Genetic Scores of eNOS, ACE and VEGFA Genes Are Predictive of Endothelial Dysfunction Associated Osteoporosis in Postmenopausal Women

 ,
, 
Abstract
:1. Introduction
2. Subjects and Methods
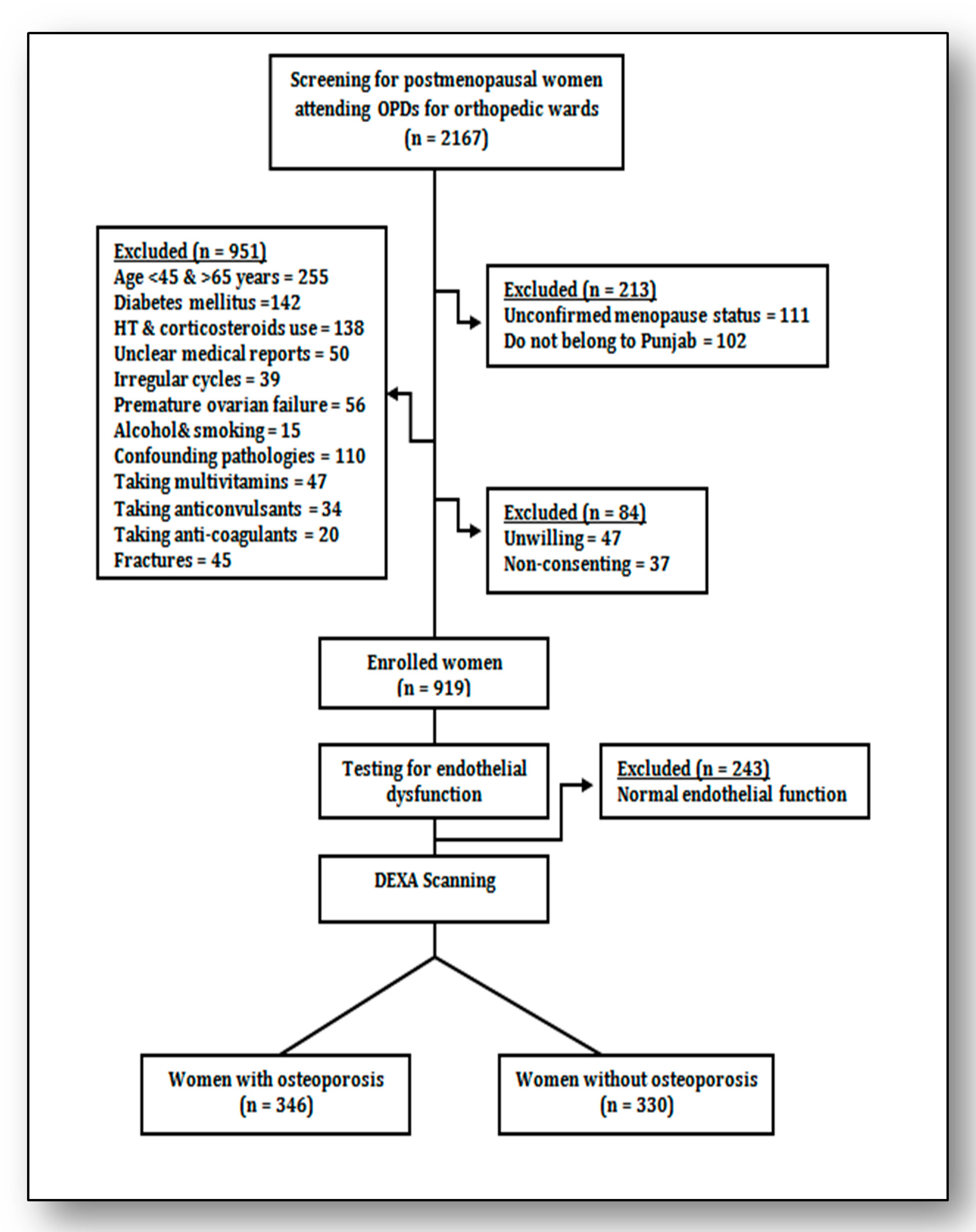
2.1. Subjects
2.2. Description of Variables
2.3. Examination of Endothelial Function
2.4. BMD Evaluation
2.5. Selection of the SNPs and Genotyping
2.6. Population Stratification Analysis and Statistical Power of Genetic Association
2.7. Statistical Analysis
2.8. Receiver Operating Characteristic Curve Analysis
3. Result
3.1. Analysis of Variables at Baseline and Genetic Correlates
3.2. Identification of Independent Risk Variables
3.3. Genotype Specific Implications of Genes through Different Genetic Models
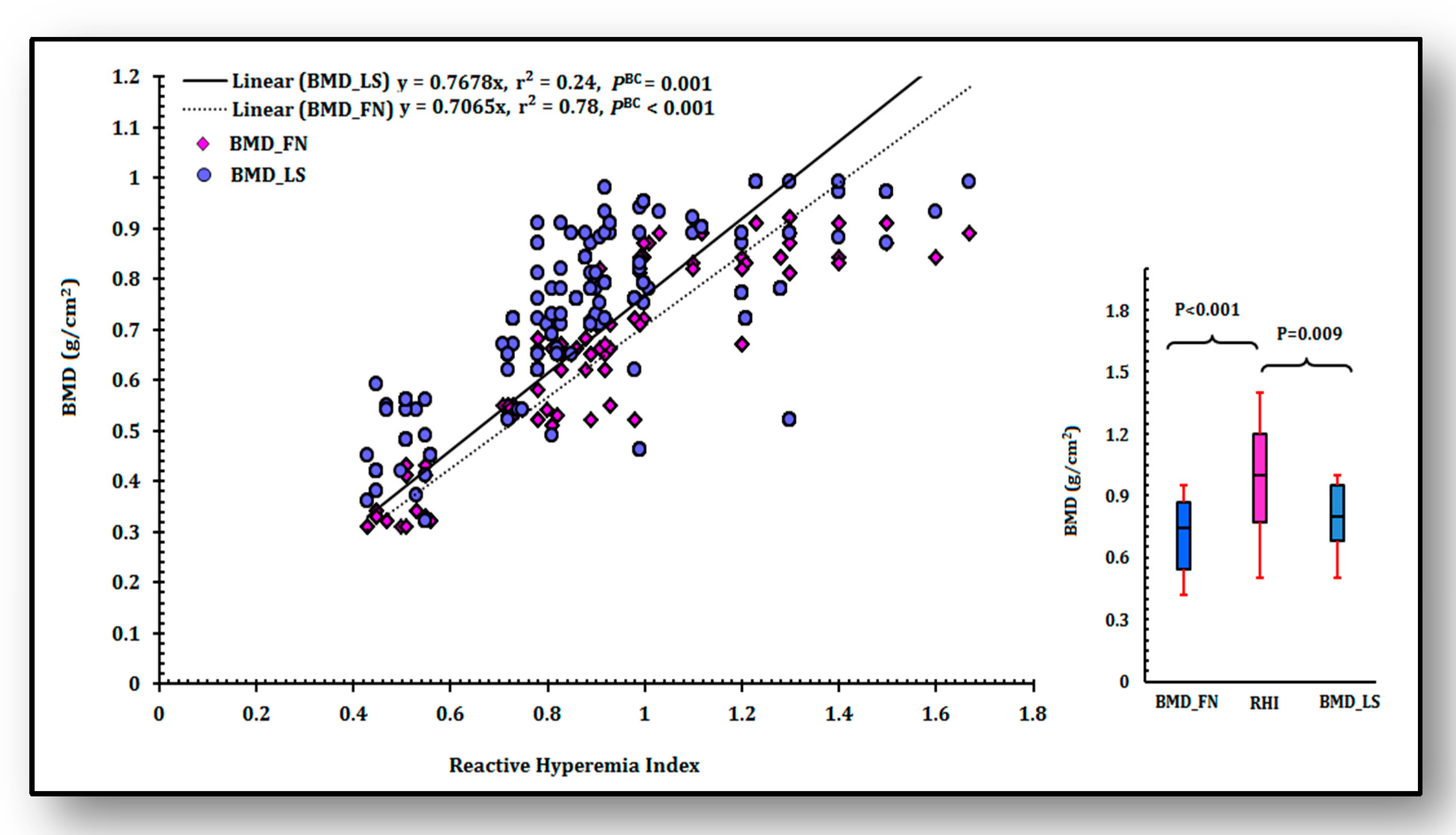
3.4. Analysis of Linear Relationship of RHI with BMD
3.5. SNP-SNP Cross Talks, Risky Traits and Their Modes of Association
3.6. Haplotype Analysis, Their Contribution and Best Mode of Impact for Osteoporosis Risk
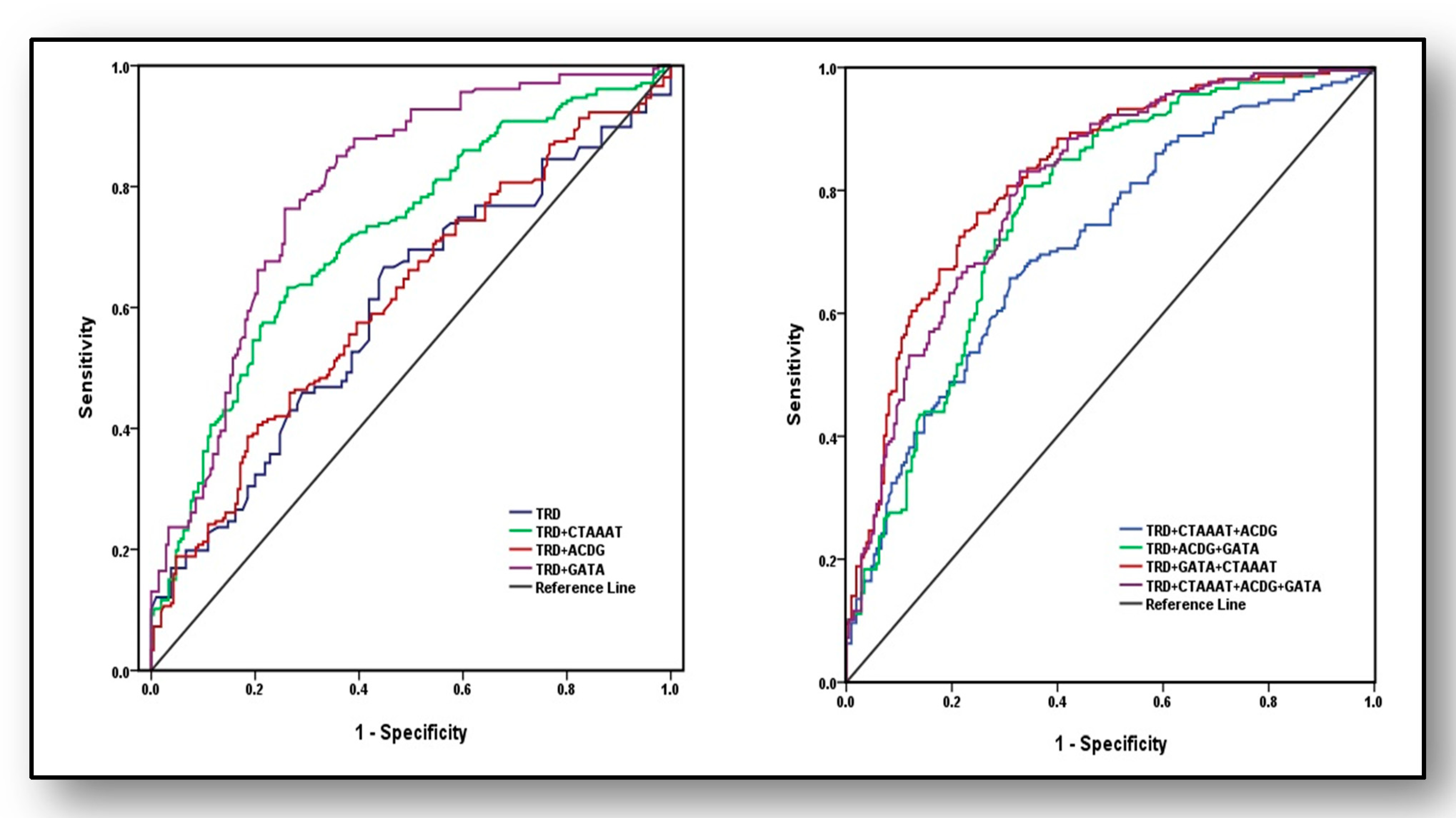
3.7. Predictive Ability of Haplotypes and Traditional Risk Factors for the Diagnosis of Osteoporosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prasad, M.; Reriani, M.; Khosla, S.; Gössl, M.; Lennon, R.; Gulati, R.; Prasad, A.; Lerman, L.O.; Lerman, A. Coronary microvascular endothelial dysfunction is an independent predictor of development of osteoporosis in postmenopausal women. Vasc. Health Risk Manag. 2014, 10, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, V.M.; Duckles, S.P. Vascular actions of estrogens: Functional implications. Pharmacol. Rev. 2008, 60, 210–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnal, J.F.; Scarabin, P.Y.; Trémollières, F.; Laurell, H.; Gourdy, P. Estrogens in vascular biology and disease: Where do we stand today? Curr. Opin. Lipidol. 2007, 18, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Van’T Hof, R.J.; Ralston, S.H. Nitric oxide and bone. Immunology 2001, 103, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, E.C.; Adams, R.H. Biology of Bone: The Vasculature of the Skeletal System. Cold Spring Harb. Perspect. Med. 2018, 8, a031559. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, M.E.; Karas, R.H. The protective effects of estrogen on the cardiovascular system. N. Engl. J. Med. 1999, 340, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.; Maas, A.; Prasad, M.; Schierbeck, L.; Lerman, A. Endothelial Vascular Function as a Surrogate of Vascular Risk and Aging in Women. Mayo Clin. Proc. 2020, 95, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yuhanna, I.S.; Galcheva-Gargova, Z.; Karas, R.H.; Mendelsohn, M.E.; Shaul, P.W. Estrogen receptor alpha mediates the nongenomic activation of endothelial nitric oxide synthase by estrogen. J. Clin. Investig. 1999, 103, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, R.E.; Silva, M.J. Skeletal Blood Flow in Bone Repair and Maintenance. Bone Res. 2013, 1, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Marenzana, M.; Arnett, T.R. The Key Role of the Blood Supply to Bone. Bone Res. 2013, 1, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Rucci, N. Molecular biology of bone remodelling. Clin. Cases Miner. Bone Metab. 2008, 5, 49–56. [Google Scholar] [PubMed]
