Phenological Response in the Trophic Levels to Climate Change in Korea



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Selection and Classification
2.2. Data and Analysis
3. Results
3.1. Phenological Shift of Trophic Levels
3.2. Phenological Temperature Sensitivity
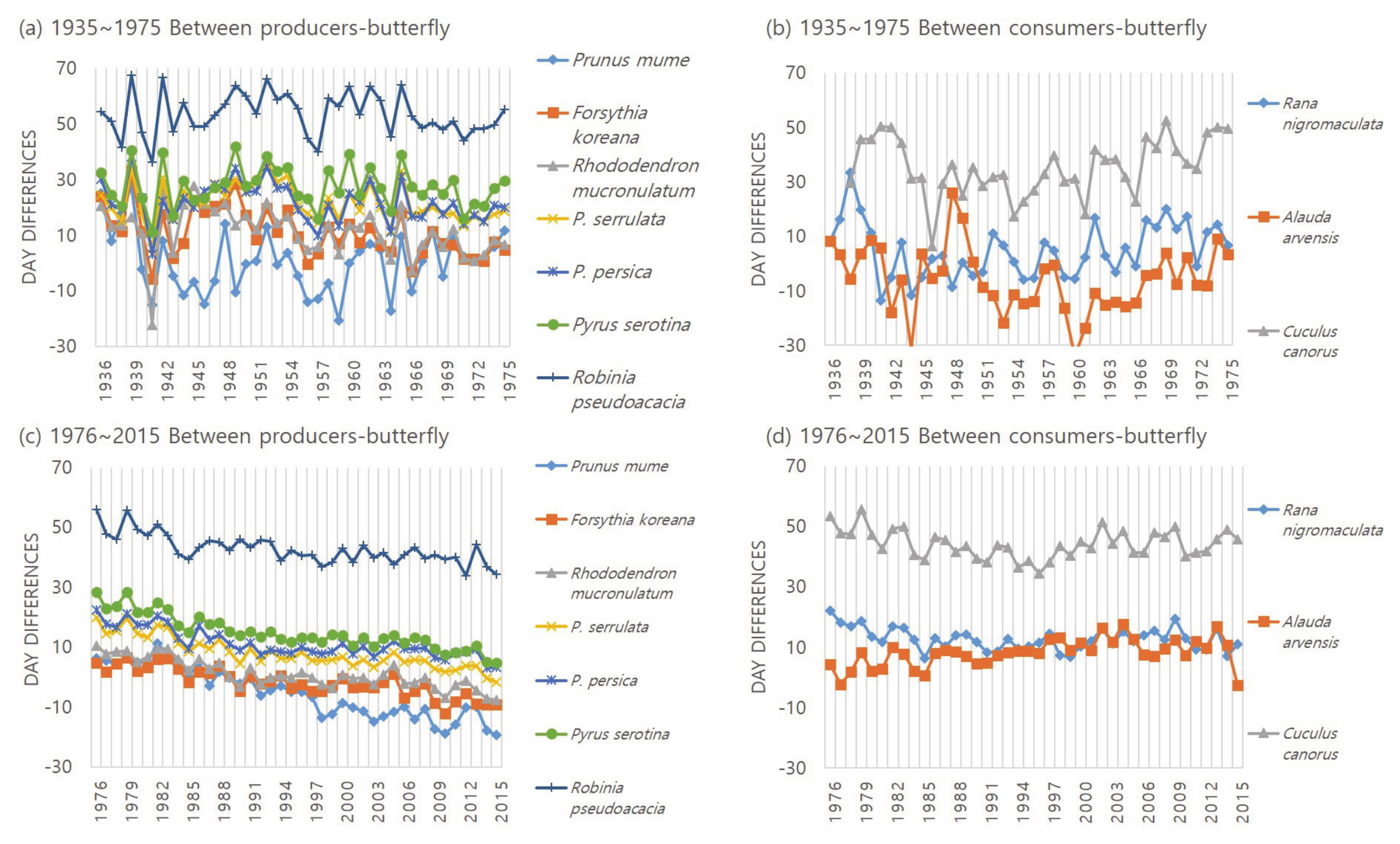
3.3. Change of Day Difference between Interacting Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Korea Meteorological Administration. Korean Climate Change Assessment Report 2020; Korea Meteorological Administration: Seoul, Korea, 2020. [Google Scholar]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How many species are there on earth and in the ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.D.; Kwon, S.-S. Carbon sequestration in the urban areas of Seoul with climate change: Implication for open innovation in environmental industry. J. Open Innov. Technol. Mark. Complex. 2018, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Pounds, J.A.; Puschendorf, R. Ecology: Clouded futures. Nature 2004, 427, 107–109. [Google Scholar] [CrossRef]
- Visser, M.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. B. 2005, 272, 2561–2569. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Mysterud, A. Climate, changing phenology, and other life history traits: Nonlinearity and match-mismatch to the environment. Proc. Natl. Acad. Sci. USA 2002, 99, 13379–13381. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E.; Both, C.; Lambrechts, M.M. Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 2004, 35, 89–110. [Google Scholar] [CrossRef] [Green Version]
- Thakur, M.P. Climate warming and trophic mismatches in terrestrial ecosystems: The green–brown imbalance hypothesis. Biol. Lett. 2020, 16, 20190770. [Google Scholar] [CrossRef] [Green Version]
- Thackeray, S.J.; Henrys, P.A.; Hemming, D.; Bell, J.R.; Botham, M.S.; Burthe, S.; Helaouet, P.; Johns, D.G.; Jones, I.D.; Leech, D.I.; et al. Phenological sensitivity to climate across taxa and trophic levels. Nature 2016, 535, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Kharouba, H.M.; Wolkovich, E.M. Disconnects between ecological theory and data in phenological mismatch research. Nat. Clim. Chang. 2020, 10, 406–415. [Google Scholar] [CrossRef]
- Walther, G.-R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Ibáñez, I.; Primack, R.B.; Miller-Rushing, A.J.; Ellwood, E.; Higuchi, H.; Lee, S.D.; Kobori, H.; Silander, J.A. Forecasting phenology under global warming. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3247–3260. [Google Scholar] [CrossRef]
- Charmantier, A.; Mccleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef] [Green Version]
- Both, C.; Van Asch, M.; Bijlsma, R.G.; Burg, A.B.V.D.; Visser, M.E. Climate change and unequal phenological changes across four trophic levels: Constraints or adaptations? J. Anim. Ecol. 2009, 78, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Bauer, Z.; Trnka, M.; Bauerová, J.; Možný, M.; Štěpánek, P.; Bartošová, L.; Zalud, Z. Changing climate and the phenological response of great tit and collared flycatcher populations in floodplain forest ecosystems in Central Europe. Int. J. Biometeorol. 2010, 54, 99–111. [Google Scholar] [CrossRef]
