
Substance P and Neurokinin 1 Receptor in Chronic Inflammation and Cancer of the Head and Neck: A Review of the Literature


 ,
,  and
and 
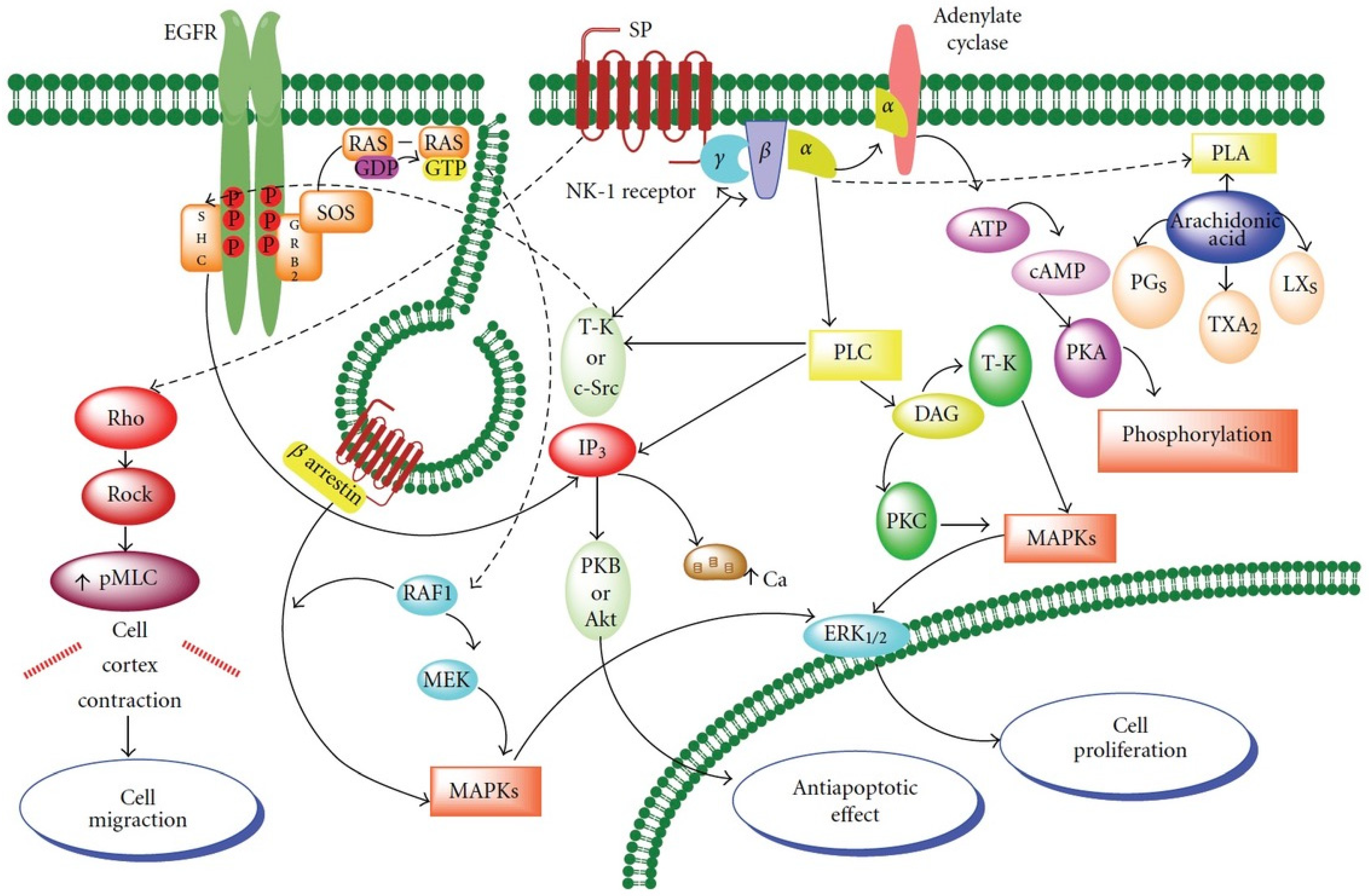{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. SP/NK1R in Head and Neck Mucosal Inflammation and Cancer
3. SP/NK1R in Head and Neck Cancer
3.1. Oral Potentially Malignant Disorders and Oral Cancer
3.2. Laryngeal Cancer
3.3. Thyroid Cancer
3.4. Relationship with Environmental Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, S.D.; Ferlito, A.; Takes, R.P.; Brakenhoff, R.H.; Valentin, M.D.; Woolgar, J.A.; Bradford, C.R.; Rodrigo, J.P.; Rinaldo, A.; Hier, M.P.; et al. Advances and applications of oral cancer basic research. Oral Oncol. 2011, 47, 783–791. [Google Scholar] [CrossRef]
- Polanska, H.; Raudenska, M.; Gumulec, J.; Sztalmachova, M.; Adam, V.; Kizek, R.; Masarik, M. Clinical significance of head and neck squamous cell cancer biomarkers. Oral Oncol. 2014, 50, 168–177. [Google Scholar] [CrossRef] [PubMed]
- González-Moles, M.Á.; Ramos-García, P.; Esteban, F. Significance of the Overexpression of Substance P and Its Receptor NK-1R in Head and Neck Carcinogenesis: A Systematic Review and Meta-Analysis. Cancers 2021, 13, 1349. [Google Scholar] [CrossRef] [PubMed]
- Hökfelt, T.; Pernow, B.; Wahren, J. Substance P: A pioneer amongst neuropeptides. J. Intern. Med. 2001, 249, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. Involvement of substance P and the NK-1 receptor in human pathology. Amino Acids 2014, 46, 1727–1750. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Rosso, M.; Pérez, A.; Coveñas, R.; Rosso, R.; Zamarriego, C.; Soult, J.A.; Montero, I. Antitumoral action of the neurokinin-1-receptor antagonist L-733,060 and mitogenic action of substance P on human retinoblastoma cell lines. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2567–2570. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Sharif, T.R.; Sharif, M. Substance P-induced mitogenesis in human astrocytoma cells correlates with activation of the mitogen-activated protein kinase signaling pathway. Cancer Res. 1996, 56, 4983–4991. [Google Scholar] [PubMed]
- Muñoz, M.; Pérez, A.; Rosso, M.; Zamarriego, C.; Rosso, R. Antitumoral action of the neurokinin-1 receptor antagonist L-733 060 on human melanoma cell lines. Melanoma Res. 2004, 14, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Rosso, M.; Pérez, A.; Coveñas, R.; Rosso, R.; Zamarriego, C.; Piruat, J.I. The NK1 receptor is involved in the antitumoural action of L-733,060 and in the mitogenic action of substance P on neuroblastoma and glioma cell lines. Neuropeptides 2005, 39, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Esteban, F.; Muñoz, M.; González-Moles, M.A.; Rosso, M. A role for substance P in cancer promotion and progression: A mechanism to counteract intracellular death signals following oncogene activation or DNA damage. Cancer Metastasis Rev. 2006, 25, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Seegers, H.C.; Hood, V.C.; Kidd, B.L.; Cruwys, S.C.; Walsh, D.A. Enhancement of angiogenesis by endogenous substance P release and neurokinin-1 receptors during neurogenic inflammation. J. Pharmacol. Exp. Ther. 2003, 306, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ma, G.; Ma, Q.; Li, W.; Liu, J.; Han, L.; Duan, W.; Xu, Q.; Liu, H.; Wang, Z.; et al. Neurotransmitter substance P mediates pancreatic cancer perineural invasion via NK-1R in cancer cells. Mol. Cancer Res. 2013, 11, 294–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Yuan, S.; Cheng, J.; Kang, S.; Zhao, W.; Zhang, J. Substance P Promotes the Progression of Endometrial Adenocarcinoma. Int. J. Gynecol. Cancer 2016, 26, 845–850. [Google Scholar] [CrossRef]
