
Changing the Recipe: Pathogen Directed Changes in Tick Saliva Components



Abstract
:1. Introduction
2. Effect of Saliva on Pathogen Transmission
Saliva Assisted Transmission (SAT)
3. Role of Tick Salivary Components during Tick Feeding and Pathogen Transmission
3.1. Effects on Hemostasis and Angiogenesis
3.2. Immunomodulatory Components: Effects on Host Defenses
3.2.1. Host Defense
3.2.2. Complement
3.2.3. Immune Cells
3.2.4. Cytokine and Chemokine Secretion
3.2.5. Histamine Secretion
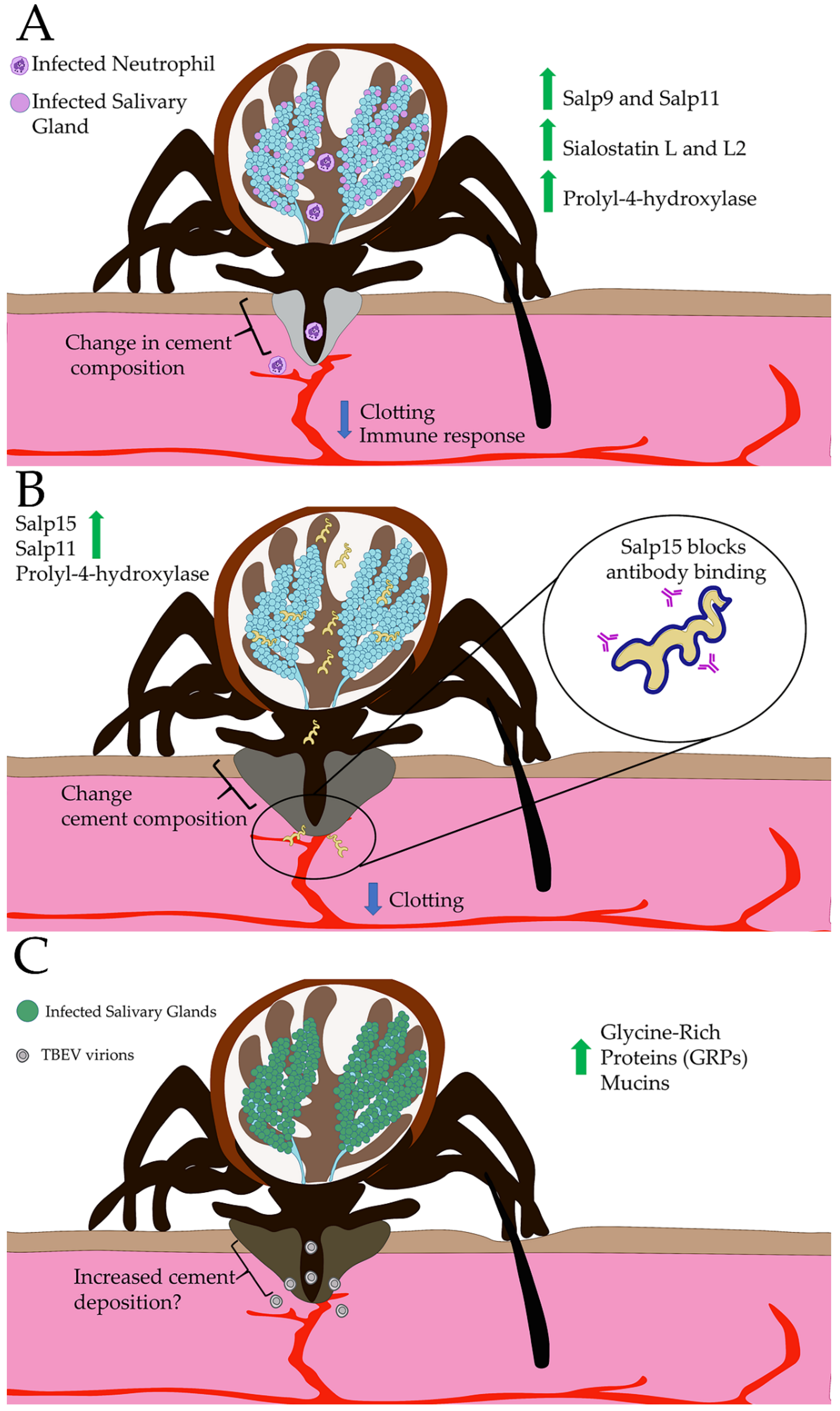
4. Global Manipulation of Tick Sialome by Pathogens
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.C.; Murrell, A. Systematics and evolution of ticks with a list of valid genus and species names. Parasitology 2004, 129, S15–S36. [Google Scholar] [CrossRef] [PubMed]
- Wikel, S.K. Tick-Host Interactions. In Biology of Ticks, 2nd ed.; Sonenshine, D., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2, p. 88. [Google Scholar]
- Alarcon-Chaidez, F. Salivary glands. In Biology of Ticks, 2nd ed.; Sonenshine, D., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 1. [Google Scholar]
- Anderson, J.F.; Magnarelli, L.A. Biology of ticks. Infect. Dis. Clin. N. Am. 2008, 22, 195–215. [Google Scholar] [CrossRef]
- Richter, D.; Matuschka, F.-R.; Spielman, A.; Mahadevan, L. How Ticks Get Under your Skin: Insertion Mechanics of the Feeding Apparatus of Ixodes ricinus Ticks. Proc. Biol. Sci. 2013, 280, 20131758. [Google Scholar] [CrossRef] [Green Version]
- Brossard, M.; Wikel, S.K. Tick Immunobiology. Parasitology 2004, 129, S161–S176. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.B.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M.C. The Role of Saliva in Tick Feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef] [Green Version]
- Lebre, M.C.; Am van, d.A.; van, B.L.; van, C.T.M.; Schuitemaker, J.H.; Kapsenberg, M.L.; de, J.E.C. Human Keratinocytes Express Functional Toll-like Receptor 3, 4, 5, and 9. J. Investig. Dermatol. 2007, 127. [Google Scholar] [CrossRef] [Green Version]
- Nestle, F.O.; Di Meglio, P.; Qin, J.-Z.; Nickoloff, B.J. Skin Immune Sentinels in Health and Disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Klicznik, M.M.; Szenes-Nagy, A.B.; Campbell, D.J.; Gratz, I.K. Taking the Lead-How Keratinocytes Orchestrate Skin T cell Immunity. Immunol. Lett. 2018, 200, 43–51. [Google Scholar] [CrossRef]
- Maouia, A.; Rebetz, J.; Kapur, R.; Semple, J.W. The Immune Nature of Platelets Revisited. Transfus. Med. Rev. 2020, 34, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Zuchtriegel, G.; Uhl, B.; Puhr-Westerheide, D.; Pörnbacher, M.; Lauber, K.; Krombach, F.; Reichel, C.A. Platelets Guide Leukocytes to Their Sites of Extravasation. PLoS Biol. 2016, 14, e1002459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissenpfennig, A.; Henri, S.; Dubois, B.; Laplace-Builhé, C.; Perrin, P.; Romani, N.; Tripp, C.H.; Douillard, P.; Leserman, L.; Kaiserlian, D.; et al. Dynamics and Function of Langerhans Cells in vivo: Dermal Dendritic cells Colonize Lymph Node Areas Distinct from Slower Migrating Langerhans cells. Immunity 2005, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nithiuthai, S.; Allen, J.R. Langerhans cells Present Tick Antigens to Lymph Node cells from Tick-sensitized Guinea-pigs. Immunology 1985, 55, 157. [Google Scholar]
- Gillespie, R.D.; Dolan, M.C.; Piesman, J.; Titus, R.G. Identification of an IL-2 Binding Protein in the Saliva of the Lyme Disease Vector Tick, Ixodes scapularis. J. Immunol. 2001, 166, 4319–4326. [Google Scholar] [CrossRef] [Green Version]
- Hajnická, V.; Vančová-Štibrániová, I.; Slovák, M.; Kocáková, P.; Nuttall, P.A. Ixodid Tick Salivary Gland Products Target Host Wound Healing Growth Factors. Int. J. Parasitol. 2011, 41, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, A.; Diouf, I.; Bakkali, N.; Missé, D.; Pagès, F.; Fusai, T.; Rogier, C.; Almeras, L. Implication of Haematophagous Arthropod Salivary Proteins in Host-vector Interactions. Parasit. Vectors 2011, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuttall, P.A. Tick Saliva and its Role in Pathogen Transmission. Wien. Klin. Wochenschr. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.C.; Mather, T.N. Ixodes scapularis:Salivary Kininase Activity Is a Metallo Dipeptidyl Carboxypeptidase. Exp. Parasitol. 1998, 89, 213–221. [Google Scholar] [CrossRef]
- Hidano, A.; Konnai, S.; Yamada, S.; Githaka, N.; Isezaki, M.; Higuchi, H.; Nagahata, H.; Ito, T.; Takano, A.; Ando, S.; et al. Suppressive Effects of Neutrophil by Salp16-like Salivary Gland Proteins from Ixodes persulcatus Schulze tick. Insect Mol. Biol. 2014, 23, 466–474. [Google Scholar] [CrossRef]
- Jaworski, D.C.; Jasinskas, A.; Metz, C.N.; Bucala, R.; Barbour, A.G. Identification and Characterization of a Homologue of the Pro-inflammatory Cytokine Macrophage Migration Inhibitory Factor in the Tick, Amblyomma americanum. Insect Mol. Biol. 2001, 10, 323–331. [Google Scholar] [CrossRef]
- Páleníková, J.; Lieskovská, J.; Langhansová, H.; Kotsyfakis, M.; Chmelař, J.; Kopecký, J. Ixodes ricinus Salivary Serpin IRS-2 Affects Th17 Differentiation via Inhibition of the Interleukin-6/STAT-3 Signaling Pathway. Infect. Immun. 2015, 83, 1949–1956. [Google Scholar] [CrossRef] [Green Version]
- Preston, S.G.; Majtán, J.; Kouremenou, C.; Rysnik, O.; Burger, L.F.; Cabezas Cruz, A.; Chiong Guzman, M.; Nunn, M.A.; Paesen, G.C.; Nuttall, P.A.; et al. Novel Immunomodulators from Hard Ticks Selectively Reprogramme Human Dendritic Cell Responses. PLoS Pathog. 2013, 9, e1003450. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The Essential Role of Tick Salivary Glands and Saliva in Tick Feeding and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Becchimanzi, A.; Tate, R.; Campbell, E.M.; Gigliotti, S.; Bowman, A.S.; Pennacchio, F. A Salivary Chitinase of Varroa destructor Influences Host Immunity and Mite’s Survival. PLoS Pathog. 2020, 16, e1009075. [Google Scholar] [CrossRef]