- Thackeray, S.J.; Sparks, T.H.; Frederiksen, M.; Burthe, S.; Bacon, P.J.; Bell, J.R.; Botham, M.S.; Brereton, T.M.; Bright, P.W.; Carvalho, L.; et al. Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments. Glob. Chang. Biol. 2010, 16, 3304–3313. [Google Scholar] [CrossRef] [Green Version]
- Kharouba, H.M.; Ehrlén, J.; Gelman, A.; Bolmgren, K.; Allen, J.M.; Travers, S.E.; Wolkovich, E.M. Global shifts in the phenological synchrony of species interactions over recent decades. Proc. Natl. Acad. Sci. USA 2018, 115, 5211–5216. [Google Scholar] [CrossRef] [Green Version]
- Burthe, S.J.; Daunt, F.; Butler, A.; Elston, D.; Frederiksen, M.; Johns, D.; Newell, M.; Thackeray, S.; Wanless, S. Phenological trends and trophic mismatch across multiple levels of a North Sea pelagic food web. Mar. Ecol. Prog. Ser. 2012, 454, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Miller-Rushing, A.J.; Høye, T.T.; Inouye, D.W.; Post, E. The effects of phenological mismatches on demography. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3177–3186. [Google Scholar] [CrossRef] [Green Version]
- Forrest, J.; Miller-Rushing, A.J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos. Trans. R. Soc. B 2010, 365, 3101–3112. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.D. Global warming leading to phenological responses in the process of urbanization, South Korea. Sustainability 2017, 9, 2203. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yi, Q.; Xing, F.; Tang, C.; Wang, L.; Ye, W.; Ng, I.I.; Chan, T.I.; Chen, H.; Liu, D. Rapid shifts of peak flowering phenology in 12 species under the effects of extreme climate events in Macao. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Kharouba, H.M.; Vellend, M. Flowering time of butterfly nectar food plants is more sensitive to temperature than the timing of butterfly adult flight. J. Anim. Ecol. 2015, 84, 1311–1321. [Google Scholar] [CrossRef]
- Fu, Y.; Piao, S.; Ciais, P.; Huang, M.; Menzel, A.; Peaucelle, M.; Peng, S.; Song, Y.; Vitasse, Y.; Zeng, Z.; et al. Long-term linear trends mask phenological shifts. Int. J. Biometeorol. 2016, 60, 1611–1613. [Google Scholar] [CrossRef] [Green Version]
- Korea Meteorological Administration. Full Text of Seasonal Observation Guidelines; Korea Meteorological Administration: Seoul, Korea, 2016. [Google Scholar]
- Cooke, P. Green governance and green clusters: Regional & national policies for the climate change challenge of Central & Eastern Europe. J. Open Innov. Technol. Mark. Complex. 2015, 1, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.D. Wintering habitat use pattern of red-crowned cranes in the Korean demilitarized zone. J. Open Innov. Technol. Mark. Complex. 2018, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Primack, R.B.; Ibanez, I.; Higuchi, H.; Lee, S.D.; Miller-Rushing, A.J.; Wilson, A.M.; Silander, J.A. Spatial and inter-specific vaiability in phenological responses to warming temperatures. Biol. Conserv. 2009, 142, 2569–2577. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Katsuki, T.; Primack, R.B.; Ishii, Y.; Lee, S.D.; Higuchi, H. Impact of global warming on a group of related species and their hybrids: Cherry tree (Rosaceae) flowering at Mt. Takao, Japan. Am. J. Bot. 2007, 94, 1470–1478. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.B.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef]
