
Longitudinal Impact of WTC Dust Inhalation on Rat Cardiac Tissue Transcriptomic Profiles

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. WTC Dust and Generation for Exposures
2.3. Experimental Design
2.4. Data Analysis: Overview
2.5. RNA-seq Analysis
2.6. Differential Gene Expression Analysis
2.7. Statistical Analysis
3. Results
3.1. Cross-Sectional Analyses of Differentially Expressed Genes (DEGs) at Days 1, 30, 240 and 360
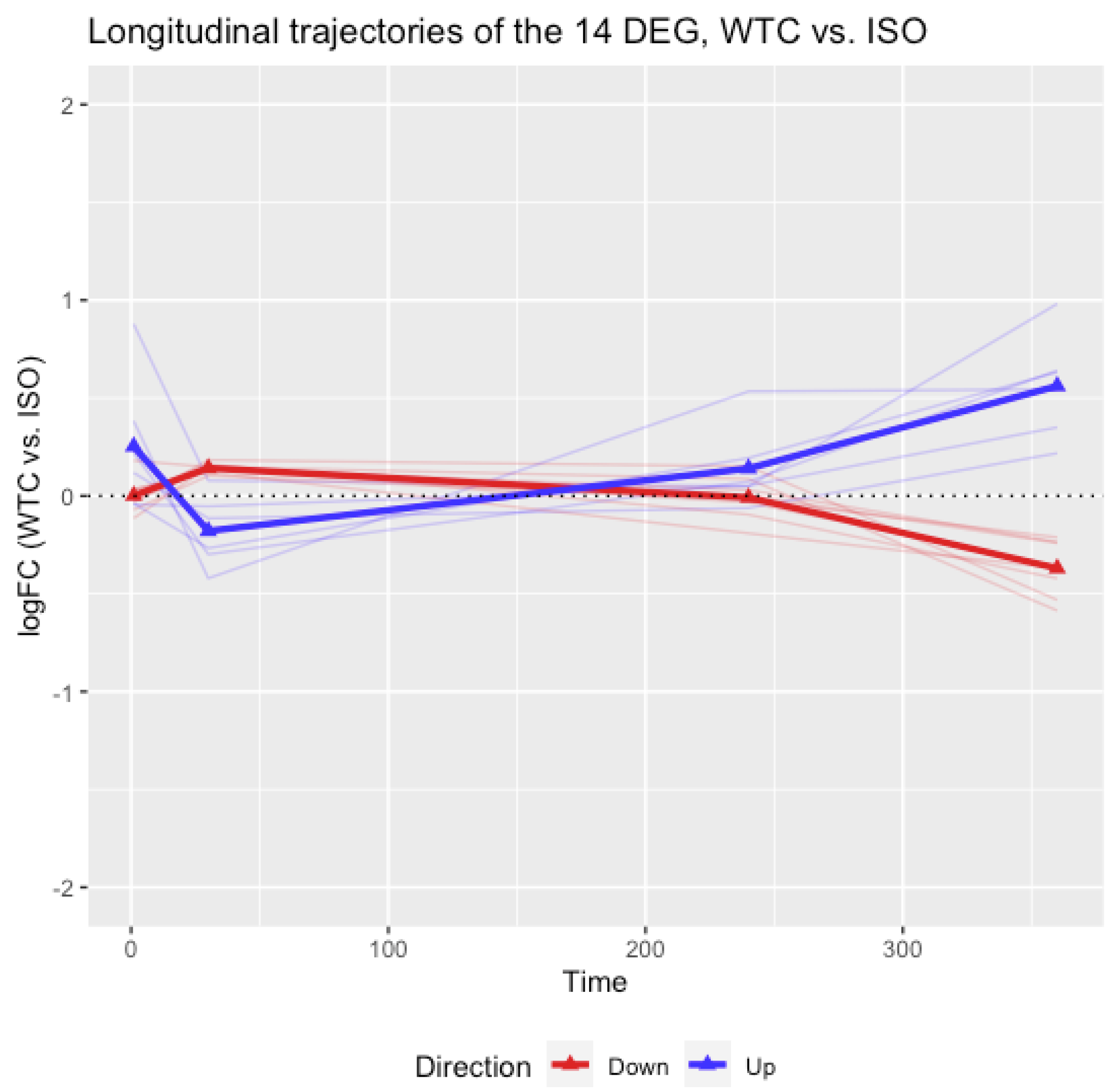
3.2. Longitudinal Analyses of the 14 Selected DEGs
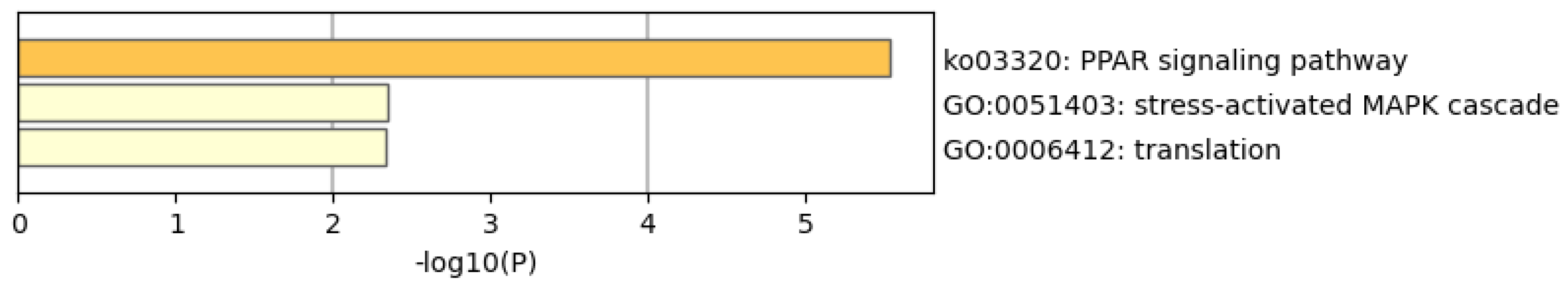
3.3. Potential WTC Effects in the Presence of ISO-WTC Dust Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lippmann, M.; Cohen, M.D.; Chen, L.C. Health effects of World Trade Center (WTC) Dust: An unprecedented disaster’s inadequate risk management. Crit. Rev. Toxicol. 2015, 45, 492–530. [Google Scholar] [CrossRef]
- McGee, J.K.; Chen, L.C.; Cohen, M.D.; Chee, G.R.; Prophete, C.M.; Haykal-Coates, N.; Wasson, S.J.; Conner, T.L.; Costa, D.L.; Gavett, S.H. Chemical analysis of World Trade Center fine particulate matter for use in toxicologic assessment. Environ. Health Perspect. 2003, 111, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.W.; Zeig-Owens, R.; Joe, C.; Hall, C.B.; Webber, M.P.; Weiden, M.D.; Cleven, K.L.; Jaber, N.; Skerker, M.; Yip, J.; et al. Long-term Cardiovascular Disease Risk Among Firefighters After the World Trade Center Disaster. JAMA Netw. Open 2019, 2, e199775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, H.T.; Miller-Archie, S.A.; Cone, J.E.; Morabia, A.; Stellman, S.D. Heart disease among adults exposed to the September 11, 2001 World Trade Center disaster: Results from the World Trade Center Health Registry. Prev. Med. 2011, 53, 