- Blum, A.; Zarqh, O.; Peleg, A.; Sirchan, R.; Blum, N.; Salameh, Y.; Ganaem, M. Vascular inflammation and endothelial dysfunction in fracture healing. Am. J. Orthop. Belle Mead NJ 2012, 41, 87–91. [Google Scholar] [PubMed]
- Zhao, Y.; Xie, L. Unique bone marrow blood vessels couple angiogenesis and osteogenesis in bone homeostasis and diseases. Ann. N. Y. Acad. Sci. 2020, 1474, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Massett, M.P. Genetic Regulation of Endothelial Vasomotor Function. Front. Physiol. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.C.; Hingorani, A.D. Genetic regulation of endothelial function. Heart 2005, 9, 1275–1277. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Singh, M.; Valecha, S.; Khinda, R.; Kumar, N.; Mastana, S.; Singh, S.; Juneja, P.K.; Kaur, T. Investigation of eNOS gene polymorphism exposes a genetic association between endothelial dysfunction and osteoporosis in postmenopausal women. Menopause 2020, 27, 714–721. [Google Scholar] [CrossRef]
- Elakkad, A.M.; Abou-Aisha., K.; Hassanein, S.I.; Gad, M.Z. T-786C variation in the promoter sequence of human eNOS gene markedly influences its expression level. Drug Discov. Ther. 2017, 1, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Brindisi, M.; Fiorillo, M.; Frattaruolo, L.; Sotgia, F.; Lisanti, M.P.; Cappello, A.R. Cholesterol and Mevalonate: Two Metabolites Involved in Breast Cancer Progression and Drug Resistance through the ERRα Pathway. Cells 2020, 9, 1819. [Google Scholar] [CrossRef]
- Ghanbari, F.; Mader, S.; Philip, A. Cholesterol as an Endogenous Ligand of ERRα Promotes ERRα-Mediated Cellular Proliferation and Metabolic Target Gene Expression in Breast Cancer Cells. Cells 2020, 9, 1765. [Google Scholar] [CrossRef]
- Sumi, D.; Ignarro, L.J. Estrogen-related receptor alpha 1 up-regulates endothelial nitric oxide synthase expression. Proc. Natl. Acad. Sci. USA 2003, 100, 14451–14456. [Google Scholar] [CrossRef] [Green Version]
- Butler, R.; Morris, A.D.; Burchell, B.; Struthers, A.D. DD angiotensin-converting enzyme gene polymorphism is associated with endothelial dysfunction in normal humans. Hypertension 1999, 33, 1164–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkin, C.E.; Gerstenfeld, L.C. VEGF and bone cell signalling: An essential vessel for communication? Cell Biochem. Funct. 2013, 31, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bussolati, B.; Dunk, C.; Grohman, M.; Kontos, C.D.; Mason, J.; Ahmed, A. Vascular endothelial growth factor receptor-1 modulates vascular endothelial growth factor-mediated angiogenesis via nitric oxide. Am. J. Pathol. 2001, 159, 993–1008. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Berendsen, A.D.; Jia, S.; Lotinun, S.; Baron, R.; Ferrara, N.; Olsen, B.R. Intracellular VEGF regulates the balance between osteoblast and adipocyte differentiation. J. Clin. Investig. 