- Wang, L.; Wang, N.; Zhang, R.; Dong, D.; Liu, R.; Zhang, L.; Ji, W.; Yu, M.; Zhang, F.; Niu, R.; et al. TGFβ regulates NK1R-Tr to affect the proliferation and apoptosis of breast cancer cells. Life Sci. 2020, 256, 117674. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. Neurokinin receptor antagonism: A patent review (2014-present). Expert Opin. Ther. Pat. 2020, 30, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. The Neurokinin-1 Receptor Antagonist Aprepitant: An Intelligent Bullet against Cancer? Cancers 2020, 12, 2682. [Google Scholar] [CrossRef]
- Muñoz, M.; Coveñas, R. The Neurokinin-1 Receptor Antagonist Aprepitant, a New Drug for the Treatment of Hematological Malignancies: Focus on Acute Myeloid Leukemia. J. Clin. Med. 2020, 9, 1659. [Google Scholar] [CrossRef]
- Moles, M.A.G.; Esteban, F.; Ruiz-Avila, I.; Montoya, J.A.G.; Brener, S.; Bascones-Martínez, A.; Muñoz, M. A role for the substance P/NK-1 receptor complex in cell proliferation and apoptosis in oral lichen planus. Oral Dis. 2009, 15, 162–169. [Google Scholar] [CrossRef] [PubMed]
- González-Moles, M.A.; Brener, S.; Ruiz-Avila, I.; Gil-Montoya, J.A.; Tostes, D.; Bravo, M.; Esteban, F. Substance P and NK-1R expression in oral precancerous epithelium. Oncol. Rep. 2009, 22, 1325–1331. [Google Scholar] [CrossRef] [Green Version]
- Esteban, F.; Gonzalez-Moles, M.A.; Castro, D.; Martin-Jaen, M.d.M.; Redondo, M.; Ruiz-Avila, I.; Muñoz, M. Expression of substance P and neurokinin-1-receptor in laryngeal cancer: Linking chronic inflammation to cancer promotion and progression. Histopathology 2009, 54, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Isorna, I.; Esteban, F.; Solanellas, J.; Coveñas, R.; Muñoz, M. The substance P and neurokinin-1 receptor system in human thyroid cancer: An immunohistochemical study. Eur. J. Histochem. 2020, 64, 3117. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Manning, C.B.; Vallyathan, V.; Mossman, B.T. Diseases caused by asbestos: Mechanisms of injury and disease development. Int. Immunopharmacol. 2002, 2, 191–200. [Google Scholar] [CrossRef]
- Hilleman, M.R. Critical overview and outlook: Pathogenesis, prevention, and treatment of hepatitis and hepatocarcinoma caused by hepatitis B virus. Vaccine 2003, 21, 4626–4649. [Google Scholar] [CrossRef]
- Naumann, M.; Crabtree, J.E. Helicobacter pylori-induced epithelial cell signalling in gastric carcinogenesis. Trends Microbiol. 2004, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Itzkowitz, S.H. Cancer in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Aaltonen, L.M.; Rihkanen, H.; Vaheri, A. Human papillomavirus in larynx. Laryngoscope 2002, 112, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, H.; Paleri, V.; West, C.M.L.; Nutting, C. Head and neck cancer—Part 1: Epidemiology, presentation, and prevention. BMJ 2010, 341, c4684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnakulasuriya, S.; Kujan, O.; Aguirre-Urizar, J.M.; Bagan, J.V.; González-Moles, M.Á.; Kerr, A.R.; Lodi, G.; Mello, F.W.; Monteiro, L.; Ogden, G.R.; et al. Oral potentially malignant disorders: A consensus report from an international seminar on nomenclature and classification, convened by the WHO Collaborating Centre for Oral Cancer. Oral Dis. 2020, 27, 1862–1880. [Google Scholar] [CrossRef] [PubMed]
