |
| Breakfast | |
|---|---|
| Weetabix (oats) | 2 |
| Semi skimmed milk | 100 mL |
| Sugar | 2 teaspoons |
| Tea | 200 mL |
| Coffee (instant) | 200 mL |
| Actimel probiotic yoghurt | 100 mL |
| 10 a.m. | |
| Chicken cup soup | 1 |
| Bread, whole meal | 1 slice |
| Event | |
| Grapes | 16 |
| Water | 1750 mL |
| Post-marathon | |
| Semi skimmed milk | 500 mL |
| Mountain fuel recovery powder | 2 scoops |
| Neovite colostrum powder | 1 teaspoon |
| Protein PHD pharma whey protein | 35 g |
| Banana | 1 |
| Evening | |
| lamb, roast potatoes, parsnips, carrots, green beans, leaks, gravy | |
| apple crumble and double cream | |
| Tea | 600 mL |
| Bed | |
| Milk | 200 mL |
| Sugar | 1 teaspoon |
| Instant drinking chocolate | 1 teaspoon |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, N.; Cooley, D.; Graham, M.; Harrison, C.; Campbell, G.; Best, R. Consistency Is Key When Setting a New World Record for Running 10 Marathons in 10 Days. Int. J. Environ. Res. Public Health 2021, 18, 12066. https://doi.org/10.3390/ijerph182212066
Berger N, Cooley D, Graham M, Harrison C, Campbell G, Best R. Consistency Is Key When Setting a New World Record for Running 10 Marathons in 10 Days. International Journal of Environmental Research and Public Health. 2021; 18(22):12066. https://doi.org/10.3390/ijerph182212066
Chicago/Turabian StyleBerger, Nicolas, Daniel Cooley, Michael Graham, Claire Harrison, Georgia Campbell, and Russ Best. 2021. "Consistency Is Key When Setting a New World Record for Running 10 Marathons in 10 Days" International Journal of Environmental Research and Public Health 18, no. 22: 12066. https://doi.org/10.3390/ijerph182212066
APA StyleBerger, N., Cooley, D., Graham, M., Harrison, C., Campbell, G., & Best, R. (2021). Consistency Is Key When Setting a New World Record for Running 10 Marathons in 10 Days. International Journal of Environmental Research and Public Health, 18(22), 12066. https://doi.org/10.3390/ijerph182212066