
Photosynthesis and Related Physiological Parameters Differences Affected the Isoprene Emission Rate among 10 Typical Tree Species in Subtropical Metropolises

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
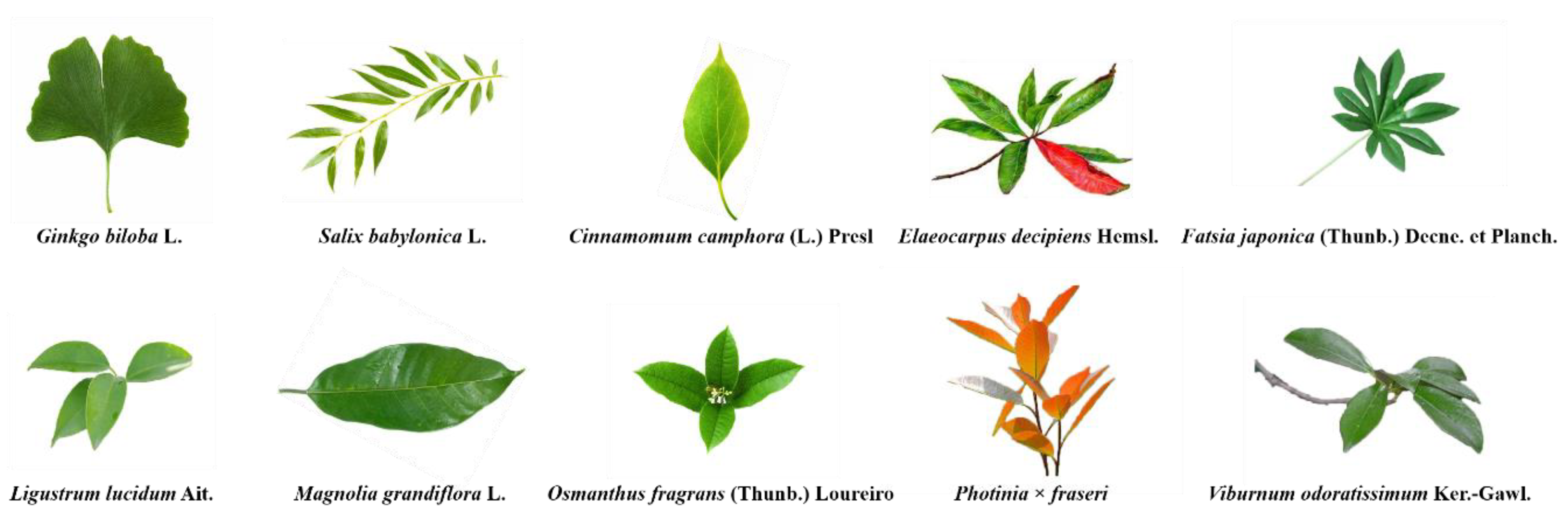
2.1. Sampling Sites and Tree Species
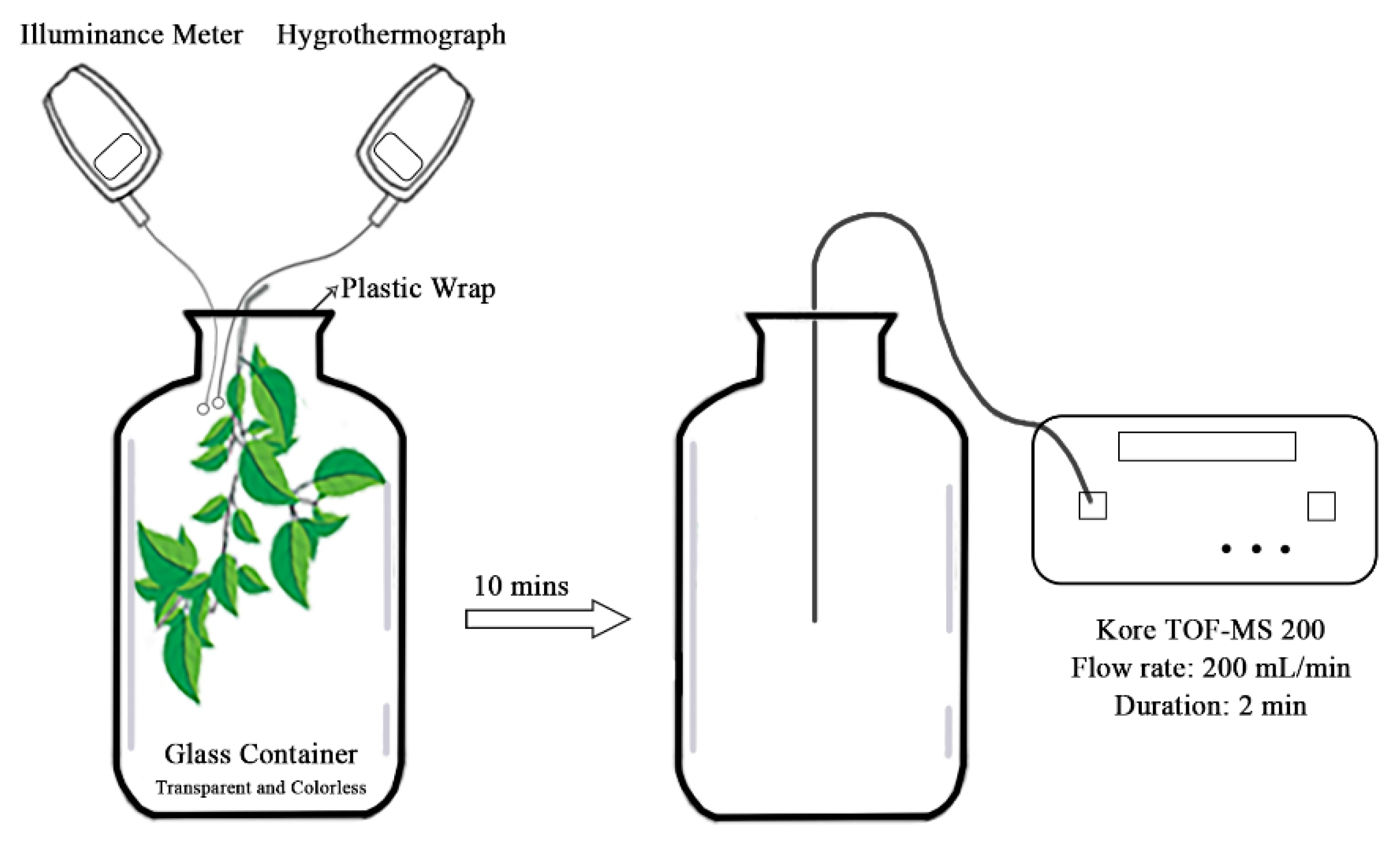
2.2. Measurement of Isoprene Emission Rate
2.3. Standardization of the Isoprene Emission Rate
2.4. Measurement of Plant Photosynthesis and Related Physiological Parameters
2.5. Data Processing
3. Results
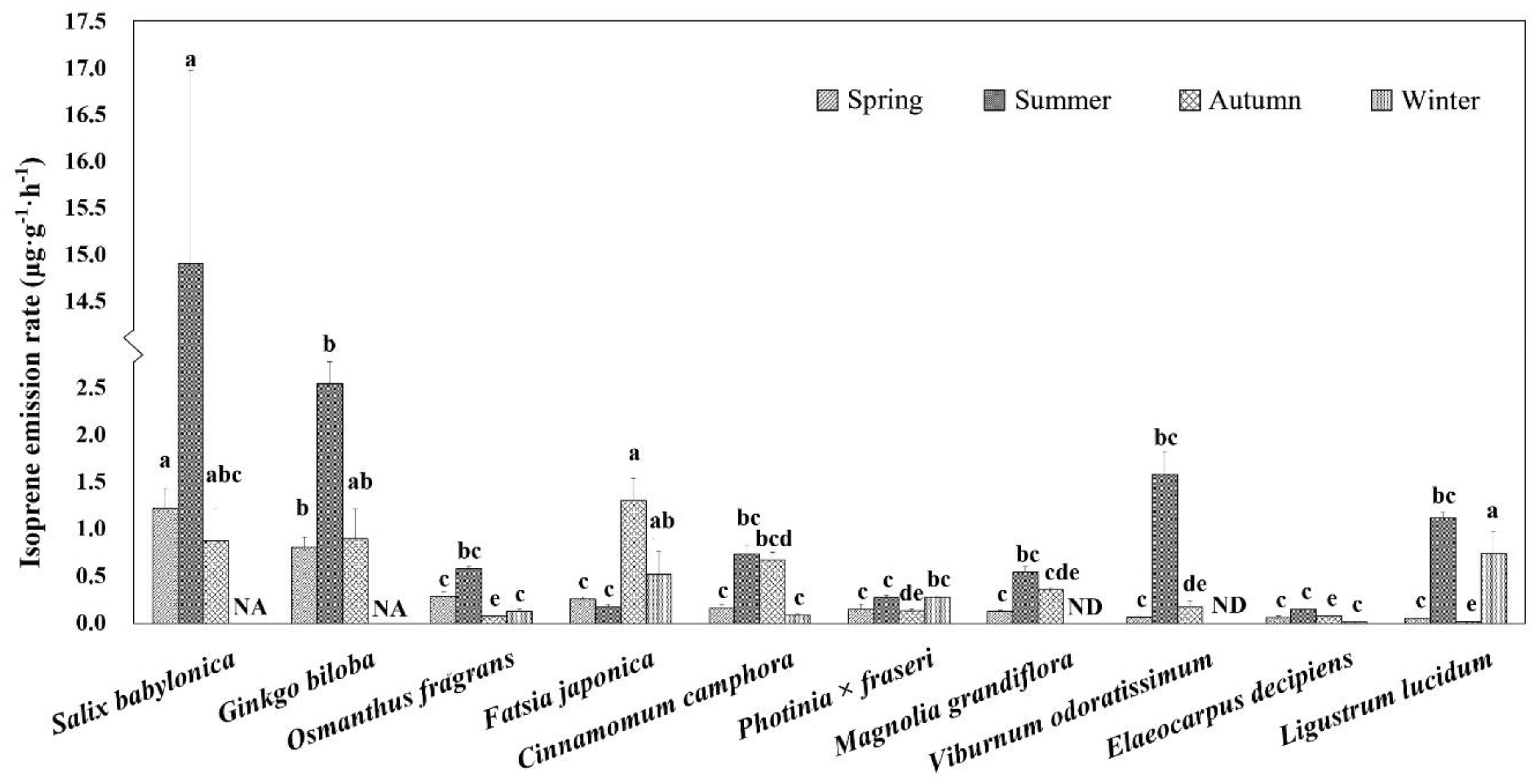
3.1. Isoprene Emission Rate of Different Trees in Four Seasons
3.2. Photosynthesis and Related Physiological Parameters of Different Plants in Growing Seasons
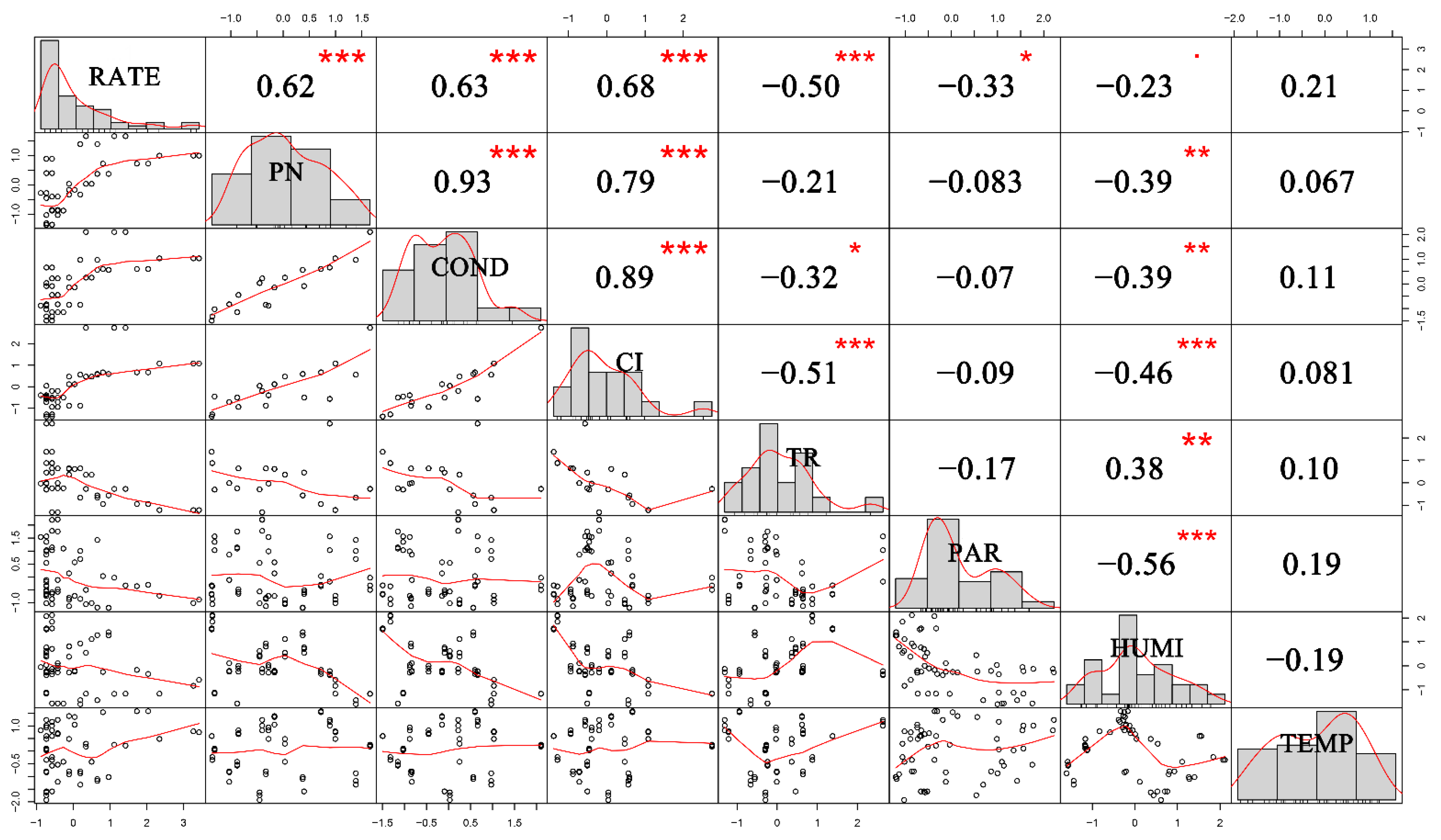
3.3. Correlations between Plant Parameters and Isoprene Emission Rate
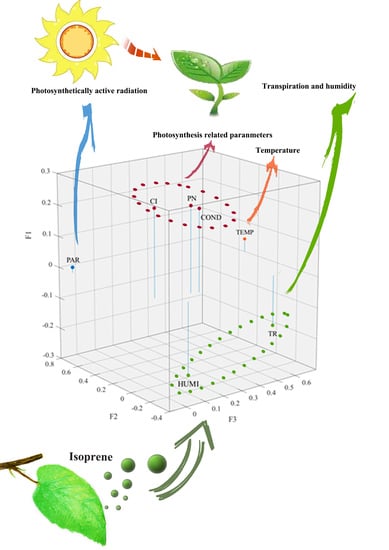
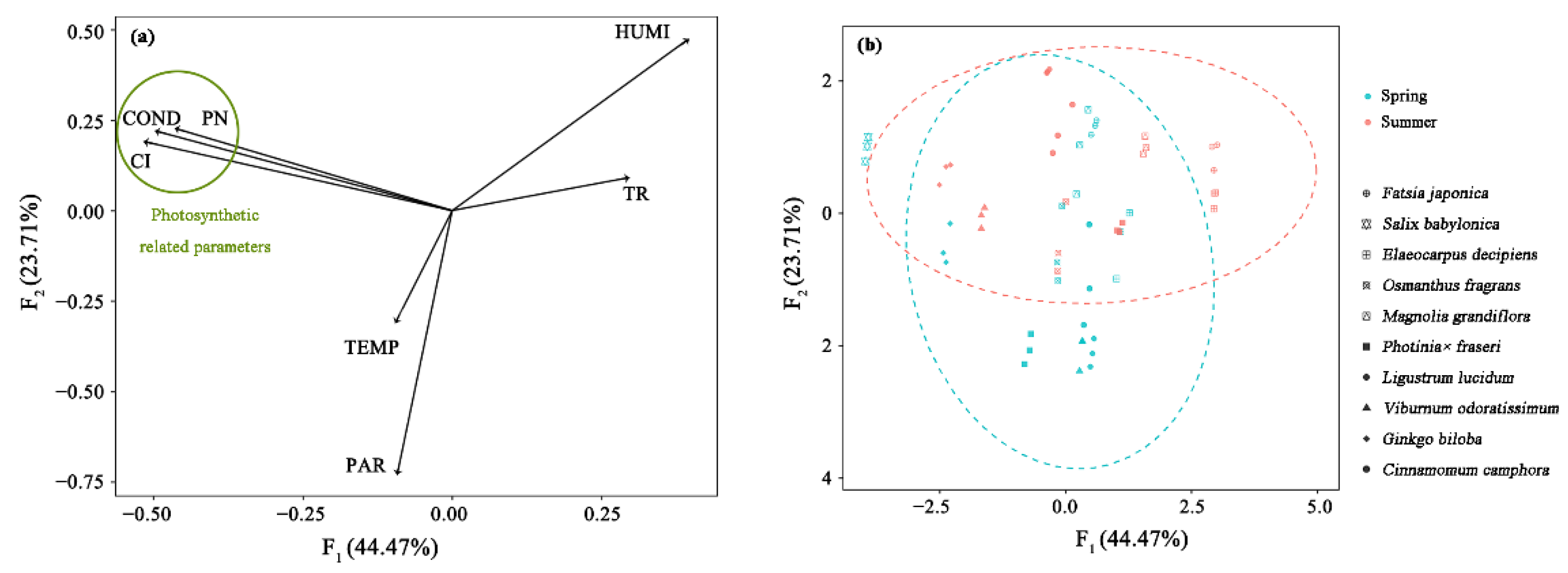
3.4. Key Factors of Isoprene Emission Rate Based on PCA
4. Discussion
4.1. Close Relationship between Photosynthesis Related Parameters and Isoprene Emission
4.2. Other Factors Influencing Isoprene Emission
4.3. Species Selection and Priority Control
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Escobedo, F.J.; Adams, D.C.; Timilsina, N. Urban forest structure effects on property value. Ecosyst. Serv. 2015, 12, 209–217. [Google Scholar] [CrossRef]
- Willis, K.J.; Petrokofsky, G. The natural capital of city trees. Sciecce 2017, 356, 6336. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Monson, R.K. Isoprene research—60 years later, the biology is still enigmatic. Plant Cell Environ. 2017, 40, 1671–1678. [Google Scholar] [CrossRef]
- Iijima, Y. Recent Advances in the Application of Metabolomics to Studies of Biogenic Volatile Organic Compounds (BVOC) Produced by Plant. Metabolites 2014, 4, 699–721. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Llusià, J. Plant VOC emissions: Making use of the unavoidable. Trends Ecol. Evol. 2004, 19, 402–404. [Google Scholar] [CrossRef]