- Richards, E.H.; Jones, B.; Bowman, A. Salivary Secretions from the Honeybee Mite, Varroa destructor: Effects on Insect Haemocytes and Preliminary Biochemical Characterization. Parasitology 2011, 138, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, J.S.; Chavez, J.D.; Johnson, R.; Hosseinzadeh, S.; Mahoney, J.E.; Mohr, J.P.; Robison, F.; Zhong, X.; Hall, D.G.; MacCoss, M.; et al. Protein Interaction Networks at the Host-microbe Interface in Diaphorina citri, the Insect Vector of the Citrus Greening Pathogen. R. Soc. Open Sci. 2017, 4, 160545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlovskis, Z.; Canale, M.C.; Thole, V.; Pecher, P.; Lopes, J.R.; Hogenhout, S.A. Insect-borne Plant Pathogenic Bacteria: Getting a Ride Goes Beyond Physical Contact. Curr. Opin. Insect Sci. 2015, 9, 16–23. [Google Scholar] [CrossRef]
- Arena, G.D.; Ramos-Gonzalez, P.L.; Rogerio, L.A.; Ribeiro-Alves, M.; Casteel, C.L.; Freitas-Astua, J.; Machado, M.A. Making a Better Home: Modulation of Plant Defensive Response by Brevipalpus Mites. Front. Plant Sci. 2018, 9, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogenhout, S.A.; Bos, J.I.B. Effector Proteins that Modulate Plant–insect Interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Bosque-Pérez, N.A.; Davis, T.S. Insect-Borne Plant Pathogens and Their Vectors: Ecology, Evolution, and Complex Interactions. Annu. Rev. Entomol. 2018, 63, 169–191. [Google Scholar] [CrossRef]
- Su, Y.L.; Li, J.M.; Li, M.; Luan, J.B.; Ye, X.D.; Wang, X.W.; Liu, S.S. Transcriptomic Analysis of the Salivary Glands of an Invasive Whitefly. PLoS ONE 2012, 7, e39303. [Google Scholar] [CrossRef] [PubMed]
- Reppert, E.; Galindo, R.C.; Ayllón, N.; Breshears, M.A.; Kocan, K.M.; Blouin, E.F.; de la Fuente, J. Studies of Anaplasma phagocytophilum in Sheep Experimentally Infected with the Human NY-18 Isolate: Characterization of Tick Feeding Sites. Ticks Tick Borne Dis. 2014, 5, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granquist, E.G.; Aleksandersen, M.; Bergström, K.; Dumler, S.J.; Torsteinbø, W.O.; Stuen, S. A Morphological and Molecular Study of Anaplasma phagocytophilum Transmission Events at the Time of Ixodes ricinus Tick Bite. Acta Vet. Scand. 2010, 52, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, C.M.; Chao, L.L.; Yu, C.P. Chemotactic Migration of the Lyme Disease Spirochete (Borrelia burgdorferi) to Salivary Gland Extracts of Vector Ticks. Am. J. Trop. Med. Hyg. 2002, 66, 616–621. [Google Scholar] [CrossRef] [Green Version]
- Kotsyfakis, M.; Horka, H.; Salat, J.; Andersen, J.F. The crystal structures of two salivary cystatins from the tick Ixodes scapularis and the effect of these inhibitors on the establishment of Borrelia burgdorferi infection in a murine model. Mol Microbiol 2010, 77, 456–470. [Google Scholar] [CrossRef] [Green Version]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.B.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M.C. Antiinflammatory and Immunosuppressive Activity of Sialostatin L, a Salivary Cystatin from the Tick Ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsing, L.C.; Rudensky, A.Y. The Lysosomal Cysteine Proteases in MHC class II Antigen Presentation. Immunol. Rev. 2005, 207, 229–241. [Google Scholar] [CrossRef]
- Klein, M.; Brühl, T.J.; Staudt, V.; Reuter, S.; Grebe, N.; Gerlitzki, B.; Hoffmann, M.; Bohn, T.; Ulges, A.; Stergiou, N.; et al. Tick Salivary Sialostatin L Represses the Initiation of Immune Responses by Targeting IRF4-Dependent Transcription in Murine Mast Cells. J. Immunol. 2015, 195, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieskovská, J.; Páleníková, J.; Langhansová, H.; Chagas, A.C.; Calvo, E.; Kotsyfakis, M.; Kopecký, J. Tick Sialostatins L and L2 Differentially Influence Dendritic Cell Responses to Borrelia Spirochetes. Parasit. Vectors 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieskovská, J.; Páleníková, J.; Širmarová, J.; Elsterová, J.; Kotsyfakis, M.; Campos Chagas, A.; Calvo, E.; Růžek, D.; Kopecký, J. Tick Salivary Cystatin Sialostatin L2 Suppresses IFN Responses in Mouse Dendritic Cells. Parasite Immunol. 2015, 37, 70–78. [Google Scholar] [CrossRef]
- Sá-Nunes, A.; Bafica, A.; Antonelli, L.R.; Choi, E.Y.; Francischetti, I.M.; Andersen, J.F.; Shi, G.P.; Chavakis, T.; Ribeiro, J.M.; Kotsyfakis, M. The Immunomodulatory Action of Sialostatin L on Dendritic Cells Reveals its Potential to Interfere with Autoimmunity. J. Immunol. 2009, 182, 7422–7429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Wang, X.; Severo, M.S.; Sakhon, O.S.; Sohail, M.; Brown, L.J.; Sircar, M.; Snyder, G.A.; Sundberg, E.J.; Ulland, T.K.; et al. The Tick Salivary Protein Sialostatin L2 Inhibits Caspase-1-Mediated Inflammation during Anaplasma phagocytophilum Infection. Infect. Immun. 2014, 82, 2553–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.; Benz, E.J.; Silberstein, L.E.; Heslop, H.; Weitz, J.; Anastasi, J. Overview of Hemostasis and Thrombosis. In Hematology: Basic Principles and Practice, 6th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2012; pp. 1774–1783. [Google Scholar]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Narasimhan, S.; Koski, R.A.; Beaulieu, B.; Anderson, J.F.; Ramamoorthi, N.; Kantor, F.; Cappello, M.; Fikrig, E. A Novel Family of Anticoagulants from the Saliva of Ixodes scapularis. Insect Mol. Biol. 2002, 11. [Google Scholar] [CrossRef]