2012, 122, 3101–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazizadeh, H.; Rezaei, M.; Avan, A.; Fazilati, M.; Pasdar, A.; Tavallaie, S.; Kazemi, E.; Seyedi, S.M.R.; Ferns, G.A.; Azimi-Nezhad, M.; et al. Association between serum cell adhesion molecules with hs-CRP, uric acid and VEGF genetic polymorphisms in subjects with metabolic syndrome. Mol. Biol. Rep. 2020, 47, 867–875. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Menashe, I.; Rosenberg, P.S.; Chen, B.E. PGA: Power calculator for case-control genetic association analyses. BMC Genet. 2008, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Runesha, H.B.; Dvorkin, D.; Garbe, J.R.; Da, Y. Parallel and serial computing tools for testing single-locus and epistatic SNP effects of quantitative traits in genome-wide association studies. BMC Bioinform. 2008, 9, 315. [Google Scholar] [CrossRef] [Green Version]
- Stram, D.O. Multi-SNP Haplotype Analysis Methods for Association Analysis. Methods Mol. Biol. 2017, 1666, 485–504. [Google Scholar] [CrossRef]
- Liu, S.Z.; Yan, H.; Hou, W.K.; Xu, P.; Tian, J.; Tian, L.F.; Zhu, B.F.; Ma, J.; Lu, S.M. Relationships between endothelial nitric oxide synthase gene polymorphisms and osteoporosis in postmenopausal women. J. Zhejiang Univ. Sci. B. 2009, 10, 609–618. [Google Scholar] [CrossRef]
- Cakmak, B.; Inanir, A.; Karakus, N.; Ates, O.; Yigit, S. Association between the ACE gene I/D polymorphism and osteoporosis in a Turkish population. Z. Rheumatol. 2015, 74, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.; Paramanathan, S.; Mac Donald, D.; Wierzbicki, A.S.; Hampson, G. Factors regulating circulating vascular endothelial growth factor (VEGF): Association with bone mineral density (BMD) in post-menopausal osteoporosis. Cytokine 2009, 46, 376–381. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, E.; Scheid, J.L.; West, S.L.; De Souza, M.J. Impaired vascular function in exercising anovulatory premenopausal women is associated with low bone mineral density. Scand. J. Med. Sci. Sports 2019, 29, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Naber, C.K.; Baumgart, D.; Altmann, C.; Siffert, W.; Erbel, R.; Heusch, G. eNOS 894T allele and coronary blood flow at rest and during adenosine-induced hyperemia. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1908–H1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Mori, S.; Ishizaki, T.; Takahashi, A.; Matsuda, K.; Koretsune, Y.; Minami, S.; Higashiyama, M.; Imai, S.; Yoshimori, K.; et al. Genetic risk score based on the prevalence of vertebral fracture in Japanese women with osteoporosis. Bone Rep. 2016, 5, 168–172. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variables | Endothelial Dysfunction | p Value | |
|---|---|---|---|
| With Osteoporosis | Without Osteoporosis | ||
| Number of subjects | 346 | 330 | ------- |
| Age (years) | 60.21 ± 12.4 | 61.19 ± 8.92 | 0.934 |
| Years since menopause (years) | 8.86 ± 5.03 | 8.93 ± 3.67 | 0.106 |