- Guenther, A.B.; Hewitt, C.N.; Erickson, D.; Fall, R.; Geron, C.; Graedel, T.; Harley, P.; Klinger, L.; Lerdau, M.; Mckay, W.A. A global model of natural volatile organic compound emissions. J. Geophys. Res. Atmos. 1995, 100, 8873–8892. [Google Scholar] [CrossRef]
- Simpson, D.; Winiwarter, W.; Börjesson, G.; Cinderby, S.; Ferreiro, A.; Guenther, A.; Hewitt, C.N.; Janson, R.; Khalil, M.A.K.; Owen, S. Inventorying emissions from nature in Europe. J. Geophys. Res. Atmos. 1999, 104, 8113–8152. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Ren, Y.; Shi, Y.; Zhu, Y.; Ge, Y.; Hong, S.; Jiao, L.; Lin, F.; Peng, C.; Mochizuki, T.; et al. An inventory of biogenic volatile organic compounds for a subtropical urban–rural complex. Atmos. Environ. 2012, 56, 115–123. [Google Scholar] [CrossRef]
- Pacheco, C.K.; Fares, S.; Ciccioli, P. A highly spatially resolved GIS-based model to assess the isoprenoid emissions from key Italian ecosystems. Atmos. Environ. 2014, 96, 50–60. [Google Scholar] [CrossRef]
- Loreto, F.; Dicke, M.; Schnitzler, J.-P.; Turlings, T. Plant volatiles and the environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef]
- Ren, Y.; Qu, Z.; Du, Y.; Xu, R.; Ma, D.; Yang, G.; Shi, Y.; Fan, X.; Akira, T.; Guo, P.; et al. Air quality and health effects of biogenic volatile organic compounds emissions from urban green spaces and the mitigation strategies. Environ. Pollut. 2017, 230, 849–861. [Google Scholar] [CrossRef]
- Li, J. Emission of Two Tree Species of Platycladus orientalis and Pinus tabulaeformis in Urban Environment. Ph.D. Thesis, China Academy of Forestry Sciences, Beijing, China, 2009. (In Chinese). [Google Scholar]
- Llusia, J.; Roahtyn, S.; Yakir, D.; Rotenberg, E.; Seco, R.; Guenther, A.; Peñuelas, J. Photosynthesis, stomatal conductance and terpene emission response to water availability in dry and mesic Mediterranean forests. Trees 2016, 30, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Brilli, F.; Tsonev, T.; Mahmood, T.; Velikova, V.; Loreto, F.; Centritto, M. Ultradian variation of isoprene emission, photosynthesis, mesophyll conductance, and optimum temperature sensitivity