
Edible Insect Consumption for Human and Planetary Health: A Systematic Review

 , , ,
, , ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Study Selection and Data Extraction
2.4. Quality Assessment
3. Results
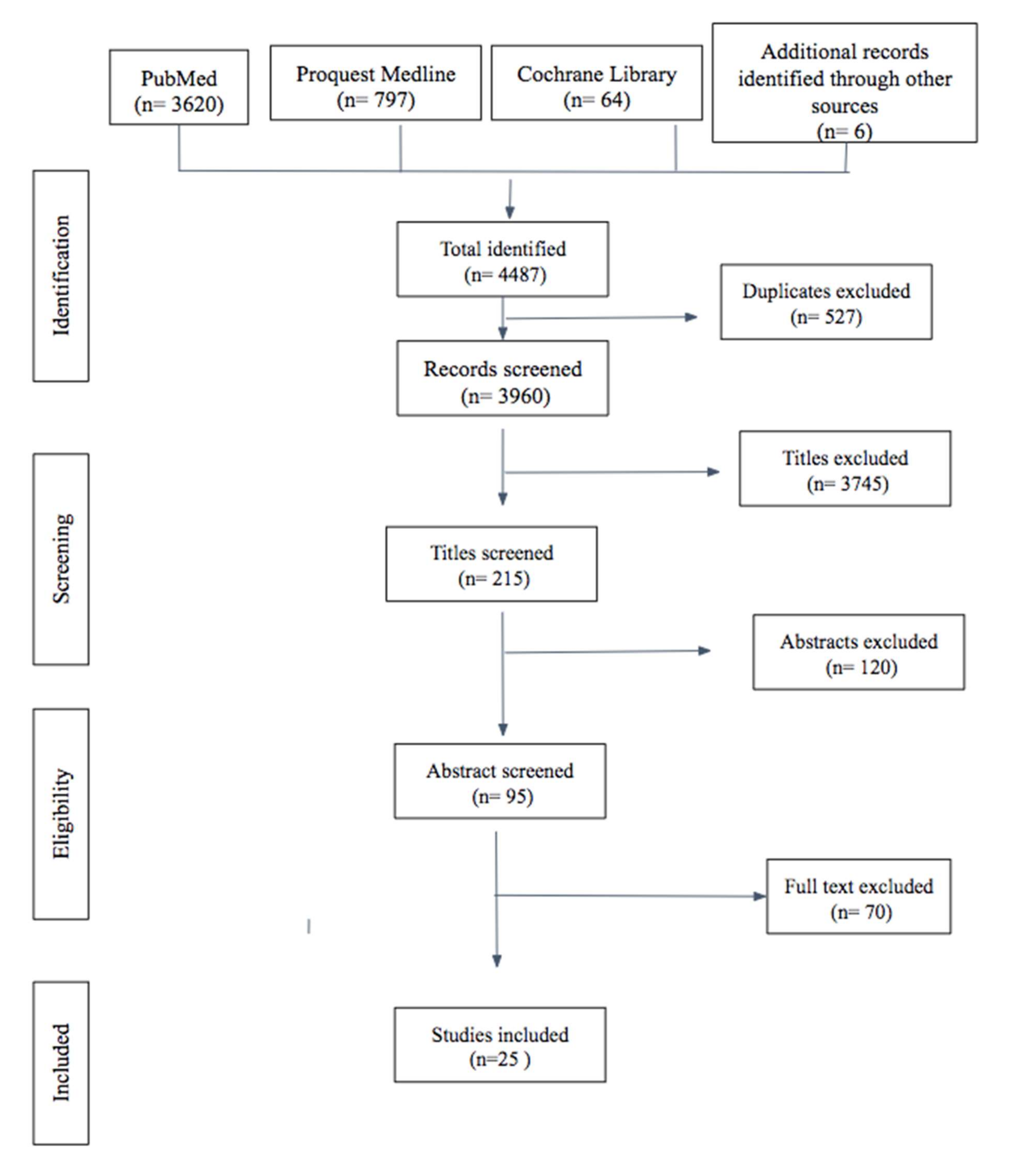
3.1. Literature Search Results
3.2. Health Outcomes in Animal Studies
3.2.1. Appetite Control
3.2.2. Growth Performance
3.2.3. Metabolic Traits
3.2.4. Crude Protein Digestibility
3.2.5. Intestinal Morphology
3.3. Health Outcomes in Human Studies
3.3.1. Growth Performance
3.3.2. Metabolic Traits
3.3.3. Gut Microbiota Composition
3.3.4. Changes in Muscle Mass Composition and Strength
3.3.5. Crude Protein Digestibility
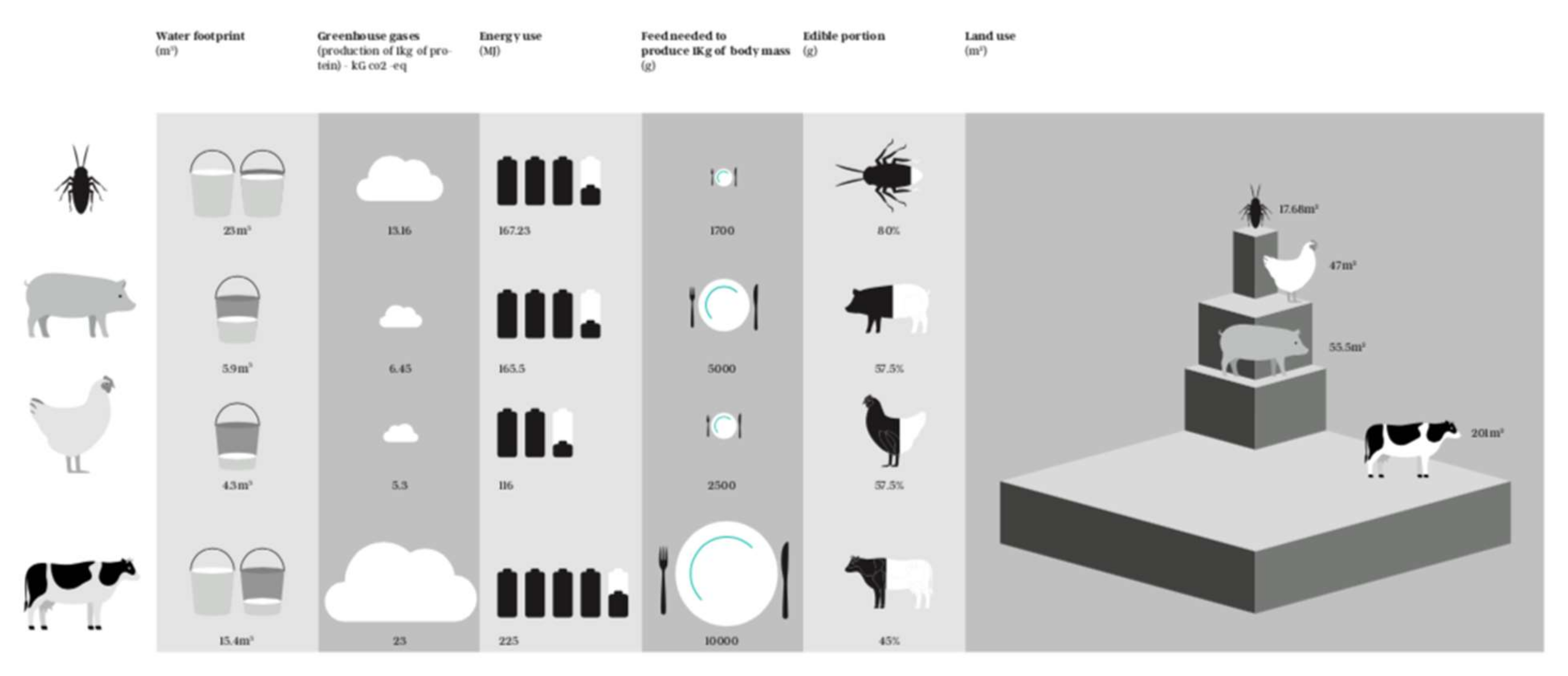
3.4. Environmental Impacts of Edible Insects
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Panel, G. Future Food Systems: For People, our Planet, and Prosperity; Global Panel on Agriculture and Food Systems for Nutrition: London, UK, 2020. [Google Scholar]
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; De Vries, W.; De Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [PubMed]
- Sterner, T.; Barbier, E.B.; Bateman, I.; Bijgaart, I.V.D.; Crépin, A.-S.; Edenhofer, O.; Fischer, C.; Habla, W.; Hassler, J.; Johansson-Stenman, O.; et al. Policy design for the Anthropocene. Nat. Sustain. 2019, 2, 14–21. [Google Scholar] [CrossRef]
- Gerten, D.; Heck, V.; Jägermeyr, J.; Bodirsky, B.L.; Fetzer, I.; Jalava, M.; Kummu, M.; Lucht, W.; Rockström, J.; Schaphoff, S.; et al. Feeding ten billion people is possible within four terrestrial planetary boundaries. Nat. Sustain. 2020, 3, 200–208. [Google Scholar] [CrossRef]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. Bioscience 2017, 67, 386–391. [Google Scholar] [CrossRef]
- Gu, D.; Andreev, K.; Dupre, M.E. Major trends in population growth around the world. China CDC Wkly. 2021, 3, 604. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans-a life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Murray, C.J. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Salter, A.M.; Lopez-Viso, C. Role of novel protein sources in sustainably meeting future global requirements. Proc. Nutr. Soc. 2021, 80, 186–194. [Google Scholar] [CrossRef]
- Weindl, I.; Ost, M.; Wiedmer, P.; Schreiner, M.; Neugart, S.; Klopsch, R.; Kühnhold, H.; Kloas, W.; Henkel, I.M.; Schlüter, O.; et al. Sustainable food protein supply reconciling human and ecosystem health: A Leibniz Position. Glob. Food Secur. 2020, 25, 100367. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Nuţă, D. New legal requirements regarding the placing of novel foods on the European Union market. Annals. Food Sci. Technol. 2017. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Knutsen, H.K. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar] [PubMed]
- Committee, E.S. Risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Knutsen, H.K. Safety of frozen and dried formulations from migratory locust (Locusta migratoria) as a Novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06667. [Google Scholar]
- Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K. Safety of partially defatted house cricket (Acheta domesticus) powder as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20, e07258. [Google Scholar] [PubMed]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K. EFSA NDA Panel (EFSA Panel on Nutrition, Novel Foods and Food Allergens), 2019. Sci. Opin. Saf. Phenylcapsaicin A Nov. Foodpursuant Regul. 2015, 2283. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Payne, C.L.R.; Scarborough, P.; Rayner, M.; Nonaka, K. Are edible insects more or less ‘healthy’ than commonly consumed meats? A comparison using two nutrient profiling models developed to combat over- and undernutrition. Eur. J. Clin. Nutr. 2016, 70, 285–291. [Google Scholar] [CrossRef]
- Da Silva Lucas, A.J.; de Oliveira, L.M.; Da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]
- Hlongwane, Z.T.; Slotow, R.; Munyai, T.C. Nutritional Composition of Edible Insects Consumed in Africa: A Systematic Review. Nutrients 2020, 12, 2786. [Google Scholar] [CrossRef]
- Weru, J.; Chege, P.; Kinyuru, J. Nutritional potential of edible insects: A systematic review of published data. Int. J. Trop. Insect Sci. 2021, 41, 2015–2037. [Google Scholar] [CrossRef]
- Bertola, M.; Mutinelli, F. A Systematic Review on Viruses in Mass-Reared Edible Insect Species. Viruses 2021, 13, 2280. [Google Scholar] [CrossRef] [PubMed]
- Ayensu, J.; Annan, R.A.; Edusei, A.; Lutterodt, H. Beyond nutrients, health effects of entomophagy: A systematic review. Nutr. Food Sci. 2018, 49, 2–17. [Google Scholar] [CrossRef]