- Prevot, P.-P.; Adam, B.; Boudjeltia, K.Z.; Brossard, M.; Lins, L.; Cauchie, P.; Brasseur, R.; Vanhaeverbeek, M.; Vanhamme, L.; Godfroid, E. Anti-hemostatic Effects of a Serpin from the Saliva of the Tick Ixodes ricinus. J. Biol. Chem. 2006, 281, 26361–26369. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Narasimhan, S.; Zhang, L.; Liu, L.; Wang, P.; Fikrig, E. Tick Histamine Release Factor Is Critical for Ixodes scapularis Engorgement and Transmission of the Lyme Disease Agent. PLoS Pathogen. 2010, 6, e1001205. [Google Scholar] [CrossRef]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin, a Plasminogen Activator, Is Key to the Availability of Blood-Meals for Ixodid Ticks. PLoS Pathogen. 2011, 7, e1001312. [Google Scholar] [CrossRef]
- Batista, I.F.C.; Ramos, O.H.P.; Ventura, J.S.; Junqueira-de-Azevedo, I.L.M.; Ho, P.L.; Chudzinski-Tavassi, A.M. A New Factor Xa Inhibitor from Amblyomma cajennense with a Unique Domain Composition. Arch. Biochem. Biophys. 2010, 493, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Branco, V.G.; Iqbal, A.; Alvarez-Flores, M.P.; Sciani, J.M.; Andrade, S.A.; Iwai, L.K.; Serrano, S.M.T.; Chudzinski-Tavassi, A.M. Amblyomin-X having a Kunitz-type Homologous Domain, is a Noncompetitive Inhibitor of FXa and Induces Anticoagulation in vitro and in vivo. Biochim. Biophys. Acta Proteins Proteom. 2016, 1864, 1428–1435. [Google Scholar] [CrossRef]
- Carneiro-Lobo, T.C.; Konig, S.; Machado, D.E.; Nasciutti, L.E.; Forni, M.F.; Francischetti, I.M.B.; Sogayar, M.C.; Monteiro, R.Q. Ixolaris, a Tissue Factor Inhibitor, Blocks Primary Tumor Growth and Angiogenesis in a Glioblastoma Model. J. Thromb. Haemost. 2009, 7, 1855–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Wu, H.; Li, D. An Inhibitor Selective for Collagen-stimulated Platelet Aggregation from the Salivary glands of Hard tick Haemaphysalis longicornis and its Mechanism of Action. Sci. China Life Sci. 1999, 42, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Chmelar, J.; Oliveira, C.J.; Rezacova, P.; Francischetti, I.M.B.; Kovarova, Z.; Pejler, G.; Kopacek, P.; Ribeiro, J.M.C.; Mares, M.; Kopecky, J.; et al. A tick salivary protein targets cathepsin G and chymase and inhibits host inflammation and platelet aggregation. Blood 2011, 117, 736–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decrem, Y.; Beaufays, J.; Blasioli, V.; Lahaye, K.; Brossard, M.; Vanhamme, L.; Godfroid, E. A Family of Putative Metalloproteases in the Salivary Glands of the Tick Ixodes ricinus. FEBS J. 2008, 275, 1485–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decrem, Y.; Rath, G.; Blasioli, V.; Cauchie, P.; Robert, S.; Beaufays, J.; Frère, J.-M.; Feron, O.; Dogné, J.-M.; Dessy, C.; et al. Ir-CPI, a Coagulation Contact Phase Inhibitor from the Tick Ixodes ricinus, Inhibits Thrombus Formation Without Impairing Hemostasis. J. Exp. Med. 2009, 206, 2381–2395. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Martín, V.; Manzano-Román, R.; Oleaga, A.; Encinas-Grandes, A.; Pérez-Sánchez, R. Cloning and Characterization of a Plasminogen-binding Enolase from the Saliva of the Argasid tick Ornithodoros moubata. Vet. Parasitol. 2013, 191, 301–314. [Google Scholar] [CrossRef]
- Francischetti, I.M.B.; Valenzuela, J.G.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M.C. Ixolaris, a Novel Recombinant Tissue Factor Pathway Inhibitor (TFPI) from the Salivary Gland of the Tick, Ixodes scapularis: Identification of Factor X and Factor Xa as Scaffolds for the Inhibition of Factor VIIa/tissue Factor Complex. Blood 2002, 99, 3602–3612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukumoto, S.; Sakaguchi, T.; You, M.; Xuan, X.; Fujisaki, K. Tick Troponin I-like Molecule is a Potent Inhibitor for Angiogenesis. Microvasc. Res. 2006, 71, 218–221. [Google Scholar] [CrossRef]
- Ibelli, A.M.G.; Kim, T.K.; Hill, C.C.; Lewis, L.A.; Bakshi, M.; Miller, S.; Porter, L.; Mulenga, A. A Blood Meal-Induced Ixodes scapularis Tick Saliva Serpin Inhibits Trypsin and Thrombin, and Interferes with Platelet Aggregation and Blood Clotting. Int. J. Parasitol. 2014, 44, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.K.; Tsuji, N.; Miyoshi, T.; Alim, M.A.; Huang, X.; Hatta, T.; Fujisaki, K. The Kunitz-like modulatory protein haemangin is vital for hard tick blood-feeding success. PLoS Pathog. 2009, 5, e1000497. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Endris, R.; Connolly, T.M. Disagregin is a Fibrinogen Receptor Antagonist Lacking the Arg-Gly-Asp Sequence from the Tick, Ornithodoros moubata. J. Biol. Chem. 1994, 269, 6702. [Google Scholar] [CrossRef]
- Kim, T.K.; Tirloni, L.; Radulovic, Z.; Lewis, L.; Bakshi, M.; Hill, C.; da Silva Vaz, I.; Logullo, C.; Termignoni, C.; Mulenga, A. Conserved Amblyomma americanum Tick Serpin19, an Inhibitor of Blood Clotting Factors Xa and XIa, Trypsin and Plasmin, Has Anti-haemostatic Functions. Int. J. Parasitol. 2015, 45, 613–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, C.Y.; Kazimirova, M.; Trimnell, A.; Takac, P.; Labuda, M.; Nuttall, P.A.; Kini, R.M. Variegin, a Novel Fast and Tight Binding Thrombin Inhibitor from the Tropical Bont Tick. J. Biol. Chem. 2007, 282, 29101–29113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, C.Y.; Kumar, S.; Kazimirova, M.; Nuttall, P.A.; Radhakrishnan, U.P.; Kim, S.; Jagadeeswaran, P.; Imamura, T.; Mizuguchi, J.; Iwanaga, S.; et al. Crystal Structure of Thrombin in Complex with S-variegin: Insights of a Novel Mechanism of Inhibition and Design of Tunable Thrombin Inhibitors. PLoS ONE 2011, 6, e26367. [Google Scholar] [CrossRef] [Green Version]