- Ramos-García, P.; Gonzalez-Moles, M.A.; Warnakulasuriya, S. Oral cancer development in lichen planus and related conditions -3.0 evidence level-: A systematic review of systematic reviews. Oral Dis. 2021, 27, 1919–1935. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 2010; Volume 93. [Google Scholar]
- Almendro, V.; Garcia-Recio, S.; Gascon, P. Tyrosine Kinase Receptor Transactivation Associated to G Protein-Coupled Receptors. Curr. Drug Targets 2010, 11, 1169–1180. [Google Scholar] [CrossRef]
- Ames, B.N.; Gold, L.S. Too many rodent carcinogens: Mitogenesis increases mutagenesis. Science 1990, 249, 970–971. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J. Cancer cell cycles. Science 1996, 274, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Weitzman, S.A.; Gordon, L.I. Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 1990, 76, 655–663. [Google Scholar] [CrossRef]
- Macarthur, M.; Hold, G.L.; El-Omar, E.M. Inflammation and Cancer II. Role of chronic inflammation and cytokine gene polymorphisms in the pathogenesis of gastrointestinal malignancy. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G515–G520. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, M.; Patsias, A.; Posner, M.; Sikora, A. The Role of Inflammation in Head and Neck Cancer. In Inflammation and Cancer; Springer: Basel, Switzerland, 2014; pp. 107–127. [Google Scholar]
- Aguirre-Urizar, J.M.; de Mendoza, L.-I.I.; Warnakulasuriya, S. Malignant transformation of oral leukoplakia: Systematic review and meta-analysis of the last 5 years. Oral Dis. 2021, 27, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Iocca, O.; Sollecito, T.P.; Alawi, F.; Weinstein, G.S.; Newman, J.G.; De Virgilio, A.; Di Maio, P.; Spriano, G.; López, S.P.; Shanti, R.M. Potentially malignant disorders of the oral cavity and oral dysplasia: A systematic review and meta-analysis of malignant transformation rate by subtype. Head Neck 2019, 42, 539–555. [Google Scholar] [CrossRef] [PubMed]
- González-Moles, M.Á.; Ramos-García, P.; Warnakulasuriya, S. An appraisal of highest quality studies reporting malignant transformation of oral lichen planus based on a systematic review. Oral Dis. 2020, 27, 1908–1918. [Google Scholar] [CrossRef]
- Ramos-García, P.; González-Moles, M.Á.; Mello, F.W.; Bagan, J.V.; Warnakulasuriya, S. Malignant transformation of oral proliferative verrucous leukoplakia: A systematic review and meta-analysis. Oral Dis. 2021, 27, 1896–1907. [Google Scholar] [CrossRef] [PubMed]
- Kujan, O.; Mello, F.W.; Warnakulasuriya, S. Malignant transformation of oral submucous fibrosis: A systematic review and meta-analysis. Oral Dis. 2020, 27, 1936–1946. [Google Scholar] [CrossRef] [PubMed]