for isoprene emission in water-stressed Eucalyptus citriodora saplings. J. Exp. Bot. 2013, 64, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Huve, K.; Christ, M.; Kleist, E.; Uerlings, R.; Niinemets, U.; Walter, A.; Wildt, J. Simultaneous growth and emission measurements demonstrate an interactive control of methanol release by leaf expansion and stomata. J. Exp. Bot. 2007, 58, 1783–1793. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Wiberley, A.E.; Donohue, A.R. Isoprene Emission from Plants: Why and How. Ann. Bot. 2008, 101, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Sharkey, T.D. A gas-exchange study of photosynthesis and isoprene emission in Quercus rubra L. Planta 1990, 182, 523–531. [Google Scholar] [CrossRef]
- Loreto, F.; Bagnoli, F.; Fineschi, S. One species, many terpenes: Matching chemical and biological diversity. Trends Plant Sci. 2009, 14, 416–420. [Google Scholar] [CrossRef]
- Singh, A.P.; Varshney, C.K.; Singh, U.K. Seasonal Variations in Isoprene Emission from Tropical Deciduous Tree Species. Environ. Monit. Assess. 2007, 131, 231–235. [Google Scholar] [CrossRef]
- Karl, M.; Guenther, A.; Köble, R.; Leip, A.; Seufert, G. A new European plant-specific emission inventory of biogenic volatile organic compounds for use in atmospheric transport models. Biogeosciences 2009, 6, 1059–1087. [Google Scholar] [CrossRef]
- Aydin, Y.M.; Yaman, B.; Koca, H.; Dasdemir, O.; Kara, M.; Altiok, H.; Dumanoglu, Y.; Bayram, A.; Tolunay, D.; Odabasi, M.; et al. Biogenic volatile organic compound (BVOC) emissions from forested areas in Turkey: Determination of specific emission rates for thirty-one tree species. Sci. Total Environ. 2014, 490, 239–253. [Google Scholar] [CrossRef]
- Fitzky, A.C.; Sandén, H.; Kar, T.; Fares, S.; Calfapietra, C.; Grote, R.; Saunier, A.; Rewald, B. The Interplay Between Ozone and Urban Vegetation—BVOC Emissions, Ozone Deposition, and Tree Ecophysiology. Front. For. Glob. Chang. 2019, 2, 1–17. [Google Scholar] [CrossRef]
- Djen, C.S. The urban climate of Shanghai. Atmos. Environ. Part B Urban Atmos. 1992, 26, 9–15. [Google Scholar] [CrossRef]