- Mans, B.J.; Louw, A.I.; Neitz, A.W.H. Savignygrin, a Platelet Aggregation Inhibitor from the Soft Tick Ornithodoros savignyi, Presents the RGD Integrin Recognition Motif on the Kunitz-BPTI Fold. J. Biol. Chem. 2002, 277, 21371–21378. [Google Scholar] [CrossRef] [Green Version]
- Mans, B.J.; Ribeiro, J.M.C. Function, Mechanism and Evolution of the Moubatin-clade of Soft Tick Lipocalins. Insect Biochem. Mol. Biol. 2008, 38, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Mulenga, A.; Macaluso, K.R.; Simser, J.A.; Azad, A.F. The American Dog Tick, Dermacentor variabilis, Encodes a Functional Histamine Release Factor Homolog. Insect Biochem. Mol. Biol. 2003, 33, 911–919. [Google Scholar] [CrossRef]
- Nazareth, R.A.; Tomaz, L.S.; Ortiz-Costa, S.; Atella, G.C.; Ribeiro, J.M.C.; Francischetti, I.M.B.; Monteiro, R.Q. Antithrombotic Properties of Ixolaris, a Potent Inhibitor of the Extrinsic Pathway of the Coagulation Cascade. Thromb. Haemost. 2006, 96, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Nienaber, J.; Gaspar, A.R.M.; Neitz, A.W.H. Savignin, a Potent Thrombin Inhibitor Isolated from the Salivary Glands of the Tick Ornithodoros savignyi (Acari: Argasidae). Exp. Parasitol. 1999, 93, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Poole, N.M.; Mamidanna, G.; Smith, R.A.; Coons, L.B.; Cole, J.A. Prostaglandin E(2) in Tick Saliva Regulates Macrophage Cell Migration and Cytokine Profile. Parasit. Vectors 2013, 6, 261. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.; Makoul, G.T.; Levine, J.; Robinson, D.R.; Spielman, A. Antihemostatic, Antiinflammatory, and Immunosuppressive Properties of the Saliva of a Tick, Ixodes dammini. J. Exp. Med. 1985, 161, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Schuijt, T.J.; Bakhtiari, K.; Daffre, S.; DePonte, K.; Wielders, S.J.H.; Marquart, J.A.; Hovius, J.W.; van der Poll, T.; Fikrig, E.; Bunce, M.W.; et al. Factor Xa Activation of Factor V is of Paramount Importance in Initiating the Coagulation System: Lessons from a Tick Salivary Protein. Circulation 2013, 128, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Stutzer, C.; Mans, B.J.; Gaspar, A.R.; Neitz, A.W.; Maritz-Olivier, C. Ornithodoros savignyi: Soft Tick Apyrase Belongs to the 5’-nucleotidase Family. Exp. Parasitol. 2009, 122, 318–327. [Google Scholar] [CrossRef] [PubMed]
- van de Locht, A.; Stubbs, M.T.; Bode, W.; Friedrich, T.; Bollschweiler, C.; Höffken, W.; Huber, R. The Ornithodorin-thrombin Crystal Structure, a Key to the TAP enigma? EMBO J. 1996, 15, 6011–6017. [Google Scholar] [CrossRef]
- Wang, X.; Coons, L.B.; Taylor, D.B.; Stevens, S.E.; Gartner, T.K. Variabilin, a Novel RGD-containing Antagonist of Glycoprotein IIb-IIIa and Platelet Aggregation Inhibitor from the Hard Tick Dermacentor variabilis. J. Biol. Chem. 1996, 271, 17785–17790. [Google Scholar] [CrossRef] [Green Version]
- Waxman, L.; Connolly, T. Isolation of an Inhibitor Selective for Collagen-stimulated Platelet Aggregation from the Soft Tick Ornithodoros moubata. J. Biol. Chem. 1993, 268, 5445. [Google Scholar] [CrossRef]
- Waxman, L.; Smith, D.; Arcuri, K.; Vlasuk, G. Tick Anticoagulant Peptide (TAP) is a Novel Inhibitor of Blood Coagulation Factor Xa. Science 1990, 248, 593–596. [Google Scholar] [CrossRef]
- Xu, T.; Lew-Tabor, A.; Rodriguez-Valle, M. Effective Inhibition of Thrombin by Rhipicephalus microplus Serpin-15 (RmS-15) Obtained in the Yeast Pichia pastoris. Ticks Tick Borne Diseas. 2016, 7, 180–187. [Google Scholar] [CrossRef]
- Štibrániová, I.; BartÍKovÁ, P.; Holíková, V.; Kazimírová, M. Deciphering Biological Processes at the Tick-Host Interface Opens New Strategies for Treatment of Human Diseases. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmelař, J.; Kotál, J.; Karim, S.; Kopacek, P.; Francischetti, I.M.B.; Pedra, J.H.F.; Kotsyfakis, M. Sialomes and Mialomes: A Systems-Biology View of Tick Tissues and Tick–Host Interactions. Trend. Parasitol. 2016, 32, 242–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotál, J.; Langhansová, H.; Lieskovská, J.; Andersen, J.F.; Francischetti, I.M.B.; Chavakis, T.; Kopecký, J.; Pedra, J.H.F.; Kotsyfakis, M.; Chmelař, J. Modulation of Host Immunity by Tick Saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, J.G. Exploring Tick Saliva: From Biochemistry to ’Sialomes’ and Functional Genomics. Parasitology 2004, 129, S83–S94. [Google Scholar] [CrossRef] [PubMed]
- Anguita, J.; Ramamoorthi, N.; Hovius, J.W.R.; Das, S.; Thomas, V.; Persinski, R.; Conze, D.; Askenase, P.W.; Rincón, M.; Kantor, F.S.; et al. Salp15, an Ixodes scapularis Salivary Protein, Inhibits CD4+ T Cell Activation. Immunity 2002, 16, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Beaufays, J.; Adam, B.; Menten-Dedoyart, C.; Fievez, L.; Grosjean, A.; Decrem, Y.; Prévôt, P.-P.; Santini, S.; Brasseur, R.; Brossard, M.; et al. Ir-LBP, an Ixodes ricinus Tick Salivary LTB4-Binding Lipocalin, Interferes with Host Neutrophil Function. PLoS ONE 2008, 3, e3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blisnick, A.A.; Šimo, L.; Grillon, C.; Fasani, F.; Brûlé, S.; Le Bonniec, B.; Prina, E.; Marsot, M.; Relmy, A.; Blaise-Boisseau, S.; et al. The Immunomodulatory Effect of IrSPI, a Tick Salivary Gland Serine Protease Inhibitor Involved in Ixodes ricinus Tick Feeding. Vaccines 2019, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Daix, V.; Schroeder, H.; Praet, N.; Georgin, J.P.; Chiappino, I.; Gillet, L.; Fays, K.; Decrem, Y.; Leboulle, G.; Godfroid, E.; et al. Ixodes Ticks Belonging to the Ixodes ricinus Complex Encode a Family of Anticomplement Proteins. Insect Mol. Biol. 2007, 16, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Esteves, E.; Bizzarro, B.; Costa, F.B.; Ramírez-Hernández, A.; Peti, A.P.F.; Cataneo, A.H.D.; Wowk, P.F.; Timóteo, R.P.; Labruna, M.B.; Silva Junior, P.I.; et al. Amblyomma sculptum Salivary PGE(2) Modulates the Dendritic Cell-Rickettsia rickettsii Interactions in vitro and in vivo. Front. Immunol. 2019, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Garg, R.; Juncadella, I.J.; Ramamoorthi, N.; Ashish; Ananthanarayanan, S.K.; Thomas, V.; Rincón, M.; Krueger, J.K.; Fikrig, E.; Yengo, C.M.; et al. Cutting Edge: CD4 is the Receptor for the Tick Saliva Immunosuppressor, Salp15. J. Immunol. 2006, 177, 6579–6583. [Google Scholar] [CrossRef]