- D’Antonio, V.; Battista, N.; Sacchetti, G.; Di Mattia, C.; Serafini, M. Functional properties of edible insects: A systematic review. Nutr. Res. Rev. 2021, 1–54. [Google Scholar] [CrossRef] [PubMed]
- Testa, M.; Stillo, M.; Maffei, G.; Andriolo, V.; Gardois, P.; Zotti, C.M. Ugly but tasty: A systematic review of possible human and animal health risks related to entomophagy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3747–3759. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.S.; Ayllón, T.; Malirat, V.; Câmara, D.C.P.; Dias, C.M.G.; Louzada, G.; Fernandes-Ferreira, D.; Medronho, R.D.A.; Acevedo, R.C. High Prevalence of a Newly Discovered Wutai Mosquito Phasivirus in Mosquitoes from Rio de Janeiro, Brazil. Insects 2019, 10, 135. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Moher, D. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Hall, H.; Fitches, E.; Smith, R. Insects as Animal Feed: Novel Ingredients for Use in Pet, Aquaculture and Livestock Diets; CABI: Wallingford, UK, 2021. [Google Scholar]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savović, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C.; et al. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef]
- Kim, J.; Yun, E.-Y.; Park, S.-W.; Goo, T.-W.; Seo, M. Allomyrina dichotoma larvae regulate food intake and body weight in high fat diet-induced obese mice through mTOR and Mapk signaling pathways. Nutrients 2016, 8, 100. [Google Scholar] [CrossRef]
- Seo, M.; Goo, T.W.; Chung, M.Y.; Baek, M.; Hwang, J.S.; Kim, M.A.; Yun, E.Y. Tenebrio molitor larvae inhibit adipogenesis through AMPK and MAPKs signaling in 3T3-L1 adipocytes and obesity in high-fat diet-induced obese mice. Int. J. Mol. Sci. 2017, 18, 518. [Google Scholar] [CrossRef]
- Bergmans, R.S.; Nikodemova, M.; Stull, V.J.; Rapp, A.; Malecki, K.M.C. Comparison of cricket diet with peanut-based and milk-based diets in the recovery from protein malnutrition in mice and the impact on growth, metabolism and immune function. PLoS ONE 2020, 15, e0234559. [Google Scholar] [CrossRef] [PubMed]
- Dabbou, S.; Gai, F.; Biasato, I.; Capucchio, M.T.; Biasibetti, E.; Dezzutto, D.; Meneguz, M.; Plachà, I.; Gasco, L.; Schiavone, A. Black soldier fly defatted meal as a dietary protein source for broiler chickens: Effects on growth performance, blood traits, gut morphology and histological features. J. Anim. Sci. Biotechnol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Bovera, F.; Loponte, R.; Pero, M.E.; Cutrignelli, M.I.; Calabrò, S.; Musco, N.; Vassalotti, G.; Panettieri, V.; Lombardi, P.; Piccolo, G.; et al. Laying performance, blood profiles, nutrient digestibility and inner organs traits of hens fed an insect meal from Hermetia illucens larvae. Res. Veter. Sci. 2018, 120, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Veter. Res. 2018, 14, 383. [Google Scholar] [CrossRef]
- Gasco, L.; Dabbou, S.; Trocino, A.; Xiccato, G.; Capucchio, M.T.; Biasato, I.; Dezzutto, D.; Birolo, M.; Meneguz, M.; Schiavone, A.; et al. Effect of dietary supplementation with insect fats on growth performance, digestive efficiency and health of rabbits. J. Anim. Sci. Biotechnol. 2019, 10, 4. [Google Scholar] [CrossRef]
- Agbemafle, I.; Hanson, N.; Bries, A.E.; Reddy, M.B. Alternative Protein and Iron Sources from Edible Insects but Not Solanum torvum Improved Body Composition and Iron Status in Malnourished Rats. Nutrients 2019, 11, 2481. [Google Scholar] [CrossRef] [PubMed]
- Pessina, F.; Frosini, M.; Marcolongo, P.; Fusi, F.; Saponara, S.; Gamberucci, A.; Dreassi, E. Antihypertensive, cardio-and neuro-protective effects of Tenebrio molitor (Coleoptera: Tenebrionidae) defatted larvae in spontaneously hypertensive rats. PLoS ONE 2020, 15, e0233788. [Google Scholar] [CrossRef]
- Ahn, M.Y.; Kim, B.J.; Kim, H.J.; Jin, J.M.; Yoon, H.J.; Hwang, J.S.; Lee, B.M. Anti-diabetic activity of field cricket glycosaminoglycan by ameliorating oxidative stress. BMC Complement. Med. Ther. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Ahn, M.Y.; Hwang, J.S.; Kim, M.-J.; Park, K.-K. Antilipidemic effects and gene expression profiling of the glycosaminoglycans from cricket in rats on a high fat diet. Arch. Pharmacal Res. 2016, 39, 926–936. [Google Scholar] [CrossRef]
- Skau, J.K.H.; Touch, B.; Chhoun, C.; Chea, M.; Unni, U.S.; Makurat, J.; Filteau, S.; Wieringa, F.T.; Dijkhuizen, M.A.; Ritz, C.; et al. Effects of animal source food and micronutrient fortification in complementary food products on body composition, iron status, and linear growth: A randomized trial in Cambodia. Am. J. Clin. Nutr. 2015, 101, 742–751. [Google Scholar] [CrossRef]