- Hao, R. Analysis and evaluation of soil fertility characteristics of Shanghai urban green area. Acta Agric. Shanghai 2014, 30, 79–84. [Google Scholar]
- Xiong, F.; Lyu, J.; Zhou, P.; Yin, S. lsoprene emission rate of five plant species based on in-situ time-of-flight mass spectrometry. J. Shanghai Jiaotong Univ. 2019, 37, 89–94. (In Chinese) [Google Scholar]
- Pier, P.A. Isoprene emission rates from northern red oak using a whole-tree chamber. Atmos. Environ. 1995, 29, 1347–1353. [Google Scholar] [CrossRef]
- Christof, S.; Bettina, D.; Thomas, K.; Crowley, J.N.; Jonathan, W. Glyoxal measurement with a proton transfer reaction time of flight mass spectrometer (PTR-TOF-MS): Characterization and calibration. J. Mass Spectrom. JMS 2017, 52, 30–35. [Google Scholar]
- Taiti, C.; Costa, C.; Nissim, W.G.; Bibbiani, S.; Azzarello, E.; Masi, E.; Pandolfi, C.; Pallottino, F.; Menesatti, P.; Mancuso, S. Assessing VOC emission by different wood cores using the PTR-ToF-MS technology. Wood Sci. Technol. 2017, 51, 273–295. [Google Scholar] [CrossRef]
- Guenther, A.B.; Zimmerman, P.R.; Wildermuth, M. Natural volatile organic compound emission rate estimates for U.S. woodland landscapes. Atmos. Environ. 1994, 28, 1197–1210. [Google Scholar]
- Jardine, K.J.; Zorzanelli, R.F.; Gimenez, B.O.; Oliveira Piva, L.R.d.; Teixeira, A.; Fontes, C.G.; Robles, E.; Higuchi, N.; Chambers, J.Q.; Martin, S.T. Leaf isoprene and monoterpene emission distribution across hyperdominant tree genera in the Amazon basin. Phytochemistry 2020, 175, 112366. [Google Scholar] [CrossRef]
- Guenther, A.B.; Zimmerman, P.R.; Harley, P.C.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Model evaluations and sensitivity analyses. J. Geophys. Res. Atmos. 1993, 98, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Sanadze, G.A. Biogenic Isoprene (A Review). Russ. J. Plant Physiol. 2004, 51, 729–741. [Google Scholar] [CrossRef]