- Hannier, S.; Liversidge, J.; Sternberg, J.M.; Bowman, A.S. Characterization of the B-cell Inhibitory Protein Factor in Ixodes ricinus Tick Saliva: A Potential Role in Enhanced Borrelia burgdoferi Transmission. Immunology 2004, 113, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Hourcade, D.E.; Akk, A.M.; Mitchell, L.M.; Zhou, H.-f.; Hauhart, R.; Pham, C.T.N. Anti-Complement Activity of the Ixodes scapularis Salivary Protein Salp20. Mol. Immunol. 2016, 69, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Hovius, J.W.R.; de Jong, M.A.W.P.; den Dunnen, J.; Litjens, M.; Fikrig, E.; van der Poll, T.; Gringhuis, S.I.; Geijtenbeek, T.B.H. Salp15 Binding to DC-SIGN Inhibits Cytokine Expression by Impairing both Nucleosome Remodeling and mRNA Stabilization. PLoS Pathog. 2008, 4, e31. [Google Scholar] [CrossRef] [Green Version]
- Inokuma, H.; Kemp, D.H.; Willadsen, P. Prostaglandin E2 Production by the Cattle Tick (Boophilus microplus) into Feeding Sites and its Effect on the Response of Bovine Mononuclear Cells to Mitogen. Vet. Parasitol. 1994, 53, 293–299. [Google Scholar] [CrossRef]
- Katharine, R.T.; Christopher, E.; Aravinda, M.d.S. A Novel Mechanism of Complement Inhibition Unmasked by a Tick Salivary Protein That Binds to Properdin. J. Immunol. 2008, 180, 3964–3968. [Google Scholar] [CrossRef] [Green Version]
- Kotál, J.; Stergiou, N.; Buša, M.; Chlastáková, A.; Beránková, Z.; Řezáčová, P.; Langhansová, H.; Schwarz, A.; Calvo, E.; Kopecký, J.; et al. The Structure and Function of Iristatin, a Novel Immunosuppressive Tick Salivary Cystatin. Cell. Mol. Life Sci. 2019, 76, 2003–2013. [Google Scholar] [CrossRef]
- Leboulle, G.; Crippa, M.; Decrem, Y.; Mejri, N.; Brossard, M.; Bollen, A.; Godfroid, E. Characterization of a Novel Salivary Immunosuppressive Protein from Ixodes ricinus ticks. J. Biol. Chem. 2002, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Renneker, S.; Beyer, D.; Kullmann, B.; Seitzer, U.; Ahmed, J.; Bakheit, M.A. Identification and Partial Characterization of a Salp15 Homolog from Ixodes ricinus. Ticks Tick Borne Dis. 2014, 5, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Anguita, J.; Norgard, M.V.; Kantor, F.S.; Anderson, J.F.; et al. The Lyme Disease Agent Exploits a Tick Protein to Infect the Mammalian Host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sá-Nunes, A.; Bafica, A.; Lucas, D.A.; Conrads, T.P.; Veenstra, T.D.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M.; Francischetti, I.M. Prostaglandin E2 is a Major Inhibitor of Dendritic Cell Maturation and Function in Ixodes scapularis Saliva. J. Immunol. 2007, 179, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Wang, F.; Pan, W.; Wu, Q.; Wang, J.; Dai, J. An Immunosuppressive Tick Salivary Gland Protein DsCystatin Interferes With Toll-Like Receptor Signaling by Downregulating TRAF6. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Chen, W.; Mo, G.; Chen, R.; Fang, M.; Yedid, G.; Yan, X. An Immunosuppressant Peptide from the Hard Tick Amblyomma variegatum. Toxins 2016, 8, 133. [Google Scholar] [CrossRef] [Green Version]
- Tyson, K.; Elkins, C.; Patterson, H.; Fikrig, E.; Silva, A. Biochemical and Functional Characterization of Salp20, an Ixodes scapularis Tick Salivary Protein that Inhibits the Complement Pathway. Insect Mol. Biol. 2007, 16, 469–479. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Charlab, R.; Mather, T.N.; Ribeiro, J.M. Purification, Cloning, and Expression of a Novel Salivary Anticomplement Protein from the Tick, Ixodes scapularis. J. Biol. Chem. 2000, 275. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Liang, J.; Yu, H.; Wu, H.; Xu, C.; Liu, J.; Lai, R. A Tick B-cell Inhibitory Protein from Salivary Glands of the Hard Tick, Hyalomma asiaticum asiaticum. Biochem. Biophys. Res. Comm. 2006, 343, 585–590. [Google Scholar] [CrossRef]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A Key System for Immune Surveillance and Homeostasis. Nat. Immunol. 2010, 11. [Google Scholar] [CrossRef] [Green Version]
- Jason, R.D.; Wen-Chao, S. Complement and its Role in Innate and Adaptive Immune Responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Lubbers, R.; van, E.M.F.; van, K.C.; La, T. Production of Complement Components by Cells of the Immune System. Clin. Exp. Immunol. 2017, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, E.S.; Mastellos, D.C.; Hajishengallis, G.; Lambris, J.D. New Insights Into the Immune Functions of Complement. Nat. Rev. Immunol. 2019, 19. [Google Scholar] [CrossRef]
- Dulipati, V.; Meri, S.; Panelius, J. Complement Evasion Strategies of Borrelia burgdorferi sensu lato. FEBS Lett. 2020, 594, 2645–2656. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; Coumou, J.; Narasimhan, S.; Dai, J.; Deponte, K.; Wouters, D.; Brouwer, M.; Oei, A.; Roelofs, J.J.; van, D.A.P.; et al. A Tick Mannose-binding Lectin Inhibitor Interferes with the Vertebrate Complement Cascade to Enhance Transmission of the Lyme Disease Agent. Cell Host Microbe 2011, 10. [Google Scholar] [CrossRef] [Green Version]