- Bauserman, M.; Lokangaka, A.; Gado, J.; Close, K.; Wallace, D.; Kodondi, K.-K.; Tshefu, A.; Bose, C. A cluster-randomized trial determining the efficacy of caterpillar cereal as a locally available and sustainable complementary food to prevent stunting and anaemia. Public Health Nutr. 2015, 18, 1785–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirmala, I.; Pramono, M.S. Sago worms as a nutritious traditional and alternative food for rural children in Southeast Sulawesi, Indonesia. Asia Pac. J. Clin. Nutr. 2017, 26, s40–s49. [Google Scholar] [PubMed]
- Stull, V.J.; Finer, E.; Bergmans, R.S.; Febvre, H.P.; Longhurst, C.; Manter, D.K.; Patz, J.A.; Weir, T.L. Impact of Edible Cricket Consumption on Gut Microbiota in Healthy Adults, a Double-blind, Randomized Crossover Trial. Sci. Rep. 2018, 8, 10762. [Google Scholar] [CrossRef]
- Vangsoe, M.T.; Joergensen, M.S.; Heckmann, L.-H.L.; Hansen, M. Effects of insect protein supplementation during resistance training on changes in muscle mass and strength in young men. Nutrients 2018, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Vangsoe, M.T.; Thogersen, R.; Bertram, H.C.; Heckmann, L.-H.L.; Hansen, M. Ingestion of Insect Protein Isolate Enhances Blood Amino Acid Concentrations Similar to Soy Protein in A Human Trial. Nutrients 2018, 10, 1357. [Google Scholar] [CrossRef] [PubMed]
- Lokman, I.H.; Ibitoye, E.B.; Hezmee, M.N.M.; Goh, Y.M.; Zuki, A.B.Z.; Jimoh, A.A. Effects of chitin and chitosan from cricket and shrimp on growth and carcass performance of broiler chickens. Trop. Anim. Health Prod. 2019, 51, 2219–2225. [Google Scholar] [CrossRef]
- Van Zanten, H.H.; Mollenhorst, H.; Oonincx, D.G.; Bikker, P.; Meerburg, B.G.; de Boer, I.J. From environmental nuisance to environmental opportunity: Housefly larvae convert waste to livestock feed. J. Clean. Prod. 2015, 102, 362–369. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental impact of food waste bioconversion by insects: Application of life cycle assessment to process using Hermetia illucens. J. Clean. Prod. 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Muys, B.; Roffeis, M. Life cycle assessment of proteins from insects. In Proceedings of the Insects to feed the world: 1st International Conference, Wageningen, The Netherlands, 14–17 May 2014. [Google Scholar]
- Thévenot, A.; Rivera, J.L.; Wilfart, A.; Maillard, F.; Hassouna, M.; Senga-Kiesse, T.; LE Feon, S.; Aubin, J. Mealworm meal for animal feed: Environmental assessment and sensitivity analysis to guide future prospects. J. Clean. Prod. 2018, 170, 1260–1267. [Google Scholar] [CrossRef]
- Halloran, A.; Roos, N.; Eilenberg, J.; Cerutti, A.; Bruun, S. Life cycle assessment of edible insects for food protein: A review. Agron. Sustain. Dev. 2016, 36, 57. [Google Scholar] [CrossRef]
- Miglietta, P.P.; De Leo, F.; Ruberti, M.; Massari, S. Mealworms for food: A water footprint perspective. Water 2015, 7, 6190–6203. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Headey, D.; Rue, C.; Thomas, T. Food Systems for Human and Planetary Health: Economic Perspectives and Challenges. Annu. Rev. Resour. Econ. 2021, 13, 131–156. [Google Scholar] [CrossRef]
- Gill, S.R.; Benatar, S.R. Reflections on the political economy of planetary health. Rev. Int. Politi. Econ. 2020, 27, 167–190. [Google Scholar] [CrossRef]
- Gasco, L.; Józefiak, A.; Henry, M. Beyond the protein concept: Health aspects of using edible insects on animals. J. Insects Food Feed 2021, 7, 715–741. [Google Scholar] [CrossRef]
- Ozcan, L.; Tabas, I. Role of endoplasmic reticulum stress in metabolic disease and other disorders. Annu. Rev. Med. 2012, 63, 317–328. [Google Scholar] [CrossRef]
- Yoshida, T.; Yoshida, J. Simultaneous analytical method for urinary metabolites of organophosphorus compounds and moth repellents in general population. J. Chromatogr. B Anal. Technol. Biomed Life Sci. 2012, 880, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals Fed Insect-Based Diets: State-of-the-Art on Digestibility, Performance and Product Quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.; Gavin-Smith, B.; Ferraboschi, C.; Kraemer, K. Food fortification: The advantages, disadvantages and lessons from sight and life programs. Nutrients 2021, 13, 1118. [Google Scholar] [CrossRef]
- Owino, V.O.; Skau, J.; Omollo, S.; Konyole, S.; Kinyuru, J.; Estambale, B.; Owuor, B.; Nanna, R.; Friis, H. WinFood data from Kenya and Cambodia: Constraints on field procedures. Food Nutr. Bull. 2015, 36, S41–S46. [Google Scholar] [CrossRef]
- Moruzzo, R.; Mancini, S.; Guidi, A. Edible Insects and Sustainable Development Goals. Insects 2021, 12, 557. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Kinyuru, J.N.; Kenji, G.M.; Njoroge, S.M.; Ayieko, M. Effect of processing methods on the in vitro protein digestibility and vitamin content of edible winged termite (Macrotermes subhylanus) and grasshopper (Ruspolia differens). Food Bioprocess Technol. 2010, 3, 778–782. [Google Scholar] [CrossRef]