- Monson, R.K.; Jaeger, C.H.; Adams, W.W.; Driggers, E.M.; Silver, G.M.; Fall, R. Relationships among Isoprene Emission Rate, Photosynthesis, and Isoprene Synthase Activity as Influenced by Temperature. Plant Physiol. 2012, 98, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.D.; Li, H.; Rauchfuss, T.B.; Bénard, M.; Rohmer, M.M. Diiron Azadithiolates as Models for the Iron-Only Hydrogenase Active Site: Synthesis, Structure, and Stereoelectronics This research was supported by the NIH and the Centre Universitaire et Regional de Ressources Informatiques of ULP and CNRS. Angew. Chem. Int. Ed. Engl. 2001, 40, 1768–1771. [Google Scholar] [CrossRef]
- Owen, S.M.; Harley, P.; Guenther, A.; Hewitt, C.N. Light dependency of VOC emissions from selected Mediterranean plant species. Atmos. Environ. 2002, 36, 3147–3159. [Google Scholar] [CrossRef] [Green Version]
- Bamberger, I.; Ruehr, N.K.; Schmitt, M.; Gast, A.; Wohlfahrt, G.; Arneth, A. Isoprene emission and photosynthesis during heatwaves and drought in black locust. Biogeosciences 2017, 14, 3649–3667. [Google Scholar] [CrossRef] [Green Version]
- Dani, K.G.S.; Benavides, A.M.S.; Michelozzi, M.; Peluso, G.; Torzillo, G.; Loreto, F. Relationship between isoprene emission and photosynthesis in diatoms, and its implications for global marine isoprene estimates. Mar. Chem. 2017, 189, 17–24. [Google Scholar] [CrossRef]
- Medeiros, D.B.; Martins, S.C.V.; Cavalcanti, J.H.F.; Daloso, D.M.; Martinoia, E.; Adriano, N.-N.; DaMatta, F.M.; Fernie, A.R.; Araújo, W.L. Enhanced Photosynthesis and Growth in atquac1 Knockout Mutants Are Due to Altered Organic Acid Accumulation and an Increase in Both Stomatal and Mesophyll Conductance. Plant Physiol. 2016, 170, 86–101. [Google Scholar] [CrossRef] [Green Version]
- Gago, J.; Daloso, D.; Figueroa, C.M.; Flexas, J.; Fernie, A.R.; Nikoloski, Z. Relationships of leaf net photosynthesis, stomatal conductance, and mesophyll conductance to primary plant metabolism: A multi-species meta-analysis approach. Plant Physiol. 2016, 171, 265–279. [Google Scholar] [CrossRef]