- Wagemakers, A.; Coumou, J.; Schuijt, T.J.; Oei, A.; Nijhof, A.M.; van ’t Veer, C.; van der Poll, T.; Bins, A.D.; Hovius, J.W. An Ixodes ricinus Tick Salivary Lectin Pathway Inhibitor Protects Borrelia burgdorferi sensu lato from Human Complement. Vector Borne Zoonotic Dis. 2016, 16, 223–228. [Google Scholar] [CrossRef]
- Kirchner, S.; Lei, V.; MacLeod, A.S. The Cutaneous Wound Innate Immunological Microenvironment. Int. J. Mol. Sci. 2020, 21, 8748. [Google Scholar] [CrossRef]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The Immunological Anatomy of the Skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef]
- Brossard, M.; Wikel, S.K. Immunology of Interactions Between Ticks and Hosts. Med. Vet. Entomol. 1997, 11, 270–276. [Google Scholar] [CrossRef]
- Wikel, S.K. Host Immunity to Ticks. Ann. Rev. Entomol. 1996, 41, 1–22. [Google Scholar] [CrossRef]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of Acquired Resistance to Ticks. Front. Immunol. 2020, 11, 601504. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.R. Immunology of Interactions Between Ticks and Laboratory Animals. Exp. App. Acarol. 1989, 7, 5–13. [Google Scholar] [CrossRef]
- Trager, W. Acquired Immunity to Ticks. J. Parasit. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Lynn, G.E.; Diktas, H.; DePonte, K.; Fikrig, E. Naturally Acquired Resistance to Ixodes scapularis Elicits Partial Immunity against Other Tick Vectors in a Laboratory Host. Am. J. Trop. Med. Hyg. 2021, 104, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Ismail, N.; Soong, L.; Popov, V.L.; Whitworth, T.; Bouyer, D.H.; Walker, D.H. Differential interaction of dendritic cells with Rickettsia conorii: Impact on host susceptibility to murine spotted fever rickettsiosis. Infect Immun 2007, 75, 3112–3123. [Google Scholar] [CrossRef] [Green Version]
- Zeidner, N.S.; Schneider, B.S.; Rutherford, J.S.; Dolan, M.C. Suppression of Th2 Cytokines Reduces Tick-transmitted Borrelia burgdorferi Load in Mice. J. Parasitol. 2008, 94, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Hovius, J.W.; Schuijt, T.J.; Groot, K.A.; Roelofs, J.J.T.H.; Oei, G.A.; Marquart, J.A.; Beer, R.; van ’t Veer, C.; van der Poll, T.; Ramamoorthi, N.; et al. Preferential Protection of Borrelia burgdorferi Sensu Stricto by a Salp15 Homologue in Ixodes ricinus Saliva. J. Infect. Dis. 2008, 198, 1189–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchal, C.; Schramm, F.; Kern, A.; Luft, B.J.; Yang, X.; Schuijt, T.; Hovius, J.; Jaulhac, B.; Boulanger, N. Antialarmin Effect of Tick Saliva during the Transmission of Lyme Disease. Infect. Immun. 2010, 79, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukumaran, B.; Narasimhan, S.; Anderson, J.F.; DePonte, K.; Marcantonio, N.; Krishnan, M.N.; Fish, D.; Telford, S.R.; Kantor, F.S.; Fikrig, E. An Ixodes scapularis Protein Required for Survival of Anaplasma phagocytophilum in Tick Salivary Glands. J. Exp. Med. 2006, 203, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Hiroshi, U.; Hiroshi, T.; Tetsuya, K.; Masutaka, F. Cytokines and Chemokines in the Epidermis. J. Dermatol. Sci. 2000, 24, S29–S38. [Google Scholar] [CrossRef]
- Larry, C.B.; John, W.S. 2. Cytokines and Chemokines. J. Allerg. Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef]
- Strle, K.; Sulka, K.B.; Pianta, A.; Crowley, J.T.; Arvikar, S.L.; Anselmo, A.; Sadreyev, R.; Steere, A.C. T-Helper 17 Cell Cytokine Responses in Lyme Disease Correlate With Borrelia burgdorferi Antibodies During Early Infection and With Autoantibodies Late in the Illness in Patients With Antibiotic-Refractory Lyme Arthritis. Clin. Infect. Dis. 2017, 64, 930–938. [Google Scholar] [CrossRef] [Green Version]
- Storan, E.R.; O’Gorman, S.M.; McDonald, I.D.; Steinhoff, M. Role of Cytokines and Chemokines in Itch. Handb. Exp. Pharmacol. 2015, 226, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J.; Clemetson, J.M.; Proudfoot, A.E.; Power, C.A.; Baggiolini, M.; Wells, T.N. Functional Expression of CCR1, CCR3, CCR4, and CXCR4 Chemokine Receptors on Human Platelets. Blood 2000, 96, 4046–4054. [Google Scholar] [CrossRef]
- Akdis, C.A.; Blaser, K. Histamine in the Immune Regulation of Allergic Inflammation. J. Allergy Clin. Immunol. 2003, 112, 15–22. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Francischetti, I.M.; Pham, V.M.; Garfield, M.K.; Mather, T.N.; Ribeiro, J.M. Exploring the Sialome of the Tick Ixodes scapularis. J. Exp. Biol. 2002, 205, 2843–2864. [Google Scholar] [PubMed]
- Ribeiro, J.M.C.; Mans, B.J. TickSialoFam (TSFam): A Database That Helps to Classify Tick Salivary Proteins, a Review on Tick Salivary Protein Function and Evolution, With Considerations on the Tick Sialome Switching Phenomenon. Front. Cell. Infect. Microbiol. 2020, 10, 374. [Google Scholar] [CrossRef] [PubMed]
- Ayllón, N.; Villar, M.; Galindo, R.C.; Kocan, K.M.; Šíma, R.; López, J.A.; Vázquez, J.; Alberdi, P.; Cabezas-Cruz, A.; Kopáček, P.; et al. Systems Biology of Tissue-specific Response to Anaplasma phagocytophilum Reveals Differentiated Apoptosis in the Tick Vector Ixodes scapularis. PLoS Genet. 2015, 11, e1005120. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.C.; Alarcon-Chaidez, F.; Francischetti, I.M.B.; Mans, B.J.; Mather, T.N.; Valenzuela, J.G.; Wikel, S.K. An Annotated Catalog of Salivary Gland Transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 2006, 36, 111–129. [Google Scholar] [CrossRef]
- Cotté, V.; Sabatier, L.; Schnell, G.; Carmi-Leroy, A.; Rousselle, J.-C.; Arsène-Ploetze, F.; Malandrin, L.; Sertour, N.; Namane, A.; Ferquel, E.; et al. Differential Expression of Ixodes ricinus Salivary Gland Proteins in the Presence of the Borrelia burgdorferi sensu lato Complex. J. Proteom. 2014, 96, 29–43. [Google Scholar] [CrossRef]