- Odell, E.; Kujan, O.; Warnakulasuriya, S.; Sloan, P. Oral epithelial dysplasia: Recognition, grading and clinical significance. Oral Dis. 2021, 27, 1947–1976. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Moles, M.A.; Scully, C.; Gil-Montoya, J.A. Oral lichen planus: Controversies surrounding malignant transformation. Oral Dis. 2008, 14, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Mignogna, M.D.; Fedele, S.; Russo, L.L.; Muzio, L.L.; Bucci, E. Immune activation and chronic inflammation as the cause of malignancy in oral lichen planus: Is there any evidence? Oral Oncol. 2004, 40, 120–130. [Google Scholar] [CrossRef]
- González-Moles, M.A.; Bascones-Ilundain, C.; Montoya, J.A.G.; Ruiz-Avila, I.; Delgado-Rodríguez, M.; Bascones-Martínez, A. Cell cycle regulating mechanisms in oral lichen planus: Molecular bases in epithelium predisposed to malignant transformation. Arch. Oral Biol. 2006, 51, 1093–1103. [Google Scholar] [CrossRef]
- González-Moles, M.Á.; Ramos-García, P.; Warnakulasuriya, S. A Scoping Review on Gaps in the Diagnostic Criteria for Proliferative Verrucous Leukoplakia: A Conceptual Proposal and Diagnostic Evidence-Based Criteria. Cancers 2021, 13, 3669. [Google Scholar] [CrossRef]
- Abboud, C.S.; Brandão, E.H.d.S.; Cunha, K.R.L.; Brito, K.S.; Gallo, C.d.B.; Molon, A.C.; Horliana, A.C.R.T.; Franco, A.S.L.; Thongprasom, K.; Rodrigues, M.F.S.D. Serum and salivary cytokines in patients with oral lichen planus treated with Photobiomodulation. Oral Dis. 2021; Epub ahead of printing. [Google Scholar] [CrossRef]
- Kundu, P.; Pant, I.; Jain, R.; Rao, S.G.; Kondaiah, P. Genome-wide DNA methylation changes in oral submucous fibrosis. Oral Dis. 2021; Epub ahead of printing. [Google Scholar] [CrossRef]
- Brailo, V.; Vučićević-Boras, V.; Cekić-Arambašin, A.; Alajbeg, I.Ž.; Milenović, A.; Lukač, J. The significance of salivary interleukin 6 and tumor necrosis factor alpha in patients with oral leukoplakia. Oral Oncol. 2006, 42, 370–373. [Google Scholar] [CrossRef]
- Tin, L.L.; Yeh, J.; Friedman, J.; Yan, B.; Yang, X.; Yeh, N.T.; Van Waes, C.; Chen, Z. A signal network involving coactivated NF-κB and STAT3 and altered p53 modulates BAX/BCL-XL expression and promotes cell survival of head and neck squamous cell carcinomas. Int. J. Cancer 2008, 122, 1987–1998. [Google Scholar] [CrossRef]
- Lenouvel, D.; González-Moles, M.Á.; Talbaoui, A.; Ramos-García, P.; González-Ruiz, L.; Ruiz-Ávila, I.; Gil-Montoya, J.A. An update of knowledge on PD-L1 in head and neck cancers: Physiologic, prognostic and therapeutic perspectives. Oral Dis. 2020, 26, 511–526. [Google Scholar] [CrossRef]
- Wang, F.; Arun, P.; Friedman, J.; Chen, Z.; Waes, C.V. Current and potential inflammation targeted therapies in head and neck cancer. Curr. Opin. Pharmacol. 2009, 9, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.L.; Xin, P.Y.; Yan, B.; Friedman, J.; Duggal, P.; Bagain, L.; Dong, G.; Yeh, N.T.; Wang, J.; Zhou, J.; et al. A novel nuclear factor-κB gene signature is differentially expressed in head and neck squamous cell carcinomas in association with TP53 status. Clin. Cancer Res. 2007, 13, 5680–5691. [Google Scholar] [CrossRef] [Green Version]
- Duffy, S.; Teknos, T.; Islam, M.; Chen, Z.; Albert, P.S.; Wolf, G.; Waes, C.V. Nuclear factor-kappaB-related serum factors as longitudinal biomarkers of response and survival in advanced oropharyngeal carcinoma. Clin. Cancer Res. 2007, 13, 3182–3190. [Google Scholar]