| Body mass index (kg/m2) | 30.21 ± 3.11 | 26.2 ± 3.72 | <0.001 |
| Systolic blood pressure (mmHg) | 129.56 ± 13.72 | 125.18 ± 12.77 | <0.001 |
| Diastolic blood pressure (mmHg) | 98.10 ± 9.67 | 96.11 ± 10.13 | 0.087 |
| Low density lipoprotein (mg/dL) | 199.32 ± 20.18 | 194.08 ± 19.50 | 0.441 |
| Triglyceride (mg/dL) a | 190(189, 225) | 145 (102, 201) | <0.001 |
| High density lipoprotein (mg/dL) | 44.23 ± 4.16 | 45.79 ± 3.15 | 0.189 |
| Total cholesterol (mg/dL) | 227.81± 20.25 | 221.11 ± 20.78 | 0.061 |
| BMD_FN(g/cm2) a | 0.73 ± 0.14 | 0.89 ± 0.11 | <0.001 |
| BMD_LS(g/cm2) a | 0.84 ± 0.11 | 0.94 ± 0.14 | <0.001 |
| Reactive Hyperemia Index b | 1.23 (1.09, 1.34) | 1.44 (1.34, 1.55) | <0.001 |
| eNOS gene/SNPS | |||
| rs2070744 (MAF ± SE) c | 0.26 ± 0.024 | 0.17 ± 0.015 | 0.002 |
| rs1799983 (MAF ± SE) c | 0.21 ± 0.022 | 0.10 ± 0.016 | <0.001 |
| rs1800780 (MAF ± SE) c | 0.43 ± 0.047 | 0.42 ± 0.027 | 0.855 |
| rs3918181 (MAF ± SE) c | 0.33 ± 0.025 | 0.29 ± 0.025 | 0.259 |
| rs891512 (MAF ± SE) c | 0.24 ± 0.023 | 0.16 ± 0.020 | 0.009 |
| rs1808593 (MAF ± SE) c | 0.21 ± 0.022 | 0.17 ± 0.021 | 0.189 |
| ACE gene/SNPS | |||
| rs4459609(MAF ± SE) c | 0.36 ± 0.026 | 0.35 ± 0.026 | 0.786 |
| rs1800764(MAF ± SE) c | 0.33 ± 0.025 | 0.26 ± 0.024 | 0.044 |
| rs1799752(MAF ± SE) c | 0.56 ± 0.027 | 0.46 ± 0.020 | 0.003 |
| rs4343(MAF ± SE) c | 0.31 ± 0.025 | 0.24 ± 0.023 | 0.040 |
| VEGFA gene/SNPS | |||
| rs2010963(MAF ± SE) c | 0.20 ± 0.022 | 0.27 ± 0.024 | 0.032 |
| rs833061(MAF ± SE) c | 0.42 ± 0.027 | 0.41 ± 0.027 | 0.793 |
| rs699947(MAF ± SE) c | 0.46 ± 0.027 | 0.38 ± 0.027 | 0.036 |
| rs1570360(MAF ± SE) c | 0.31 ± 0.025 | 0.22 ± 0.023 | 0.008 |
| Variables | Univariate Analysis | Multivariate Analysis | ||||
|---|---|---|---|---|---|---|
| OR | 95%CI | p Values | OR | 95%CI | p Values | |
| Body mass index (kg.m−2) | 1.90 | 2.10–3.12 | <0.001 | 1.45 | 1.11–2.76 | 0.007 |
| Years since menopause(years) | 1.85 | 2.00–3.89 | 0.008 | 1.72 | 1.78–2.52 | 0.020 |
| Diastolic blood pressure (mmHg) | 1.61 | 0.85–2.95 | 0.281 | ----- | --------- | -------- |
| Systolic blood pressure (mmHg) | 1.90 | 1.33–3.02 | 0.009 | 1.42 | 1.12–2.89 | 0.012 |
| Total cholesterol (mg/dL) | 1.67 | 0.91–2.95 | 0.288 | ----- | --------- | ------- |
| Low density lipoprotein (mg/dL) | 2.45 | 0.87–2.90 | 0.211 | ----- | --------- | ------- |
| High density lipoprotein (mg/dL) | 1.37 | 0.61–2.35 | 0.155 | ----- | --------- | ------ |
| Triglyceride (mg/dL) | 2.31 | 1.59–3.10 | <0.001 | 1.75 | 1.24–3.09 | 0.009 |
| BMD_FN(g/cm2) | 1.77 | 1.28–3.41 | <0.001 | 1.20 | 1.10–2.55 | 0.005 |
| BMD_LS (g/cm2) | 1.51 | 1.19–3.23 | 0.006 | 1.27 | 1.12–3.00 | 0.013 |
| SNPs/Genetic Model | Input Parameters | Unadjusted OR (95% CI) | p Value | Adjusted OR (95% CI) | p Value |
|---|---|---|---|---|---|
| rs2070744 | TT | Referent | ----- | Referent | ----- |
| Codominant | TT vs. TC | 1.77 (1.26–2.47) | 0.001 | 1.55 (1.18–2.19) | 0.007 |
| Codominant | TT vs. CC | 2.15 (1.11–4.15) | 0.032 | 1.73 (1.09–3.82) | 0.043 |