- Hixson, S.M. Fish nutrition and current issues in aquaculture: The balance in providing safe and nutritious seafood, in an environmentally sustainable manner. J. Aquac. Res. Dev. 2014, 5. [Google Scholar] [CrossRef]
- Parodi, A.; Leip, A.; De Boer, I.J.M.; Slegers, P.M.; Ziegler, F.; Temme, E.H.M.; Herrero, M.; Tuomisto, H.L.; Valin, H.; Van Middelaar, C.E.; et al. The potential of future foods for sustainable and healthy diets. Nat. Sustain. 2018, 1, 782–789. [Google Scholar] [CrossRef]
- Woolf, P.J.; Fu, L.L.; Basu, A. vProtein: Identifying optimal amino acid complements from plant-based foods. PLoS ONE 2011, 6, e18836. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, D.; Yang, G.; Bae, D.-K.; Yang, Y.-H.; Kim, T.K.; Kim, D.; Kyung, J.; Yeon, S.; Koo, K.C.; et al. Silk and silkworm pupa peptides suppress adipogenesis in preadipocytes and fat accumulation in rats fed a high-fat diet. Eur. J. Nutr. 2012, 51, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-I converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.; Sagis, L.M.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Dobermann, D.; Swift, J.A.; Field, L.M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 2017, 42, 293–308. [Google Scholar] [CrossRef]
- Becker, N.S.; Margos, G.; Blum, H.; Krebs, S.; Graf, A.; Lane, R.S.; Castillo-Ramírez, S.; Sing, A.; Fingerle, V. Recurrent evolution of host and vector association in bacteria of the Borrelia burgdorferi sensu lato species complex. BMC Genom. 2016, 17, 734. [Google Scholar] [CrossRef]
- Tang, J.E.; Perco, J.G.; Moore, D.; Wilkinson, S.B.; Phillips, S. Resistance training alters the response of fed state mixed muscle protein synthesis in young men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R172–R178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candow, D.G.; Chilibeck, P.D.; Facci, M.; Abeysekara, S.; Zello, G.A. Protein supplementation before and after resistance training in older men. Eur. J. Appl. Physiol. 2006, 97, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.I.; Kim, I.Y.; Wolfe, R.R. Protein consumption and the elderly: What is the optimal level of intake? Nutrients 2016, 8, 359. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.C.; Miller, A.C.; Miller, M.E.; Xiao, H.; Wu, X. Potential health benefits of edible insects. Crit. Rev. Food Sci. Nutr. 2020, 62, 3499–3508. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- De Gier, S.; Verhoeckx, K. Insect (food) allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef]
- Barennes, H.; Phimmasane, M.; Rajaonarivo, C. Insect consumption to address undernutrition, a national survey on the prevalence of insect consumption among adults and vendors in Laos. PLoS ONE 2015, 10, e0136458. [Google Scholar] [CrossRef]
- Smetana, S.; Leonhardt, L.; Kauppi, S.-M.; Pajic, A.; Heinz, V. Insect margarine: Processing, sustainability and design. J. Clean. Prod. 2020, 264, 121670. [Google Scholar] [CrossRef]
- Tello, A.; Aganovic, K.; Parniakov, O.; Carter, A.; Heinz, V.; Smetana, S. Product development and environmental impact of an insect-based milk alternative. Future Foods 2021, 4, 100080. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Megido, R.C.; Alabi, T.; Nieus, C.; Blecker, C.; Danthine, S.; Bogaert, J.; Haubruge, A.; Francis, F. Optimisation of a cheap and residential small-scale production of edible crickets with local by-products as an alternative protein-rich human food source in Ratanakiri Province, Cambodia. J. Sci. Food Agric. 2016, 96, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Spiertz, H. Food production, crops and sustainability: Restoring confidence in science and technology. Curr. Opin. Environ. Sustain. 2010, 2, 439–443. [Google Scholar] [CrossRef]
- Abbasi, T.; Abbasi, S. Biomass energy and the environmental impacts associated with its production and utilization. Renew. Sustain. Energy Rev. 2010, 14, 919–937. [Google Scholar] [CrossRef]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59, 102272. [Google Scholar] [CrossRef]
- Springmann, M.; Clark, M.; Mason-D’Croz, D.; Wiebe, K.; Bodirsky, B.L.; Lassaletta, L.; de Vries, W.; Vermeulen, S.J.; Herrero, M.; Carlson, K.M.; et al. Options for keeping the food system within environmental limits. Nature 2018, 562, 519–525. [Google Scholar] [CrossRef]
- Yen, A.L. Entomophagy and insect conservation: Some thoughts for digestion. J. Insect Conserv. 2009, 13, 667–670. [Google Scholar] [CrossRef]
- Birch, A.N.E.; Begg, G.S.; Squire, G.R. How agro-ecological research helps to address food security issues under new IPM and pesticide reduction policies for global crop production systems. J. Exp. Bot. 2011, 62, 3251–3261. [Google Scholar] [CrossRef]