- Inouye, D.W.; Barr, B.; Armitage, K.B.; Inouye, B.D. Climate change is affecting altitudinal migrants and hibernating species. Proc. Natl. Acad. Sci. USA 2000, 97, 1630–1633. [Google Scholar] [CrossRef] [Green Version]
- Post, E.; Forchhammer, M.C. Climate change reduces reproductive success of an Arctic herbivore through trophic mismatch. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 2367–2373. [Google Scholar] [CrossRef] [Green Version]
- Ovaskainen, O.; Skorokhodova, S.; Yakovleva, M.; Sukhov, A.; Kutenkov, A.; Kutenkova, N.; Shcherbakov, A.; Meyke, E.; Delgado, M.D.M. Community-level phenological response to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 13434–13439. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E. Interactions of climate change and species. Nature 2016, 535, 236–237. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Cambrollé, J.; Hidalgo-Galvez, M.D.; Matías, L.; Montero-Ramírez, A.; Santolaya, S.; Godoy, O. Phenological responses to climate change in communities of plants species with contrasting functional strategies. Environ. Exp. Bot. 2020, 170, 103852. [Google Scholar] [CrossRef]
- Lee, S.-D.; Ellwood, E.R.; Park, S.-Y.; Primack, R.B. Late-arriving barn swallows linked to population declines. Biol. Conserv. 2011, 144, 2182–2187. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Species | Phenological Event | Mean Day of Event (Julian Day) | Trophic Level | |
|---|---|---|---|---|---|
| Common Name | Scientific Name | ||||
| Flora | Plum tree | Prunus mume | FFD | 83.13 (SD = 19.12, n = 1898) | Producer (n = 7) |
| Korean forsythia | Forsythia koreana | FFD | 87.26 (SD = 8.32, n = 2065) | ||
| Korean rosebay | Rhododendron mucronulatum | FFD | 89.92 (SD = 9.05, n = 2018) | ||
| Cherry tree | P. yedoensis | FFD | 96.84 (SD = 7.97, n = 2067) | ||
| Peach tree | P. persica | FFD | 99.87 (SD = 9.80, n = 2003) | ||
| Pear tree | Pyrus serotina | FFD | 103.57 (SD = 8.89, n = 1890) | ||
| Black locust | Robinia pseudoacacia | FFD | 131.53 (SD = 6.84, n = 1976) | ||
| Insect | Cabbage butterfly | Pieris rapae | FAD | 88.55 (SD = 12.54, n = 2021) | Primary consumer (n = 1) |
| Amphibian | Black-spotted pond frog | Rana nigromaculata | FAD | 101.18 (SD = 12.97, n = 1850) | Secondary consumer (n = 3) |
| Avian | Skylark | Alauda arvensis | FSD | 96.74 (SD = 19.45, n = 1514) | |
| Cuckoo | Cuculus canorus | FSD | 132.56 (SD = 13.98, n = 1787) | ||
| Tropic Level | Species | Trend to Earlier Spring Phenology (−) | Trend to Later Spring Phenology (+) |
|---|---|---|---|
| Producer | Prunus mume | 55 (36) | 6 (1) |
| Forsythia koreana | 53 (16) | 8 (0) | |
| Rhododendron mucronulatum | 46 (18) | 15 (0) | |
| P. yedoensis | 57 (32) | 4 (0) | |