- Taylor, S.E.; Terry, N. Limiting Factors in Photosynthesis: V. Photochemical Energy Supply Colimits Photosynthesis at Low Values of Intercellular CO2 Concentration. Plant Physiol. 1984, 75, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Fila, G.; Badeck, F.-W.; Meyer, S.; Cerovic, Z.; Ghashghaie, J. Relationships between leaf conductance to CO2 diffusion and photosynthesis in micropropagated grapevine plants, before and after ex vitro acclimatization. J. Exp. Bot. 2006, 57, 2687–2695. [Google Scholar] [CrossRef] [Green Version]
- Tiiva, P.; Faubert, P.; Michelsen, A.; Holopainen, T.; Holopainen, J.K.; Rinnan, R. Climatic warming increases isoprene emission from a subarctic heath. New Phytol. 2008, 180, 853–863. [Google Scholar] [CrossRef]
- Nguyen, T.B.; Roach, P.J.; Laskin, J.; Physics, A.L.C. Effect of humidity on the composition and yield of isoprene photooxidation secondary organic aerosol. Atmos. Chem. Phys. Discuss. 2011, 11, 9217–9248. [Google Scholar] [CrossRef]
- Lamb, B.; Guenther, A.; Gay, D.; Westberg, H. A national inventory of biogenic hydrocarbon emissions. Atmos. Environ. 1987, 21, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Grote, R.; Samson, R.; Alonso, R.; Amorim, J.H.; Cariñanos, P.; Churkina, G.; Fares, S.; Thiec, D.L.; Niinemets, Ü.; Mikkelsen, T.N. Functional traits of urban trees: Air pollution mitigation potential. Front. Ecol. Environ. 2017, 14, 543–550. [Google Scholar] [CrossRef]
- Ghirardo, A.; Xie, J.; Zheng, X.; Wang, Y.; Grote, R.; Block, K.; Wildt, J.; Mentel, T.; Kiendler-Scharr, A.; Hallquist, M. Urban stress-induced biogenic VOC emissions and SOA-forming potentials in Beijing. Atmos. Chem. Phys. 2016, 16, 2901–2920. [Google Scholar] [CrossRef] [Green Version]
- Bureau, S.L.C.A.A. Optimizing the Layout of Street Trees and Tree Species to Create “One Street, One Scene” in Shencheng (in Chinese). 2020. Available online: http://lhsr.sh.gov.cn/zz202003/20200714/324d149b95e94333b497aaeda194c22c.html (accessed on 10 December 2020).
- Gallaun, H.; Zanchi, G.; Nabuurs, G.-J.; Hengeveld, G.; Schardt, M.; Verkerk, P.J. EU-wide maps of growing stock and above-ground biomass in forests based on remote sensing and field measurements. For. Ecol. Manag. 2010, 260, 252–261. [Google Scholar] [CrossRef]