- De Vries, M.; de Boer, I.J. Comparing environmental impacts for livestock products: A review of life cycle assessments. Livest. Sci. 2010, 128, 1–11. [Google Scholar] [CrossRef]
- Flachowsky, G.; Meyer, U.; Südekum, K.-H. Land Use for Edible Protein of Animal Origin-A Review. Animals 2017, 7, 25. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C. Lekking behavior of the black soldier fly (Diptera: Stratiomyidae). Fla. Entomol. 2001, 84, 729. [Google Scholar] [CrossRef]
- Van Raamsdonk, L.W.D.; Van Der Fels-Klerx, H.J.; De Jong, J. New feed ingredients: The insect opportunity. Food Addit. Contam. Part A 2017, 34, 1384–1397. [Google Scholar] [CrossRef]
- Smith, L.G.; Kirk, G.J.D.; Jones, P.J.; Williams, A.G. The greenhouse gas impacts of converting food production in England and Wales to organic methods. Nat. Commun. 2019, 10, 4641. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste management through composting: Challenges and potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
- Béné, C.; Prager, S.D.; Achicanoy, H.A.; Toro, P.A.; Lamotte, L.; Bonilla, C.; Mapes, B.R. Global map and indicators of food system sustainability. Sci. Data 2019, 6, 279. [Google Scholar] [CrossRef] [PubMed]
- Jensen, N.H.; Lieberoth, A. We will eat disgusting foods together–Evidence of the normative basis of Western entomophagy-disgust from an insect tasting. Food Qual. Prefer. 2019, 72, 109–115. [Google Scholar] [CrossRef]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology; John Wiley & Sons: New York, NY, USA, 2014. [Google Scholar]
- Verbeke, W.; Spranghers, T.; De Clercq, P.; De Smet, S.; Sas, B.; Eeckhout, M. Insects in animal feed: Acceptance and its determinants among farmers, agriculture sector stakeholders and citizens. Anim. Feed Sci. Technol. 2015, 204, 72–87. [Google Scholar] [CrossRef]
- Melgar-Lalanne, G.; Hernández-Álvarez, A.-J.; Salinas-Castro, A. Edible insects processing: Traditional and innovative technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1166–1191. [Google Scholar] [CrossRef]
- Onwezen, M.C.; Bouwman, E.P.; Reinders, M.J.; Dagevos, H. A systematic review on consumer acceptance of alternative proteins: Pulses, algae, insects, plant-based meat alternatives, and cultured meat. Appetite 2021, 159, 105058. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Animal Studies | Selection Bias | Performance Bias | Detection Bias | Attrition Bias | Reporting Bias | Others |
|---|---|---|---|---|---|---|
| Kim et al. [32] | - | - | - | - | - | - |
| Seo et al. [33] | - | - | - | - | ? | ? |
| Bergmans et al. [34] | ? | - | - | - | - | - |
| Dabbou et al. [35] | - | - | - | - | - | ? |
| Bovera et al. [36] | - | - | - | - | - | ? |
| Biasato et al. [37] | - | - | - | - | - | - |
| Gasco et al. [38] | - | - | - | - | - | - |
| Agbemafle et al. [39] | - | - | - | - | - | - |
| Pessina et al. [40] | - | - | - | - | - | ? |
| Ahn et al. [41] | - | - | - | - | - | ? |
| Ahn et al. [42] | - | - | - | - | - | ? |
| Human Studies | Selection Bias | Performance Bias | Detection Bias | Attrition Bias | Reporting Bias | Others |
| Skau et al. [43] | - | - | - | ? | - | - |
| Bauserman et al. [44] | - | - | - | ? | - | - |
| Nirmala et al. [45] | - | ? | + | ? | - | ? |
| Stull et al. [46] | - | - | - | - | - | - |
| Vangsoe et al. [47] | - | - | - | - | - | - |
| Vangsoe et al. [48] | - | ? | - | - | - | - |
| Author, Year, Country | Type of Animal, Sample Size (Male/Female), Age | Duration (Days) | Insect | Intervention (n) | Insect Inclusion Level of CFP (g/100 g Expressed in %) | Variables/Outcomes | Results |
|---|---|---|---|---|---|---|---|
| Kim et al., 2016 (Korea) [32] | C57BL/6J mice 40 (40/-); 7 weeks | 56 | Allomyrina dichotoma larvae | All groups (a–e) start with: 8 weeks (diet-induced obesity): HFD, 60% fat (obese mice). (a) HFD 60%+ 1 μL of 20% DMSO 1 (n = 10) (b) HFD 60% + 1 μL of ALLD 1 (10 mg/mL) (n = 10) (c) LFD 10% (d) LFD, 10% + 1 μL of 20% DMSO 1 (n = 10) (e) LFD, 10% + 1 μL of ALLD 1 (10 mg/mL) (n = 10) | (b) 1% 2 | Appetite control (food intake and body weight) Metabolic traits (inflammatory indicators) | Food intake and body weight were reduced (b,e) compared to (a,b) respectively (S). ADE resulted in strong reduction of ER stress compared to (a) (S). |
| Ahn et al., 2016 (Korea) [42] | Wistar rats, 50 (50/-), 14 weeks | 30 | Gryl- lus bimaculatus | (a) Control + HFD (n = 10) (b) GbG5 + HFD (n = 10) (c) GbG10 + HFD (n = 10) (d) Pravastatin + HFD (n = 10) (e) Chitosan + HFD (n = 10) | (b) 0.0005 GbG (c) 0.001 GbG | Metabolic traits (blood parameters and blood pressure) | Weight of abdominal and epididymal fat, AST, ALT, total cholesterol, and glucose were lower after (b,c) compared to (a) (S). Blood pressure was similar after b,c compared to (a) (NS). Anticoagulant and antithrombotic effects were seen: platelet, thrombin time, prothrombin time and factor I were increased with (b,c) treatment (S). CRP levels of (b,c) decreased compared to (a) (S). |