370–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lioy, P.J.; Weisel, C.P.; Millette, J.R.; Eisenreich, S.; Vallero, D.; Offenberg, J.; Buckley, B.; Turpin, B.; Zhong, M.; Cohen, M.D.; et al. Characterization of the dust/smoke aerosol that settled east of the World Trade Center (WTC) in lower Manhattan after the collapse of the WTC 11 September 2001. Environ. Health Perspect. 2002, 110, 703–714. [Google Scholar] [CrossRef]
- Geyh, A.S.; Chillrud, S.; Williams, D.L.; Herbstman, J.; Symons, J.M.; Rees, K.; Ross, J.; Kim, S.R.; Lim, H.J.; Turpin, B.; et al. Assessing truck driver exposure at the World Trade Center disaster site: Personal and area monitoring for particulate matter and volatile organic compounds during October 2001 and April 2002. J. Occup. Environ. Hyg. 2005, 2, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Gavett, S.H. World Trade Center fine particulate matter-chemistry and toxic respiratory effects: An overview. Environ. Health Perspect. 2003, 111, 971. [Google Scholar] [CrossRef]
- Cohen, M.D. Pulmonary immunotoxicology of select metals: Aluminum, arsenic, cadmium, chromium, copper, manganese, nickel, vanadium, and zinc. J. Immunotoxicol. 2004, 1, 39–69. [Google Scholar] [CrossRef]
- Bui, Q.Q.; Clark, C.R.; Naas, D.J.; Ulrich, C.E.; Elangbam, C.S. A subacute inhalation exposure evaluation of a scrubbing solution used in petroleum refineries. J. Toxicol. Environ. Health A 1998, 54, 49–62. [Google Scholar] [CrossRef]
- Clary-Meinesz, C.; Mouroux, J.; Cosson, J.; Huitorel, P.; Blaive, B. Influence of external pH on ciliary beat frequency in human bronchi and bronchioles. Eur. Respir. J. 1998, 11, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Holma, B.; Lindegren, M.; Andersen, J.M. pH effects on ciliomotility and morphology of respiratory mucosa. Arch. Environ. Health 1977, 32, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.D.; Vaughan, J.M.; Garrett, B.; Prophete, C.; Horton, L.; Sisco, M.; Ghio, A.; Zelikoff, J.; Lung-chi, C. Impact of acute exposure to WTC dust on ciliated and goblet cells in lungs of rats. Inhal. Toxicol. 2015, 27, 354–361. [Google Scholar] [CrossRef]