| Dominant | TT vs. TC + CC | 1.94 (1.16–3.24) | 0.016 | 1.65 (1.10–3.19) | 0.037 |
| Recessive | TT + TC vs. CC | 1.84 (0.67–5.06) | 0.343 | 1.46 (0.59–4.76) | 0.287 |
| Multiplicative | 2TT + TC vs.TC + 2CC | 1.67 (1.28–2.17) | <0.001 | 1.33 (1.14–2.00) | 0.005 |
| rs1799983 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GT | 2.86 (1.97–4.15) | <0.001 | 2.53 (1.81–3.91) | 0.006 |
| Codominant | GG vs. TT | 3.18 (1.21–8.33) | 0.025 | 3.00 (1.14–8.19) | 0.042 |
| Dominant | GG vs. GT + TT | 3.19 (1.75–5.82) | <0.001 | 2.84 (1.66–5.29) | 0.009 |
| Recessive | GG + GT vs. TT | 2.72 (0.54–13.75) | 0.368 | 2.49 (0.37–10.51) | 0.222 |
| Multiplicative | 2GG + GT vs. GT + 2TT | 2.51 (1.83–3.44) | <0.001 | 2.13 (1.27–2.92) | 0.002 |
| rs1800780 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GA | 0.89 (0.64–1.26) | 0.580 | 0.72 (0.59–1.13) | 0.491 |
| Codominant | GG vs. AA | 1.13 (0.72–1.76) | 0.680 | 1.10 (0.70–1.72) | 0.672 |
| Dominant | GG vs. GA + AA | 0.91 (0.54–1.53) | 0.829 | 0.81 (0.50–1.48) | 0.800 |
| Recessive | GG + GA vs. AA | 1.48 (0.79–2.76) | 0.280 | 1.33 (0.71–2.65) | 0.271 |
| Multiplicative | 2GG + GA vs. GA + 2AA | 1.03 (0.83–1.28) | 0.890 | 0.88 (0.71–1.11) | 0.786 |
| rs3918181 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GA | 1.37 (0.99–1.88) | 0.067 | 1.20 (0.82–1.71) | 0.079 |
| Codominant | GG vs. AA | 1.17 (0.70–1.96) | 0.646 | 1.09 (0.56–1.78) | 0.538 |
| Dominant | GG vs. GA + AA | 1.16 (0.71–1.89) | 0.653 | 1.00 (0.51–1.70) | 0.540 |
| Recessive | GG + GA vs. AA | 1.47 (0.68–3.18) | 0.423 | 1.38 (0.62–3.01) | 0.411 |
| Multiplicative | 2GG + GA vs. GA + 2AA | 1.18 (0.94–1.49) | 0.177 | 0.92 (0.69–1.21) | 0.165 |
| rs891512 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GA | 1.68 (1.20–2.34) | 0.003 | 1.55 (1.15–2.28) | 0.029 |
| Codominant | GG vs. AA | 2.43 (1.16–5.11) | 0.026 | 2.20 (1.10–4.92) | 0.040 |
| Dominant | GG vs. GA + AA | 1.79 (1.06–3.03) | 0.041 | 1.67(1.02–2.77) | 0.050 |
| Recessive | GG + GA vs. AA | 1.51 (0.53–4.28) | 0.605 | 1.45 (0.45–4.12) | 0.519 |
| Multiplicative | 2GG + GA vs. GA + 2AA | 1.66 (1.27–2.18) | <0.001 | 1.51 (1.13–2.09) | 0.034 |
| rs1808593 | TT | Referent | ----- | Referent | ----- |
| Codominant | TT vs. TG | 1.26 (0.85–1.68) | 0.330 | 1.10 (0.71–1.47) | 0.290 |
| Codominant | TT vs. GG | 1.98 (0.93–4.20) | 0.105 | 1.82 (0.82–4.01) | 0.097 |
| Dominant | TT vs. TG + GG | 1.57 (0.91–2.69) | 0.134 | 1.43 (0.81–2.37) | 0.111 |
| Recessive | TT + TG vs.GG | 1.63 (0.53–5.01) | 0.558 | 1.52 (0.41–4.74) | 0.419 |
| Multiplicative | 2TT + TG vs. TG + 2GG | 1.31 (1.00–1.72) | 0.061 | 1.19 (0.87–1.49) | 0.091 |
| SNPs/Genetic Model | Input Parameters | Unadjusted OR (95%CI) | p Value | Adjusted OR (95%CI) | p Value |
|---|---|---|---|---|---|
| rs4459609 | AA | Referent | ----- | Referent | ----- |
| Codominant | AA vs. AC | 1.09 (0.79–1.51) | 0.652 | 1.00 (0.70–1.45) | 0.579 |
| Codominant | AA vs. CC | 1.05 (0.64–1.73) | 0.944 | 0.93 (0.55–161) | 0.817 |