| P. persica | 51 (16) | 10 (0) | |
| Pyrus serotine | 51 (24) | 10 (0) | |
| Robinia pseudoacacia | 47 (14) | 14 (2) | |
| Primary consumer | Pieris rapae | 16 (3) | 45 (16) |
| Secondary consumer | Rana nigromaculata | 26 (10) | 35 (13) |
| Alauda arvensis | 16 (4) | 45 (29) | |
| Cuculus canorus | 20 (3) | 41 (14) |
| Tropic Level | Species | 1976~2015 | 1936~1975 | ||||
|---|---|---|---|---|---|---|---|
| Slope | p-Value | R2 | Slope | p-Value | R2 | ||
| Producer | Prunus mume | −7.20 | 0.00 | 0.74 | −5.83 | 0.00 | 0.22 |
| Forsythia koreana | −4.37 | 0.00 | 0.71 | −5.27 | 0.00 | 0.53 | |
| Rhododendron mucronulatum | −4.76 | 0.00 | 0.79 | −5.13 | 0.00 | 0.40 | |
| P. serrulata | −4.61 | 0.00 | 0.83 | −3.60 | 0.00 | 0.66 | |
| P. persica | −4.27 | 0.00 | 0.81 | −4.18 | 0.00 | 0.51 | |
| Pyrus serotina | −4.45 | 0.00 | 0.83 | −2.53 | 0.00 | 0.28 | |
| Robinia pseudoacacia | −3.13 | 0.00 | 0.62 | −1.37 | 0.02 | 0.13 | |
| Primary consumer | Pieris rapae | −1.92 | 0.00 | 0.13 | −2.82 | 0.04 | 0.11 |
| Secondary consumer | Rana nigromaculata | −1.92 | 0.00 | 0.21 | −1.62 | 0.30 | 0.03 |
| Alauda arvensis | −0.08 | 0.95 | 0.00 | −3.10 | 0.11 | 0.07 | |
| Cuculus canorus | −0.16 | 0.79 | 0.00 | −1.96 | 0.29 | 0.03 | |
| Day Difference | Species | 1976~2015 | 1936~1975 | ||||
|---|---|---|---|---|---|---|---|
| Slope | p-Value | R2 | Slope | p-Value | R2 | ||
| Producer-Primary consumer | Prunus mume | −0.65 | 0.00 | 0.85 | −0.27 | 0.08 | 0.08 |
| Forsythia koreana | −0.33 | 0.00 | 0.74 | −0.54 | 0.00 | 0.46 | |
| Rhododendron mucronulatum | −0.35 | 0.00 | 0.73 | −0.34 | 0.01 | 0.16 | |
| P. serrulata | −0.40 | 0.00 | 0.79 | −0.33 | 0.00 | 0.30 | |
| P. persica | −0.32 | 0.00 | 0.68 | −0.28 | 0.00 | 0.27 | |
| Pyrus serotina | −0.43 | 0.00 | 0.81 | −0.06 | 0.55 | 0.01 | |
| Robinia pseudoacacia | −0.31 | 0.00 | 0.55 | −0.09 | 0.40 | 0.02 | |
| Secondary consumer-Primary consumer | Rana nigromaculata | −0.08 | 0.13 | 0.06 | 0.09 | 0.43 | 0.02 |
| Alauda arvensis | 0.25 | 0.00 | 0.40 | −0.19 | 0.28 | 0.03 | |
| Cuculus canorus | −0.07 | 0.32 | 0.03 | 0.16 | 0.32 | 0.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Lee, S.; Lee, H.; Lee, S. Phenological Response in the Trophic Levels to Climate Change in Korea. Int. J. Environ. Res. Public Health 2021, 18, 1086. https://doi.org/10.3390/ijerph18031086
Kim M, Lee S, Lee H, Lee S. Phenological Response in the Trophic Levels to Climate Change in Korea. International Journal of Environmental Research and Public Health. 2021; 18(3):1086. https://doi.org/10.3390/ijerph18031086
Chicago/Turabian StyleKim, Minkyung, Sojeong Lee, Hakyung Lee, and Sangdon Lee. 2021. "Phenological Response in the Trophic Levels to Climate Change in Korea" International Journal of Environmental Research and Public Health 18, no. 3: 1086. https://doi.org/10.3390/ijerph18031086
APA StyleKim, M., Lee, S., Lee, H., & Lee, S. (2021). Phenological Response in the Trophic Levels to Climate Change in Korea. International Journal of Environmental Research and Public Health, 18(3), 1086. https://doi.org/10.3390/ijerph18031086