- Cai, C.; Geng, F.; Tie, X.; Yu, Q.; An, J. Characteristics and source apportionment of VOCs measured in Shanghai, China. Atmos. Environ. 2010, 44, 5005–5014. [Google Scholar] [CrossRef]
- Guo, H.; Chen, K.; Wang, P.; Hu, J.; Zhang, H. Simulation of summer ozone and its sensitivity to emission changes in China. Atmos. Pollut. Res. 2019, 10, 1543–1552. [Google Scholar] [CrossRef]
- Dani, K.G.S.; Jamie, I.M.; Prentice, I.C.; Atwell, B.J. Evolution of isoprene emission capacity in plants. Trends Plant Sci. 2014, 19, 439–446. [Google Scholar] [CrossRef]
- Fares, S.; Gentner, D.R.; Park, J.H.; Ormeno, E.; Karlik, J.; Goldstein, A.H. Biogenic emissions from Citrus species in California. Atmos. Environ. 2011, 45, 4557–4568. [Google Scholar] [CrossRef]
- Pokorska, O.; Dewulf, J.; Amelynck, C.; Schoon, N.; Joo, E.; Simpraga, M.; Bloemen, J.; Steppe, K.; Langenhove, H.V. Emissions of biogenic volatile organic compounds from Fraxinus excelsior and Quercus robur under ambient conditions in Flanders (Belgium). Int. J. Environ. Anal. Chem. 2012, 92, 1729–1741. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Photosynthetic Rate (μmol CO2·m−2·s−1) | Intercellular CO2 Concentration (μmol CO2·mol−1) | Stomatal Conductance (mol H2O·m−2·s−1) | Transpiration Rate (mmol H2O·m−2·s−1) | ||||
|---|---|---|---|---|---|---|---|---|
| Spr. | Sum. | Spr. | Sum. | Spr. | Sum. | Spr. | Sum. | |
| Salix babylonica | 9.6 ± 1.8 | 11.8 ± 2.2 | 273.3 ± 11.9 | 258.0 ± 13.5 | 0.14 ± 0.04 | 0.18 ± 0.05 | 3.98 ± 0.99 | 3.21 ± 0.73 |
| Ginkgo biloba | 9.2 ± 0.9 | 9.0 ± 1.5 | 260.7 ± 16.4 | 262.2 ± 11.8 | 0.11 ± 0.01 | 0.13 ± 0.01 | 2.49 ± 0.20 | 2.76 ± 0.19 |
| Cinnamomum camphora | 8.4 ± 1.1 | 10.6 ± 1.5 | 367.2 ± 13.5 | 216.6 ± 53.3 | 0.10 ± 0.01 | 0.09 ± 0.04 | 1.73 ± 0.08 | 1.66 ± 0.50 |
| Magnolia grandiflora | 7.6 ± 0.8 | 9.9 ± 1.9 | 296.8 ± 16.8 | 264.0 ± 6.5 | 0.09 ± 0.01 | 0.16 ± 0.01 | 1.76 ± 0.18 | 2.77 ± 0.30 |
| Elaeocarpus decipiens | 6.6 ± 0.8 | 12.3 ± 0.7 | 281.1 ± 14.4 | 226.9 ± 18.2 | 0.06 ± 0.02 | 0.08 ± 0.01 | 1.84 ± 0.33 | 1.30 ± 0.23 |