- Rosso, M.; Muñoz, M.; Berger, M. The role of neurokinin-1 receptor in the microenvironment of inflammation and cancer. Sci. World J. 2012, 2012, 381434. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, T.M.; O’Connell, J.; O’Brien, D.I.; Goode, T.; Bredin, C.P.; Shanahan, F. The role of substance P in inflammatory disease. J. Cell. Physiol. 2004, 201, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Keränen, U.; Järvinen, H.; Kärkkäinen, P.; Kiviluoto, T.; Kivilaakso, E.; Soinila, S. Substance P—An underlying factor for pouchitis? Prospective study of substance P- and vasoactive intestinal polypeptide-immunoreactive innervation and mast cells. Dig. Dis. Sci. 1996, 41, 1665–1671. [Google Scholar] [CrossRef]
- Keränen, U.; Järvinen, H.; Kiviluoto, T.; Kivilaakso, E.; Soinila, S. Substance P- and vasoactive intestinal polypeptide-immunoreactive innervation in normal and inflamed pouches after restorative proctocolectomy for ulcerative colitis. Dig. Dis. Sci. 1996, 41, 1658–1664. [Google Scholar] [CrossRef]
- Muñoz, M.; Pérez, A.; Coveñas, R.; Rosso, M.; Castro, E. Antitumoural action of L-733,060 on neuroblastoma and glioma cell lines. Arch. Ital. Biol. 2004, 142, 105–112. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Coveñas, R. NK-1 receptor antagonists as new anti-tumoural agents: Action on human neuroblastoma cell lines. In Focus on Neuroblastoma Research; Fernandes, J., Ed.; Nova Science Publishers: New York, NY, USA, 2007; pp. 31–56. [Google Scholar]
- Munoz, M.; Rosso, M.; Covenas, R. A New Frontier in the Treatment of Cancer: NK-1 Receptor Antagonists. Curr. Med. Chem. 2010, 17, 504–516. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Aguilar, F.J.; González-Moles, M.A.; Redondo, M.; Esteban, F. NK-1 receptor antagonists induce apoptosis and counteract substance P-related mitogenesis in human laryngeal cancer cell line HEp-2. Investig. New Drugs 2008, 26, 111–118. [Google Scholar] [CrossRef]
- Muñoz, M.; Coveñas, R.; Esteban, F.; Redondo, M. The substance P/NK-1 receptor system: NK-1 receptor antagonists as anti-cancer drugs. J. Biosci. 2015, 40, 441–463. [Google Scholar] [CrossRef]
- Rupniak, N.M.J.; Carlson, E.; Boyce, S.; Webb, J.K.; Hill, R.G. Enantioselective inhibition of the formalin paw late phase by the NK1 receptor antagonist L-733,060 in gerbils. Pain 1996, 67, 189–195. [Google Scholar] [CrossRef]
- Bang, R.; Sass, G.; Kiemer, A.K.; Vollmar, A.M.; Neuhuber, W.L.; Tiegs, G. Neurokinin-1 receptor antagonists CP-96,345 and L-733,060 protect mice from cytokine-mediated liver injury. J. Pharmacol. Exp. Ther. 2003, 305, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVane, C.L. Substance P: A new era, a new role. Pharmacotherapy 2001, 21, 1061–1069. [Google Scholar] [CrossRef]
- Mantyh, P. Substance P and the Inflammatory and Immune Response. Ann. N. Y. Acad. Sci. 1991, 632, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Horstmann, S.; Kahle, P.J.; Borasio, G.D. Inhibitors of p38 mitogen-activated protein kinase promote neuronal survival in vitro. J. Neurosci. Res. 1998, 52, 483–490. [Google Scholar] [CrossRef]
- Brener, S.; González-Moles, M.A.; Tostes, D.; Esteban, F.; Gil-Montoya, J.A.; Ruiz-Avila, I.; Bravo, M.; Muñoz, M. A role for the substance P/NK-1 receptor complex in cell proliferation in oral squamous cell carcinoma. Anticancer Res. 2009, 29, 2323–2329. [Google Scholar]