| Seo et al., 2017 (Korea) [33] | BALB/c mice, 35 (35/-), 5 weeks | 42 | Tenebrio molitor larvae | (a) ND (10% fat) (n = 7) (b) HFD (60% fat) (n = 7) 3 (c) HFD (60% fat) with TML (n = 7) 3 (d) HFD (60% fat) with TML (n = 7) 3 (e) HFD (60% fat) with 3000 mg/kg of yerba mate (n = 7) 3 | (c) 0.01% (d) 0.30% | Metabolic traits (weight gain, fat mass, hepatic steatosis, blood parameters) | Body weight gain, epididymal white adipose tissue size and volume decreased after (c,d) compared to (b) (S). Mean adipocyte volume was reduced after (d) compared to (b) (S) Hepatic lipid droplets, plasma ALT and AST levels, visceral fat were reduced after (c,d) compared to (b) (NS). |
| Dabbou et al., 2018 (Italy) [35] | Ross 308 CD1-IGS broiler chicken, 256 (256/-) ND | 35 | Hermetia Illucens larvae | (a) HI0 (n = 64) (b) HI5 (n = 64) (c) HI10 (n = 64) (d) HI15 (n = 64) | (a) 0% (b) 5% (c) 10% (d) 15% | Growth performance (weight gain, feed intake) Metabolic traits (blood parameters, inflammatory indicators) Intestinal morphology | Dietary HI inclusion (b–d) positively influenced growth performance up to 10%, in terms of improved live weight and daily feed intake during the starter period (S). At 10, 24, and 35 days of age, live weight showed a linear and quadratic response to HI meal with a maximum observed for (c) (S). HI showed a linear response (p = 0.002) to increases up to d) for blood or serum glutathione peroxidase (NS). Intestinal villus height was lower, crypt depth was greater, and villus height-to-crypt depth ratio was lower after (d) compared to (a–c) (S). |
| Bovera et al., 2018 (Italy) [36] | Hy-line Brown hens, 162 (-/162), 16 weeks | 140 | Hermetia Illucens larvae | (a) Control group: corn-soybean meal-based diet (n = 54) (b) HI25(n = 54) (c) HI50 (n = 54) | (b) 7.3% (c) 14.6% | Metabolic traits (blood parameters) Crude protein digestibility | Serum cholesterol and triglyceride levels were reduced after (b,c) compared to (a) (S). Serum globulin levels were higher after (c) compared to (a,b) (S). Crude protein digestibility was the highest (p < 0.05) in (a), followed by (b,c) (NS). |
| Biasato et al., 2018 (Italy) [37] | Label Hubbard hybrid Chickens, 140 (-/140), 43 days | 140 | Tenebrio molitor larvae | (a) Control group: corn-soybean-gluten meal-based diet (n = 70) (b) TM 7.5 (n = 70) | (b) 7.5% | Intestinal morphology | Small intestine revealed similar villus height, crypt depth, and villus height crypt depth ratio between (a,b) (NS). |
| Gasco et al., 2019 (Italy) [38] | Crossbred rabbits, 200 (ND), 36 days | 41 | Hermetia Illucens/Tenebrio molitor larvae | (a) Control group: 1.5% soy-bean oil (n = 40) (b) H50 (n = 40) (c) H100 (n = 40) (d) T50 (n = 40) (e) T100 (n = 40) | (b) 0.75% (c) 1.5% (d) 0.75% (e) 1.50% | Growth performance (feed intake and body weight) Metabolic traits (blood parameters) Crude protein digestibility Intestinal morphology | Weight gain and feed intake was affected similarly after (a–e) (NS). Including insect lipids in rabbit diets did not influence AST, ALT, or ALP enzyme activities. Blood variables were affected similarly after (a–e) (NS). Crude protein digestibility was affected similarly after (a–e) (NS). Villi height, crypt depth, and their ratio were affected similarly after (b–e) compared to (a) (NS). |
| Agbemafle et al., 2019 (Ghana) [39] | Sprague–Dawley rats, 66 (66/-), 21 days | 35 | Acheta domesticus/Rhynchophorus phoenicis fabricius | (a) Normal rats + Casein + ferrous sulfate (n = 8) (b) MD 4 (5% protein) + low protein -Fe (n = 8)-negative control (c) MD 4 (5% protein) + S.torvum (26.7) (n = 8) (d) MD 4 (5% protein) + AD + S. torvum (n = 8) (e) MD 4 (5% protein) + Protein Fe sufficient (n = 8)-positive control (f) MD 4 (5% protein) + AD (n = 8) (g) MD4 (5% protein) + RF (n = 8) | (d) 15.4% (f) 28.3% | Growth performance (body weight recovery, fat mass) Metabolic traits (blood parameters) | After malnourished treatment, weight gain, bone mineral content and lean and fat mass increased similarly after (d,f,g) compared to (e) (NS). Hb increased after (f,g) compared to (a) (NS). |
| Lokman et al., 2019 (Malaysia) [49] | Cobb500 broiler chickens, 100 (150/-), 150 days | 42 | Gryllodes sigillatus | (a) Control: Baseline diet (n = 30) (b) Baseline diet + 0.5 g/kg cricket chitin (n = 30) (c) Baseline diet + 0.5 g/kg cricket chitosan (n = 30) (d) Baseline diet + 0.5 g/kg shrimp chitin (n = 30) (e) Baseline diet + 0.5 g/kg shrimp chitosan (n = 30) | (b) 0.05% chitin (c) 0.05% chitosan | Growth performance (body weight, feed intake and fat mass) | Body weight and feed intake improved after (b) compared to (c) (S). Body weight of a) accumulated more fat compared (b–e) (S). |