| Dominant | AA vs. AC + CC | 1.08 (0.80–1.40) | 0.660 | 1.00 (0.71–1.26) | 0.531 |
| Recessive | AA + AC vs. CC | 1.00 (0.63–1.60) | 0.920 | 0.87 (0.58–1.43) | 0.779 |
| Multiplicative | 2AA + AC vs.AC + 2CC | 1.04 (0.89–1.30) | 0.749 | 0.93 (0.76–1.23) | 0.654 |
| rs1800764 | TT | Referent | ----- | Referent | ----- |
| Codominant | TT vs. TC | 1.27 (0.92–1.75) | 0.167 | 1.18 (0.85–1.62) | 0.141 |
| Codominant | TT vs. CC | 1.89 (1.11–3.21) | 0.024 | 1.71 (1.08–3.17) | 0.038 |
| Dominant | TT vs. TC + CC | 1.38 (1.02–1.87) | 0.044 | 1.21 (1.00–1.73) | 0.053 |
| Recessive | TT + TC vs. CC | 1.70 (1.02–2.84) | 0.053 | 1.43 (0.92–2.61) | 0.092 |
| Multiplicative | 2TT + TC vs. TC + 2CC | 1.36 (1.08–1.72) | 0.012 | 1.22 (1.01–1.62) | 0.043 |
| rs1799752 | II | Referent | ----- | Referent | ----- |
| Codominant | II vs. ID | 1.57 (1.07–2.31) | 0.027 | 1.37(1.05–2.27) | 0.044 |
| Codominant | II vs. DD | 2.28 (1.40–3.56) | <0.001 | 2.19 (1.33–3.41) | 0.007 |
| Dominant | II vs. ID + DD | 1.77 (1.23–2.55) | 0.003 | 1.68 (1.20–2.38) | 0.013 |
| Recessive | II + ID vs. DD | 1.66 (1.17–2.37) | 0.006 | 1.42 (1.03–2.13) | 0.028 |
| Multiplicative | 2II + ID vs. ID + 2DD | 1.48 (1.19–1.83) | <0.001 | 1.22 (1.11–1.69) | 0.008 |
| rs4343 | AA | Referent | ----- | Referent | ----- |
| Codominant | AA vs. AG | 1.65 (1.19–2.28) | 0.003 | 1.51 (1.14–2.17) | 0.009 |
| Codominant | AA vs. GG | 1.63 (0.94–2.85) | 0.110 | 1.49 (0.81–2.13) | 0.098 |
| Dominant | AA vs. AG + GG | 1.65 (1.23–2.23) | 0.002 | 1.43 (1.17–2.00) | 0.028 |
| Recessive | AA + AG vs. GG | 1.33 (0.77–2.28) | 0.368 | 1.24 (0.65–1.99) | 0.289 |
| Multiplicative | 2AA + AG vs. AG + 2GG | 1.44 (1.13–1.84) | 0.003 | 1.29 (1.09–1.72) | 0.015 |
| SNPs/Genetic Model | Input Parameters | Unadjusted OR (95%CI) | p Value | Adjusted OR (95%CI) | p Value |
|---|---|---|---|---|---|
| rs2010963 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GC | 0.63 (0.45–0.87) | 0.007 | 0.60 (0.42–0.83) | 0.027 |
| Codominant | GG vs. CC | 0.58 (0.32–1.07) | 0.112 | 0.52 (0.29–1.10) | 0.101 |
| Dominant | GG vs. GC +CC | 0.62 (0.46–0.85) | 0.003 | 0.57 (0.41–0.79) | 0.036 |
| Recessive | GG +GC vs. CC | 0.69 (0.38–1.25) | 0.282 | 0.63 (0.29–1.10) | 0.178 |
| Multiplicative | 2GG + GC vs. GC + 2CC | 0.68 (0.53–0.88) | 0.004 | 0.65 (0.50–0.82) | 0.010 |
| rs833061 | CC | Referent | ----- | Referent | ----- |
| Codominant | CC vs. CA | 0.97 (0.69–1.35) | 0.905 | 0.84 (0.55–1.22) | 0.811 |
| Codominant | CC vs. AA | 1.12 (0.73–1.73) | 0.674 | 1.05 (0.63–1.65) | 0.555 |
| Dominant | CC vs. CA + CC | 1.01 (0.74–1.38) | 0.978 | 0.89 (0.63–1.19) | 0.818 |
| Recessive | CC +CA vs. AA | 1.15 (0.78–1.69) | 0.550 | 1.12 (0.72–1.63) | 0.532 |
| Multiplicative | 2CC + CA vs. CA + 2AA | 1.05 (0.84–1.30) | 0.713 | 0.92 (0.76–1.18) | 0.582 |
| rs699947 | TT | Referent | ----- | Referent | ----- |
| Codominant | TT vs. TC | 1.29 (0.92–1.82) | 0.161 | 1.18 (0.80–1.65) | 0.145 |
| Codominant | TT vs. CC | 1.86 (1.20–2.88) | 0.008 | 1.72 (1.11–2.72) | 0.027 |