- Nowicki, M.; Ostalska-Nowicka, D.; Konwerska, A.; Miskowiak, B. The predicting role of substance P in the neoplastic transformation of the hypoplastic bone marrow. J. Clin. Pathol. 2006, 59, 935–941. [Google Scholar] [CrossRef] [Green Version]
- Pascual, D.W.; Xu-Amano, J.; Kiyono, H.; McGhee, J.R.; Bost, K.L. Substance P acts directly upon cloned B lymphoma cells to enhance IgA and IgM production. J. Immunol. 1991, 146, 2130–2216. [Google Scholar]
- Feistritzer, C.; Clausen, J.; Sturn, D.H.; Djanani, A.; Gunsilius, E.; Wiedermann, C.J.; Kähler, C.M. Natural killer cell functions mediated by the neuropeptide substance P. Regul. Pept. 2003, 116, 119–126. [Google Scholar] [CrossRef]
- Walsh, D.T.; Weg, V.B.; Williams, T.J.; Nourshargh, S. Substance P-induced inflammatory responses in guinea-pig skin: The effect of specific NK1 receptor antagonists and the role of endogenous mediators. Br. J. Pharmacol. 1995, 114, 1343–1350. [Google Scholar] [CrossRef] [Green Version]
- Caviedes-Bucheli, J.; Gomez-Sosa, J.F.; Azuero-Holguin, M.M.; Ormeño-Gomez, M.; Pinto-Pascual, V.; Munoz, H.R. Angiogenic mechanisms of human dental pulp and their relationship with substance P expression in response to occlusal trauma. Int. Endod. J. 2017, 50, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, J.V.; Blum, A.; Walder, J.; Walder, R. Eosinophils from granulomas in murine Schistosomiasis mansoni produce substance P. J. Immunol. 1988, 141, 961–966. [Google Scholar] [PubMed]
- Cook, G.A.; Elliott, D.; Metwali, A.; Blum, A.M.; Sandor, M.; Lynch, R.; Weinstock, J.V. Molecular evidence that granuloma T lymphocytes in murine Schistosomiasis mansoni express an authentic substance P (NK-1) receptor. J. Immunol. 1994, 152, 1830–1835. [Google Scholar] [PubMed]
- Janes, S.M.; Lowell, S.; Hutter, C. Epidermal stem cells. J. Pathol. 2002, 197, 479–491. [Google Scholar] [CrossRef]
- Zhu, A.J.; Watt, F.M. Beta-catenin signalling modulates proliferative potential of human epidermal keratinocytes independently of intercellular adhesion. Development 1999, 126, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Tabor, M.P.; Brakenhoff, R.H.; van Houten, V.M.; Kummer, J.A.; Snel, M.H.; Snijders, P.J.; Snow, G.B.; Leemans, C.R.; Braakhuis, B.J. Persistence of genetically altered fields in head and neck cancer patients: Biological and clinical implications. Clin. Cancer Res. 2001, 7, 1523. [Google Scholar]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological effects of melatonin: Role of melatonin receptors and signal transduction pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef]
- Muñoz, M.; Pavón, A.; Rosso, M.; Salinas, M.V.; Pérez, A.; Carranza, A.; González-Ortega, A. Immunolocalization of NK-1 Receptor and Substance P in Human Normal Placenta. Placenta 2010, 31, 649–651. [Google Scholar] [CrossRef]
- Singh, S.; Kumaravel, S.; Dhole, S.; Roy, S.; Pavan, V.; Chakraborty, S. Neuropeptide Substance P Enhances Inflammation-Mediated Tumor Signaling Pathways and Migration and Proliferation of Head and Neck Cancers. Indian J. Surg. Oncol. 2021, 12, 93–102. [Google Scholar] [CrossRef]
- Ortiz-Prieto, A.; Bernabeu-Wittel, J.; Zulueta-Dorado, T.; Lorente-Lavirgen, A.I.; Muñoz, M. Immunolocalization of substance P and NK-1 receptor in vascular anomalies. Arch. Dermatol. Res. 2017, 309, 97–102. [Google Scholar] [CrossRef]