| Fatsia japonica | 6.4 ± 0.4 | 8.1 ± 0.7 | 239.6 ± 7.0 | 273.4 ± 13.2 | 0.09 ± 0.02 | 0.14 ± 0.02 | 1.98 ± 0.33 | 2.92 ± 0.36 |
| Viburnum odoratissimum | 6.3 ± 0.8 | 11.2 ± 0.8 | 272.5 ± 12.1 | 228.2 ± 7.0 | 0.09 ± 0.01 | 0.14 ± 0.02 | 2.14 ± 0.23 | 2.78 ± 0.26 |
| Osmanthus fragrans | 5.6 ± 0.6 | 14.0 ± 1.4 | 232.2 ± 32.5 | 251.2 ± 17.2 | 0.06 ± 0.01 | 0.12 ± 0.03 | 1.76 ± 0.26 | 2.05 ± 0.40 |
| Photinia× fraseri | 5.4 ± 0.7 | 10.3 ± 1.1 | 281.7 ± 15.5 | 218.9 ± 25.7 | 0.07 ± 0.01 | 0.07 ± 0.01 | 1.62 ± 0.26 | 1.82 ± 0.04 |
| Ligustrum lucidum | 4.9 ± 0.4 | 8.1 ± 0.6 | 272.2 ± 11.4 | 259.3 ± 16.8 | 0.06 ± 0.01 | 0.11 ± 0.03 | 1.80 ± 0.18 | 2.61 ± 0.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, J.; Xiong, F.; Sun, N.; Li, Y.; Liu, C.; Yin, S. Photosynthesis and Related Physiological Parameters Differences Affected the Isoprene Emission Rate among 10 Typical Tree Species in Subtropical Metropolises. Int. J. Environ. Res. Public Health 2021, 18, 954. https://doi.org/10.3390/ijerph18030954
Lyu J, Xiong F, Sun N, Li Y, Liu C, Yin S. Photosynthesis and Related Physiological Parameters Differences Affected the Isoprene Emission Rate among 10 Typical Tree Species in Subtropical Metropolises. International Journal of Environmental Research and Public Health. 2021; 18(3):954. https://doi.org/10.3390/ijerph18030954
Chicago/Turabian StyleLyu, Junyao, Feng Xiong, Ningxiao Sun, Yiheng Li, Chunjiang Liu, and Shan Yin. 2021. "Photosynthesis and Related Physiological Parameters Differences Affected the Isoprene Emission Rate among 10 Typical Tree Species in Subtropical Metropolises" International Journal of Environmental Research and Public Health 18, no. 3: 954. https://doi.org/10.3390/ijerph18030954
APA StyleLyu, J., Xiong, F., Sun, N., Li, Y., Liu, C., & Yin, S. (2021). Photosynthesis and Related Physiological Parameters Differences Affected the Isoprene Emission Rate among 10 Typical Tree Species in Subtropical Metropolises. International Journal of Environmental Research and Public Health, 18(3), 954. https://doi.org/10.3390/ijerph18030954