| Dominant | TT vs. TC + CC | 1.43 (1.04–1.97) | 0.032 | 1.30 (1.00–1.82) | 0.048 |
| Recessive | TT +TC vs. CC | 1.60 (1.08–2.38) | 0.024 | 1.51 (1.01–2.24) | 0.039 |
| Multiplicative | 2TT + TC vs. TC + 2CC | 1.37 (1.10–1.70) | 0.005 | 1.22 (1.05–1.61) | 0.027 |
| rs1570360 | GG | Referent | ----- | Referent | ----- |
| Codominant | GG vs. GA | 1.64 (1.18–2.27) | 0.004 | 1.52 (1.11–2.19) | 0.011 |
| Codominant | GG vs. AA | 2.26 (1.28–3.99) | 0.006 | 2.13 (1.17–3.29) | 0.026 |
| Dominant | GG vs. GA + AA | 1.74 (1.28–2.37) | <0.001 | 1.69 (1.20–2.31) | 0.017 |
| Recessive | GG +GA vs. AA | 1.87 (1.07–3.25) | 0.035 | 1.74 (0.92–2.88) | 0.062 |
| Multiplicative | 2GG + GA vs. GA + 2AA | 1.62 (1.27–2.07) | <0.001 | 1.53 (1.18–1.95) | 0.025 |
| SNP | SNP | Trait | Test | PO | PNO |  |
| rs2070744 | rs4343 | RHI | I | 0.0013 | 0.432 | |
| rs2070744 | rs1799983 | LDL | AA | 0.0052 | 1.348 | |
| rs2070744 | rs1800764 | TG | DA | 0.0011 | 0.041 | |
| rs2070744 | rs891512 | SBP | AD | 0.0035 | 0.122 | |
| rs4343 | rs2010963 | RHI | AA | 0.0090 | 0.178 | |
| rs1800764 | rs1799752 | TC | DD | 0.0290 | 2.152 | |
| rs1799983 | rs891512 | RHI | I | 0.0320 | 1.813 | |
| rs1799983 | rs699947 | BMI | DD | 0.0014 | 0.041 | |
| rs699947 | rs1799752 | RHI | I | 0.0035 | 0.082 | |
| rs891512 | rs1799752 | LDL | DA | 0.0221 | 0.078 |
| Haplotype | Endothelial Dysfunction | PCor. | Unadjusted OR (95%CI) | p Value | Adjusted OR(95% CI) a | p Value | |
|---|---|---|---|---|---|---|---|
| With | Without | ||||||
| Osteoporosis | Osteoporosis | ||||||
| eNOS gene | |||||||
| TGAAGT | 0.21 (73) | 0.20 (66) | 0.97 | Referent | -------- | Referent | ------ |
| TTAGGG | 0.11 (38) | 0.15 (49) | 0.21 | 0.70 (0.41–1.20) | 0.25 | 0.61 (0.32–1.13) | 0.19 |
| TTGGGG | 0.09 (31) | 0.12 (40) | 0.31 | 0.70 (0.39–1.25) | 0.28 | 0.52 (0.32–1.19) | 0.22 |
| CGAAGG | 0.10 (35) | 0.12 (40) | 0.71 | 0.79 (0.45–1.39) | 0.50 | 0.72 (0.33–1.21) | 0.39 |
| CTAAAT | 0.18(62) | 0.06 (20) | 1 × 10−8 | 2.80 (1.53–5.13) | 0.001 | 2.43 (1.22–4.71) | 0.007 |
| CGGAGG | 0.07 (24) | 0.08 (26) | 0.93 | 0.83 (0.44–1.59) | 0.72 | 0.65 (0.39–1.42) | 0.50 |
| CTGGAT | 0.06 (21) | 0.08 (26) | 0.63 | 0.73 (0.38–1.42) | 0.45 | 0.57 (0.34–1.21) | 0.39 |
| TTAAAT | 0.06 (21) | 0.05 (16) | 0.80 | 1.19 (0.57–2.46) | 0.78 | 1.02 (0.48–2.35) | 0.71 |
| ACE gene | |||||||
| ATIA | 0.31 (107) | 0.35 (115) | 0.50 | Referent | -------- | Referent | ------ |
| ACDG | 0.27 (93) | 0.10 (33) | 1 × 10−9 | 3.03 (1.86–4.88) | <0.001 | 2.50 (1.28–3.96) | 0.002 |
| CTIG | 0.12 (41) | 0.14 (46) | 0.72 | 0.96 (0.58–1.57) | 0.97 | 0.80 (0.42–1.21) | 0.42 |
| CCIG | 0.11 (38) | 0.12 (40) | 0.93 | 1.02 (0.61–1.71) | 0.96 | 0.98 (0.33–1.38) | 0.71 |
| ACDA | 0.09 (31) | 0.11 (36) | 0.70 | 0.93 (0.54–1.60) | 0.89 | 0.68 (0.47–1.16) | 0.67 |
| VEGFA gene | |||||||
| GCTG | 0.28 (86) | 0.29 (96) | 0.38 | Referent | -------- | Referent | ------ |
| GATA | 0.19 (66) | 0.08 (26) | 1 × 10−7 | 2.83 (1.65–4.86) | <0.001 | 2.10 (1.31–3.29) | 0.009 |