- Yilmaz, M.; Christofori, G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Walczak-Drzewiecka, A.; Ratajewski, M.; Wagner, W.; Dastych, J. HIF-1α Is Up-Regulated in Activated Mast Cells by a Process That Involves Calcineurin and NFAT. J. Immunol. 2008, 181, 1665–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieb, K.; Fiebich, B.L.; Berger, M.; Bauer, J.; Schulze-Osthoff, K. The neuropeptide substance P activates transcription factor NF-kappa B and kappa B-dependent gene expression in human astrocytoma cells. J. Immunol. 1997, 159, 4952–4958. [Google Scholar] [PubMed]
- Koon, H.W.; Zhao, D.; Na, X.; Moyer, M.P.; Pothoulakis, C. Metalloproteinases and transforming growth factor-α mediate substance p-induced mitogen-activated protein kinase activation and proliferation in human colonocytes. J. Biol. Chem. 2004, 279, 45519–45527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moles, M.A.G.; Mosqueda-Taylor, A.; Esteban, F.; Gil-Montoya, J.A.; Díaz-Franco, M.A.; Delgado, M.; Muñoz, M. Cell proliferation associated with actions of the substance P/NK-1 receptor complex in keratocystic odontogenic tumours. Oral Oncol. 2008, 44, 1127–1133. [Google Scholar] [CrossRef]
- Kramer, M.S.; Cutler, N.; Feighner, J.; Shrivastava, R.; Carman, J.; Sramek, J.J.; Reines, S.A.; Liu, G.; Snavely, D.; Wyatt-Knowles, E.; et al. Distinct mechanism for antidepressant activity by blockade of central substance P receptors. Science 1998, 281, 1640–1645. [Google Scholar] [CrossRef]
- Hon, W.K.; Pothoulakis, C. Immunomodulatory properties of substance P: The gastrointestinal system as a model. Ann. N. Y. Acad. Sci. 2006, 1088, 23–40. [Google Scholar] [CrossRef]
- Levy, M.J.; Classey, J.D.; Maneesri, S.; Meeran, K.; Powell, M.; Goadsby, P.J. The association between Calcitonin Gene-Related Peptide (CGRP), substance P and headache in pituitary tumours. Pituitary 2004, 7, 67–71. [Google Scholar] [CrossRef]
- Goso, C.; Potier, E.; Manzini, S.; Szallasi, A. Comparison of tachykinin NK1 receptors in human IM9 and U373 MG cells, using antagonist (FK888, (+/−)-CP-96,345, and RP 67580) binding. Eur. J. Pharmacol. 1994, 254, 221–227. [Google Scholar] [CrossRef]
- Ding, H.F.; Fisher, D.E. Induction of apoptosis in cancer: New therapeutic opportunities. Ann. Med. 2002, 34, 451–469. [Google Scholar] [CrossRef]
- Hennig, I.M.; Laissue, J.A.; Horisberger, U.; Reubi, J.C. Substance-P receptors in human primary neoplasms: Tumoral and vascular localization. Int. J. Cancer 1995, 61, 786–792. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Coveñas, R. The NK-1 receptor is involved in the antitumoral action of L-733,060 and in the mitogenic action of substance P on human pancreatic cancer cell lines. Lett. Drug Des. Discov. 2006, 3, 323–329. [Google Scholar] [CrossRef]
- Holm, R.; Sobrinho-Simões, M.; Nesland, J.M.; Gould, V.E.; Johannessen, J.V. Medullary carcinoma of the thyroid Gland: An immunocytochemical study. Ultrastruct. Pathol. 1985, 8, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. Involvement of substance P and the NK-1 receptor in cancer progression. Peptides 2013, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. Involvement of substance P and the NK-1 receptor in pancreatic cancer. World J. Gastroenterol. 2014, 20, 2321–2334. [Google Scholar] [CrossRef]