| Bergmans et al., 2020 (USA) [34] | Mice, 65 (65/-), 3 weeks | 66 | Gryllodes sigillatus | (a) Control group: Standard adult diet 2018 (n = 10–12) 5 (b) HD+Cricket-based diet (n = 10–12) 5 (c) HD +Milk-based diet, (n = 10–12) 5 (d) HD+Peanut-based diet (n = 10–12) 5 | Growth performance (body weight recovery) Metabolic traits (blood parameters) | After malnourished treatment and recovery diets, there was an increment weight (34%) after (b) compared to (a) (NS). Triglycerides were reduced (47%) after (b) compared to (a) (S). After six weeks on recovery protein diets, there were no differences in the splenetic expression of select inflammatory genes among (a–c) (NS). | |
| Pessina et al., 2020 (Brazil) [40] | Spontaneously hypertensive rats (SHR) 24 (24/-) and age-matched WKY rats (controls) 18 (18/-), 9 weeks | 28 | Tenebrio molitor larvae | (a) SHR SD (n = 8) (b) SHR SD + TM (n = 8) (c) SHR SD + captopril (n = 8) (d) WKY SD (n = 6) (e) WKY SD + TM (n = 6) (f) WKY SD + captopril (n = 6) | (b) 0.29% (e) 0.29% | Metabolic traits (blood parameters, blood pressure and inflammatory indicators) | Systolic BP, heart rate, and coronary perfusion pressure were reduced after (b,c) compared to (a) (S). Rat brain slices of SHR were more resistant to oxidative stress and contained lower levels of inflammatory cytokines, with no effect on vascular and liver enzyme activities (S). |
| Ahn et al., 2020 (Korea) [41] | BKS.Cg-m+/+Leprdb, heterozygous (DB-Hetero, normal) (db/+) male mice (11/-), 12 weeks and homozygous (DB-Homo, diabetes) (db/db) male db mice, 33 (33/-), 12 weeks | 30 | Gryllus bimaculatus- tus | (a) Normal Hetero (DB-Hetero) (n = 11) (b) Control Homo (DB-Homo) (n = 11)-negative control (c) DBHomo + 5 mg/kg treatment of CaG (CaG5) (n = 11) (d) DB Homo + 5 mg/kg treatment of GbG (GbG5) (n = 11) (e) DBHomo + 10 mg/kg treatment of metformin (n = 11)-positive control | (d) 0.0005% GbG | Metabolic traits (blood parameters, and antioxidant activity) | Capacity to reduce glucose, ALT, AST, ALP, LDL-cholesterol and BUN levels increased after (d) compared to (b) (S). Antioxidant activities (catalase, SOD and GPX) increased after (d) compared to (b) (S). |
| Author, Year, Country | Type of Study | Subjects, Sample Size (Male/Female), Age | Duration (Days) | Insect | Intervention | Daily Food Portion of Intervention with Insects | Insect Inclusion Level of CFP (g/100 g Expressed in %) | Insect Inclusion of CFP (Expressed in g) for Each Age Group | Protein Inclusion Level of CFP: g/100 g (Expressed in %) | Protein of CFP Per Day (Expressed in g). | Variables/Outcomes | Results |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Skau et al., 2015 (Cambodia) [43] | Randomized, single-blinded trial | Infants, 419 (220/119), 6 months | 270 | Haplopelma species | (a) WF: Rice-based 1 CFP with small fish and edible spiders (n = 106) (b) WF-L 1: Rice-based CFP with small fish (n = 104) (c) CSB++ 1: Fortified corn-soy blend product (n = 103) (d) CSB+ 1: Fortified whole-soy (n = 106) | 1. Infants 6–8 months: 50 g. 2. Infants 9–11 months: 75 g. 3. Infants 12–15 months: 125 g. | a1,a2,a3 = 1.8% | (a.1) 0.9 g (a.2) 1.35 g (a.3) 2.25 g | (a)15.4% (b)12.6% (c)16.8% (d)14.6% | Growth performance (food intake, body weight) Metabolic traits (blood parameters) | Total weight increases in (a), (b) compared to vs c) (NS). Similar growth observing no differences between (a–d) groups (NS) FFM no differences were observed between (a,b) (NS). Plasma ferritin, sTfR, and hemoglobin concentration no differences were observed between (a–d) (NS). Total weight increase in (a,b) compared to (c) (NS). | |
| Bauserman et al., 2015 (Democratic Republic of Congo) [44] | Cluster- randomized controlled trial | Infants, 222 (113/109), 6 months | 540 | Caterpillar | (a) Usual diet 2 (n = 110) (b) Caterpillar 2 cereal. (n = 110) | Infants 6–12 months of age: 30 g Infants 12–18 months: 45 g | (b.1) 6.9 (b.2) 10.3 | Growth performance (body weight recovery) Metabolic traits (blood parameters) | Stunting prevalence, no differences were observed between (a,b) (NS). Fe: no differences were observed between (a,b) (NS). Hb increased in (b) compared to (a) anemia decreased in (b) compared to (a) (S). | |||
| Nirmala et al., 2017 (Indonesia) [45] | Non-randomized controlled trial | Infants, 23 (12/11), 1–5 years | 45 | Rhynchophorus ferrugineus | (a) Usual diet (n = 10) (b) Sago worm inclusive diet (n = 13) | 2 pieces of 50 g | (b) 9.70% | (a) 3.9 ±1.7 (b) 5.9 ± 1.7 | Growth performance (body weight) | Weight and height no changes were observed between a) and b) (NS) | ||