| GACG | 0.14 (48) | 0.16 (53) | 0.73 | 1.01 (0.62–1.65) | 0.94 | 0.92 (0.42–1.19) | 0.65 |
| GCTA | 0.12 (41) | 0.13 (43) | 0.93 | 1.06 (0.63–1.79) | 0.92 | 0.87 (0.41–1.11) | 0.60 |
| CACG | 0.06 (21) | 0.14 (46) | <0.001 | 0.51 (0.28–0.92) | 0.035 | 0.78 (0.45–1.33) | 0.43 |
| CCTG | 0.09 (31) | 0.11 (36) | 0.70 | 0.96 (0.55–1.69) | 1.00 | 1.05 (0.61–1.83) | 0.96 |
| eNOS-Haplotype CTAAAT | |||||
|---|---|---|---|---|---|
| Model | a β ± SE | Wald Test | p Value | R2h | AIC |
| Dominant | 0.33 ± 0.43 | 0.77 | 0.440 | 0.6805 | 5346.49 |
| Recessive | 0.22 ± 0.29 | 0.76 | 0.291 | 0.6192 | 5892.21 |
| Multiplicative | 2.19 ± 0.86 | 2.55 | <0.001 | 1.000 | 3336.28 |
| General (0 copy) | −0.30 ± 0.39 | −0.77 | 0.440 | 0.9790 | 5342.96 |
| General (1 copy) | 2.10 ± 0.82 | 2.57 | 0.010 | 0.9790 | 5342.96 |
| ACE-Haplotype ACDG | |||||
| Dominant | 1.73 ± 0.54 | 3.19 | 0.001 | 1.000 | 1076.05 |
| Recessive | 0.43± 0.89 | 0.48 | 0.626 | 0.896 | 1298.44 |
| Multiplicative | 0.10 ± 0.36 | 0.29 | 0.769 | 0.916 | 1082.75 |
| General (0 copy) | −0.88± 0.32 | −2.70 | 0.006 | 0.891 | 1090.10 |
| General (1 copy) | −0.05± 0.41 | −0.12 | 0.898 | 0.891 | 1090.10 |
| VEGFA-Haplotype GATA | |||||
| Dominant | 3.07 ± 0.81 | 3.79 | <0.001 | 1.000 | 5324.21 |
| Recessive | 0.36 ± 0.43 | 0.84 | 0.399 | 0.680 | 5337.60 |
| Multiplicative | 0.59 ± 0.48 | 1.23 | 0.216 | 0.722 | 5335.36 |
| General (0 copy) | −0.58 ± 0.45 | −1.28 | 0.198 | 0.935 | 5621.86 |
| General (1 copy) | 0.004 ± 0.14 | 0.028 | 0.977 | 0.935 | 5621.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Singh, M.; Khinda, R.; Valecha, S.; Kumar, N.; Singh, S.; Juneja, P.K.; Kaur, T.; Mastana, S. Genetic Scores of eNOS, ACE and VEGFA Genes Are Predictive of Endothelial Dysfunction Associated Osteoporosis in Postmenopausal Women. Int. J. Environ. Res. Public Health 2021, 18, 972. https://doi.org/10.3390/ijerph18030972
Singh P, Singh M, Khinda R, Valecha S, Kumar N, Singh S, Juneja PK, Kaur T, Mastana S. Genetic Scores of eNOS, ACE and VEGFA Genes Are Predictive of Endothelial Dysfunction Associated Osteoporosis in Postmenopausal Women. International Journal of Environmental Research and Public Health. 2021; 18(3):972. https://doi.org/10.3390/ijerph18030972
Chicago/Turabian StyleSingh, Puneetpal, Monica Singh, Rubanpal Khinda, Srishti Valecha, Nitin Kumar, Surinderpal Singh, Pawan K. Juneja, Taranpal Kaur, and Sarabjit Mastana. 2021. "Genetic Scores of eNOS, ACE and VEGFA Genes Are Predictive of Endothelial Dysfunction Associated Osteoporosis in Postmenopausal Women" International Journal of Environmental Research and Public Health 18, no. 3: 972. https://doi.org/10.3390/ijerph18030972
APA StyleSingh, P., Singh, M., Khinda, R., Valecha, S., Kumar, N., Singh, S., Juneja, P. K., Kaur, T., & Mastana, S. (2021). Genetic Scores of eNOS, ACE and VEGFA Genes Are Predictive of Endothelial Dysfunction Associated Osteoporosis in Postmenopausal Women. International Journal of Environmental Research and Public Health, 18(3), 972. https://doi.org/10.3390/ijerph18030972