- Trentelman, J.J.A.; Sima, R.; Krezdorn, N.; Tomás-Cortázar, J.; Barriales, D.; Takumi, K.; Butler, J.M.; Sprong, H.; Klouwens, M.J.; Urbanova, V.; et al. A Combined Transcriptomic Approach to Identify Candidates for an Anti-tick Vaccine Blocking B. afzelii transmission. Sci. Rep. 2020, 10, 20061. [Google Scholar] [CrossRef] [PubMed]
- Valdés, J.J.; Cabezas-Cruz, A.; Sima, R.; Butterill, P.T.; Růžek, D.; Nuttall, P.A. Substrate Prediction of Ixodes ricinus Salivary Lipocalins Differentially Expressed During Borrelia afzelii Infection. Sci. Rep. 2016, 6, 32372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondanelli, G.; Volpi, C. The Double Life of Serotonin Metabolites: In the Mood for Joining Neuronal and Immune Systems. Curr. Opin. Immunol. 2020, 70, 1–6. [Google Scholar] [CrossRef]
- Hart, C.E.; Ribeiro, J.M.; Kazimirova, M.; Thangamani, S. Tick-Borne Encephalitis Virus Infection Alters the Sialome of Ixodes ricinus Ticks During the Earliest Stages of Feeding. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Labuda, M.; Trimnell, A.R.; Licková, M.; Kazimírová, M.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P.A. An Antivector Vaccine Protects Against a Lethal Vector-borne Pathogen. PLoS Pathog. 2006, 2, e27. [Google Scholar] [CrossRef] [Green Version]
- Hermance, M.E.; Widen, S.G.; Wood, T.G.; Thangamani, S. Ixodes scapularis Salivary Gland microRNAs are Differentially Expressed During Powassan Virus Transmission. Sci. Rep. 2019, 9, 13110. [Google Scholar] [CrossRef] [PubMed]
- Hackenberg, M.; Langenberger, D.; Schwarz, A.; Erhart, J.; Kotsyfakis, M. In silico Target Network Analysis of de Novo-Discovered, Tick Saliva-specific microRNAs Reveals Important Combinatorial Effects in their Interference with Vertebrate Host Physiology. RNA 2017, 23, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.I.; Nawaz, M.; Hassan, I.A.; Zhang, H.; Gong, H.; Cao, J.; Zhou, Y.; Zhou, J. A microRNA Profile of Saliva and Role of miR-375 in Haemaphysalis longicornis (Ixodida: Ixodidae). Parasit. Vectors 2019, 12, 68. [Google Scholar] [CrossRef]
- Nawaz, M.; Malik, M.I.; Zhang, H.; Gebremedhin, M.B.; Cao, J.; Zhou, Y.; Zhou, J. miRNA Profile of Extracellular Vesicles Isolated from Saliva of Haemaphysalis longicornis tick. Acta Trop. 2020, 212, 105718. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Malik, M.I.; Zhang, H.; Hassan, I.A.; Cao, J.; Zhou, Y.; Hameed, M.; Hussain Kuthu, Z.; Zhou, J. Proteomic Analysis of Exosome-Like Vesicles Isolated From Saliva of the Tick Haemaphysalis longicornis. Front. Cell. Infect. Microbiol. 2020, 10, 542319. [Google Scholar] [CrossRef]
- Zhou, W.; Tahir, F.; Wang, J.C.; Woodson, M.; Sherman, M.B.; Karim, S.; Neelakanta, G.; Sultana, H. Discovery of Exosomes From Tick Saliva and Salivary Glands Reveals Therapeutic Roles for CXCL12 and IL-8 in Wound Healing at the Tick-Human Skin Interface. Front. Cell. Dev. Biol. 2020, 8, 554. [Google Scholar] [CrossRef] [PubMed]
- Chávez, A.S.O.; O’Neal, A.J.; Santambrogio, L.; Kotsyfakis, M.; Pedra, J.H.F. Message in a Vesicle-Trans-kingdom Intercommunication at the Vector-host Interface. J. Cell. Sci. 2019, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Woodson, M.; Neupane, B.; Bai, F.; Sherman, M.B.; Choi, K.H.; Neelakanta, G.; Sultana, H. Exosomes Serve as Novel Modes of Tick-borne Flavivirus Transmission from Arthropod to Human cells and Facilitates Dissemination of Viral RNA and Proteins to the Vertebrate Neuronal Cells. PLoS Pathog. 2018, 14, e1006764. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Component | Function/Activity | Target Effector/Protein | Pathogen SAT | Tick Species | References |
|---|---|---|---|---|---|
| Apyrase | Inhibitor of platelet aggregation | Adenosine triphospahte (ATP), Adenosine diphosphate (ADP) | Ixodes scapularis, Ornithodoros savignyi | [73,75] | |
| Tick histamine release factor (tHRF) † | Binding to basophils, stimulates Histamine release, vasodilation | - | B. burgdorferi | Dermacentor andersoni, I. scapularis | [49,69] |
| Metalloproteases | Wound healing/angiogenesis | Fibrin | Ixodes ricinus | [56] | |
| Haemangin | Wound healing/angiogenesis | Trypsin, chymotrypsin, plasmin | Haemaphysalis longicornis | [62] | |
| HLTnl | Wound healing/angiogenesis | Competitive inhibitor of Vascular endothelial growth factor (VEGF) for VEGF receptor | H. longicornis | [60] | |
| PGE2 † | Wound healing/angiogenesis | PGE2 receptor, cyclic Adenosine monophosphate (AMP)-proteins kinase A (cAMP-PKA) | D. variabilis | [72] | |
| TAP | Blood coagulation | Factor Xa | Ornithodoros moubata | [79] | |
| Moubatin | Platelet activation and vasoconstriction inhibitor | Collagen-stimulated aggregation activator | O. moubata | [68,78] | |
| Disaggregin | Platelet aggregation | Platelet fibrinogen receptor | O. moubata | [63] | |
| Enolase | Blood coagulation | Fibrin, plasminogen receptor | O. moubata | [58] | |
| Savignygrin | Platelet Aggregation | Thrombin | Ornithodoros savignyi | [67] | |
| Longicornin | Platelet aggregation | Collagen | H. longicornis | [54] | |
| Ornithodorin | Blood coagulation | Thrombin | O. moubata | [76] | |
| Salp14 | Blood coagulation | Factor Xa | I. scapularis | [47] | |
| Variabilin | Platelet aggregation | Glycoprotein IIb-IIIa | D. variabilis | [77] | |
| Serpin19 | Blood coagulation | Factor Xa, factor XIa, trypsin, plasmin | Amblyomma americanum | [64] | |
| RmS-15 | Blood coagulation | Thrombin | Rhipicephalus (Boophilus) microplus | [80] | |
| Longistatin | Blood coagulation | Fibrin | H. longicornis | [50] | |