| Stull et al., 2018 (USA) [46] | Double-blinded randomized crossover trial | Healthy adults, 20 (9/11), 18–65 years | 14 | Gryllodes sigillatus | (a) Control breakfast meal (n = 10) (b) Cricket breakfast meal (n = 10) | Shake + pumpkin muffin (160 g) | (b) 14.9% Shake; 9.37% Muffin | (b) 14.78% | (a) 9 (b) 21.67 | Gut microbiome composition Metabolic traits (inflammatory indicators) | Bifidobacterium animalis increased 5.7 more in (b) compared to (a) (S) Plasma TNF-α decreased b) compared to (a) (S). | |
| Vangsoe et al. A 2018 (Denmark) [47] | Randomized, controlled, single-blinded trial | Healthy young adults, 18(18/-), 18–30 years | 56 | Alphitobius diaperinus | (a) Isocaloric carbohydrate bar (n = 9) (b) Insect protein bar (n = 9) | 2 bars a day | (b) 0.04% 3 | (a) 7.2 (b) 8 | Changes in muscle mass composition and strength | Morphological adaptations such as hypertrophy or muscle strength show no changes in (a) compared to (b) (NS). | ||
| Vangsoe et al. B 2018 (Denmark) [48] | Randomized, cross-over study | Healthy young adults,6 (6/-), 18–30 years | 1 | Alphitobius diaperinus | (a) Drink placebo (water) (b) Drink whey isolate (c) Drink soy isolate (d) Drink insect isolate | 400 mL per day | (d) 7.6% 4 | (d) 30.5 g isolate powder | (b) 25 g (c) 25 g (d) 25 g | Crude protein digestibility | Blood concentrations of EAA, BCAA and leucine increased in (b–d) compared to (a) over a 120 min period (S). Slowly digested (d) compared to (b,c) (S). |
| Animal Feed Consumption (kg Edible Protein) | Human Food Consumption (kg De Protein) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Waste-Feed Insects | Non Waste Insects | ||||||||||||
| Insect | Author, Year, Country | Land Use (m2) | GHG (Kg CO2 eq) | Energy Use (MJ) | Author, Year, Country | Land Use (m2) | GHG (Kg CO2 eq) | Energy Use (MJ) | Author, Year, Country | Water Footprint (m3) | Land Use (m2) | GHG (Kg CO2 eq) | Energy Use (MJ) |
| Tenebrio molitor larvae | Thévenot et al., 2018 (France) F,J [53] | 6.35 | 5.77 | 217.37 | Oonincx et al., 2012 (USA) F [7] | 17.68 | 13.16 | 167.23 | |||||
| Miglietta et al., 2015 (Italy) F,I [55] | 23 | ||||||||||||
| Musca domestica larvae | Van Zanten et al., 2015 (Netherlands) A,J [50] | 0.07 | 1.43 | 18.98 | |||||||||
| Hermetia illucens larvae | Salomone et al., 2016 (Italy) B,I [51] | 0.05 | 2.1 | 15.1 | |||||||||
| Muys et al. 2014 (UK) C,I [52] | 0.06 D 0.19 E | 2.1 | 15.1 | ||||||||||
| Acheta domesticus | Halloran et al., 2017 (Denmark) F,I [54] | 3.97 G 2.63 H | |||||||||||
| Traditional Livestock Animals for Food (kg de Protein) | |||||
|---|---|---|---|---|---|
| Animal | Author, Year, Country | Water Food Print (m3) | Land Use (m2) | GHG (Kg CO2 eq) | Energy Use (MJ) |
| Pork | Vries and de Boer. 2010 (Netherlands) F [97] | 47–64 | 21–53 | 95–236 | |
| Miglietta et al. 2015 (Italy) F [55] | 57 | ||||
| Chicken | Vries and de Boer. 2010 (Netherlands) F [97] | 42–52 | 18–36 | 80–152 | |
| Miglietta et al. 2015 (Italy) F [55] | 34 | ||||
| Beef | Vries and de Boer. 2010 (Netherlands) F [97] | 144–258 | 75–170 | 177–273 | |
| Miglietta et al. 2015 (Italy) F [55] | 112 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ros-Baró, M.; Casas-Agustench, P.; Díaz-Rizzolo, D.A.; Batlle-Bayer, L.; Adrià-Acosta, F.; Aguilar-Martínez, A.; Medina, F.-X.; Pujolà, M.; Bach-Faig, A. Edible Insect Consumption for Human and Planetary Health: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 11653. https://doi.org/10.3390/ijerph191811653
Ros-Baró M, Casas-Agustench P, Díaz-Rizzolo DA, Batlle-Bayer L, Adrià-Acosta F, Aguilar-Martínez A, Medina F-X, Pujolà M, Bach-Faig A. Edible Insect Consumption for Human and Planetary Health: A Systematic Review. International Journal of Environmental Research and Public Health. 2022; 19(18):11653. https://doi.org/10.3390/ijerph191811653
Chicago/Turabian StyleRos-Baró, Marta, Patricia Casas-Agustench, Diana Alícia Díaz-Rizzolo, Laura Batlle-Bayer, Ferran Adrià-Acosta, Alícia Aguilar-Martínez, Francesc-Xavier Medina, Montserrat Pujolà, and Anna Bach-Faig. 2022. "Edible Insect Consumption for Human and Planetary Health: A Systematic Review" International Journal of Environmental Research and Public Health 19, no. 18: 11653. https://doi.org/10.3390/ijerph191811653
APA StyleRos-Baró, M., Casas-Agustench, P., Díaz-Rizzolo, D. A., Batlle-Bayer, L., Adrià-Acosta, F., Aguilar-Martínez, A., Medina, F. -X., Pujolà, M., & Bach-Faig, A. (2022). Edible Insect Consumption for Human and Planetary Health: A Systematic Review. International Journal of Environmental Research and Public Health, 19(18), 11653. https://doi.org/10.3390/ijerph191811653