- Muñoz, M.; González-Ortega, A.; Rosso, M.; Robles-Frias, M.J.; Carranza, A.; Salinas-Martín, M.V.; Coveñas, R. The substance P/neurokinin-1 receptor system in lung cancer: Focus on the antitumor action of neurokinin-1 receptor antagonists. Peptides 2012, 38, 318–325. [Google Scholar] [CrossRef]
- Munoz, M.; Rosso, M.; Covenas, R. The NK-1 Receptor: A New Target in Cancer Therapy. Curr. Drug Targets 2011, 12, 909–921. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Robles-Frias, M.J.; Salinas-Martín, M.V.; Rosso, R.; González-Ortega, A.; Coveñas, R. The NK-1 receptor is expressed in human melanoma and is involved in the antitumor action of the NK-1 receptor antagonist aprepitant on melanoma cell lines. Lab. Investig. 2010, 90, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Ortega, E.; Monsalve-Guil, L.; Jiménez-Guerra, A.; Ortiz-García, I.; De La Rosa, J.C.C.; Coveñas, R.; Muñoz, M. Involvement of the substance p/neurokinin-1 receptor system in oral pain and inflammation. J. Biol. Regul. Homeost. Agents 2020, 34, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Reitsma, M.B.; Fullman, N.; Ng, M.; Salama, J.S.; Abajobir, A.; Abate, K.H.; Abbafati, C.; Abera, S.F.; Abraham, B.; Abyu, G.Y.; et al. Smoking prevalence and attributable disease burden in 195 countries and territories, 1990-2015: A systematic analysis from the Global Burden of Disease Study 2015. Lancet 2017, 389, 1885–1906. [Google Scholar] [CrossRef] [Green Version]
- Sekizawa, S.-I.; Joad, J.P.; Pinkerton, K.E.; Bonham, A.C. Distinct tachykinin NK(1) receptor function in primate nucleus tractus solitarius neurons is dysregulated after second-hand tobacco smoke exposure. Br. J. Pharmacol. 2011, 163, 782–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Moles, M.A.; Gutierrez, J.; Rodriguez, M.J.; Ruiz-Avila, I.; Rodriguez-Archilla, A. Epstein-Barr virus latent membrane protein-1 (LMP-1) expression in oral squamous cell carcinoma. Laryngoscope 2002, 112, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Leeder, J.S.; Piquette-Miller, M.; Dupuis, L.L. Aprepitant and fosaprepitant drug interactions: A systematic review. Br. J. Clin. Pharmacol. 2017, 83, 2148–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteban, F.; Ramos-García, P.; Muñoz, M.; González-Moles, M.Á. Substance P and Neurokinin 1 Receptor in Chronic Inflammation and Cancer of the Head and Neck: A Review of the Literature. Int. J. Environ. Res. Public Health 2022, 19, 375. https://doi.org/10.3390/ijerph19010375
Esteban F, Ramos-García P, Muñoz M, González-Moles MÁ. Substance P and Neurokinin 1 Receptor in Chronic Inflammation and Cancer of the Head and Neck: A Review of the Literature. International Journal of Environmental Research and Public Health. 2022; 19(1):375. https://doi.org/10.3390/ijerph19010375
Chicago/Turabian StyleEsteban, Francisco, Pablo Ramos-García, Miguel Muñoz, and Miguel Ángel González-Moles. 2022. "Substance P and Neurokinin 1 Receptor in Chronic Inflammation and Cancer of the Head and Neck: A Review of the Literature" International Journal of Environmental Research and Public Health 19, no. 1: 375. https://doi.org/10.3390/ijerph19010375
APA StyleEsteban, F., Ramos-García, P., Muñoz, M., & González-Moles, M. Á. (2022). Substance P and Neurokinin 1 Receptor in Chronic Inflammation and Cancer of the Head and Neck: A Review of the Literature. International Journal of Environmental Research and Public Health, 19(1), 375. https://doi.org/10.3390/ijerph19010375