| IxscS-1E1 | Blood coagulation | Thrombin, trypsin | I. scapularis | [61] | |
| IRS-2 † | Blood coagulation | Inhibits Cathepsin G and chymase proteases | I. ricinus | [55] | |
| Ir-CP1 | Blood coagulation | Inhibitor of contact system proteins | I. ricinus | [57] | |
| Variegin | Blood coagulation | Direct competitive inhibitor of Thrombin | Amblyomma variegatum | [65,66] | |
| Amblyomin-X | Blood coagulation | Noncompetitive inhibitor of coagulation factor, Factor Xa | A. variegatum | [51,52] | |
| Ixolaris | Blood coagulation | Inhibitor of contact system proteins, binds to Factor Xa | I. scapularis | [53,59,70] | |
| Iris † | Blood coagulation | Thrombin, Factor Xa, tissue plasminogen activation inhibitor | I. ricinus | [48] | |
| Savignin | Blood coagulation | Thrombin | O. savignyi | [71] | |
| TSGP3 | Platelet aggreagation and vasoconstriction inhibitor | Inhibition Collagen-platelet binding and interaction with thromboxane A2 | O. savignyi | [68] | |
| TIX-5 | Blood coagulation | Factor Xa, factor V | I. scapularis | [74] |
| Component | Function/Activity | Target Effector/Protein | Cells Affected | Pathogen | Tick Species | References |
|---|---|---|---|---|---|---|
| † Iris | Th 2 response modulation | Reduction in TNFα, INFγ, IL-8, IL-6, and IL-1β expression | Macrophages, T-lymphocyte | I. ricinus | [97] | |
| Salp15 | IL-2 inhibitor, T-cell proliferation (I. scapularis), IL-10 secretion inhibitor (I. ricinus) | OspC | T cells and peripheral blood mononuclear cells (PBMCs) | B. burgdorferi | I. scapularis, I. ricinus | [86,91,94,99,100] |
| IL-2 Binding Protein | T cell proliferation | IL-2 | T cells and PBMCs | I. scapularis | [16] | |
| IR-LBP | Neutrophil migration | Leukotriene B4 | Neutrophils | I. ricinus | [86] | |
| Irac I & II | Complement inhibitor | C3 convertase | I. ricinus | [88] | ||
| Isac | Complement inhibitor | C3 convertase | I. scapularis | [104] | ||
| Salp16 Iper1 | Neutrophil migration and reactive oxygen species (ROS) inhibitor | Neutrophils | A. phagocytophilum | I. persuculatus | [21] | |
| Salp16 Iper2 | Neutrophil migration and ROS inhibitor | Neutrophils | A. phagocytophilum | I. persculutaus | [21] | |
| † IRS-2 | Immune inhibitor | IL-6, IL-9, and IL-17 secretion STAT-3 phosphorylation | Dendritic cells, Neutrophils, and Th17 cells | B. burgdorferi | I. ricinus | [23] |
| Sialostatin L and L2 | Immune inhibitor | Cathepsin L and S inhibitor, inflammasome formation | B. burgderfori, L2-TBEV and A. phagocytophilum | I. scapularis | [38,41,42] | |
| Japanin | Modulates DC maturation | Inhibits IL-1β, IL-6, IL-12, IFN-γ, and TNFα secretion, CD86 and CD83 expression. Enhances IL-10 secretion and CD274 expression | Dendritic cells | R. appendiculatus | [24] | |
| IrSPI | T cell proliferation | CXCL10, CCL7, CCL4, CCL5, Eotaxin, IFN-γ, IL-1β, IL-18, IL-13, IL-6, TNFα, IL-9, and Granulocyte macrophage-colony stimulating factor (GM-CSF) inhibition and IL-2 | CD4++ T cells | I. ricinus | [87] | |
| † PGE2 | Immune inhibitor | Inhibition of IL-12, TNFα, and CD40 and upregulation of IL-10 (I. scapularis), increased macrophage PGE2, CCL5, TNFα, and sTNFRI secretion (D. variabilis), and TNFα inhibition (A. sculptum) | Bovine mononuclear cells, DCs, and macrophages | Rickettsia rickettsii | R. (Boophilus) microplus, I. scapularis, D. variabilis, and Amblyomma sculptum | [72,90,95,101] |
| Macrophage Migration Inhibitory Factor (MIF) homolog | Macrophage migration | Macrophages | A. americanum | [22] | ||
| BIP (B-cell inhibitory protein) | B cell proliferation | B cells | B. burgdorferi | I. ricinus | [91] | |
| B-cell inhibitory factor (BIF) | B cell proliferation | B cells | Hyalomma asiaticum | [105] | ||
| Amregulin | Immune inhibitor and antioxidant | TNFα, IFN-γ, IL-1, IL-8, and Nitric Monoxide (NMO) inhibitor | A. variegatum | [102] | ||
| tHRF | Histamine release | Histamine release stimulation | Basophils | B. burgdorferi | [49] | |
| TSGP2/3 | Neutrophil migration and complement inhibition | Leukotriene B4 and C5 binding | O. savignyi | [68] | ||
| Salp20 | Complement inhibition | C3 convertase (from properdin displacement) | B. burgdorferi | I. scapularis | [93,96,104] | |
| Iristatin | T cell proliferation and immune inhibition | IFN-γ, IL-2, IL-4, IL-6, and IL-9 secretion, CD4+ T cell proliferation, neutrophil migration, and nitric oxide production | Neutrophils, macrophages, T and mast cells | I. ricinus | [96] | |
| DsCystatin | Immunomodulation | Cathepsin L and B inhibitor, TNFα, IL-6, IL1β, and IFNγ inhibition, and promotes TRAF6 degradation | Macrophages | B. burgdorferi | Dermacenter silvarum | [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, M.; Underwood, J.; Oliva Chávez, A.S. Changing the Recipe: Pathogen Directed Changes in Tick Saliva Components. Int. J. Environ. Res. Public Health 2021, 18, 1806. https://doi.org/10.3390/ijerph18041806
Pham M, Underwood J, Oliva Chávez AS. Changing the Recipe: Pathogen Directed Changes in Tick Saliva Components. International Journal of Environmental Research and Public Health. 2021; 18(4):1806. https://doi.org/10.3390/ijerph18041806
Chicago/Turabian StylePham, Michael, Jacob Underwood, and Adela S. Oliva Chávez. 2021. "Changing the Recipe: Pathogen Directed Changes in Tick Saliva Components" International Journal of Environmental Research and Public Health 18, no. 4: 1806. https://doi.org/10.3390/ijerph18041806
APA StylePham, M., Underwood, J., & Oliva Chávez, A. S. (2021). Changing the Recipe: Pathogen Directed Changes in Tick Saliva Components. International Journal of Environmental Research and Public Health, 18(4), 1806. https://doi.org/10.3390/ijerph18041806