- Cohen, M.D.; Prophete, C.; Horton, L.; Sisco, M.; Park, S.H.; Lee, H.W.; Zelikoff, J.; Chen, L.C. Impact on rats from acute intratracheal inhalation exposures to WTC dusts. Inhal. Toxicol. 2020, 32, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, J.M.; Pfeffer, M.A.; Fishbein, M.C.; Frohlich, E.D. Cardiac-Function and Morphology with Aging in the Spontaneously Hypertensive Rat. Am. J. Physiol. 1979, 237, H461–H468. [Google Scholar] [CrossRef] [PubMed]
- Konigshofer, P.; Brusilovskaya, K.; Schwabl, P.; Reiberger, T. Animal models of portal hypertension. Mol. Basis Dis. 2019, 1865, 1019–1030. [Google Scholar] [CrossRef]
- Cohen, M.D.; Vaughan, J.M.; Garrett, B.; Prophete, C.; Horton, L.; Sisco, M.; Kodavanti, U.P.; Ward, W.O.; Peltier, R.E.; Zelikoff, J.; et al. Acute high-level exposure to WTC particles alters expression of genes associated with oxidative stress and immune function in the lung. J. Immunotoxicol. 2015, 12, 140–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavett, S.H.; Haykal-Coates, N.; Highfill, J.W.; Ledbetter, A.D.; Chen, L.C.; Cohen, M.D.; Harkema, J.R.; Wagner, J.G.; Costa, D.L. World Trade Center fine particulate matter causes respiratory tract hyperresponsiveness in mice. Environ. Health Perspect. 2003, 111, 981–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, J.M.; Garrett, B.J.; Prophete, C.; Horton, L.; Sisco, M.; Soukup, J.M.; Zelikoff, J.T.; Ghio, A.; Peltier, R.E.; Asgharian, B.; et al. A novel system to generate WTC dust particles for inhalation exposures. J. Expo. Sci. Environ. Epidemiol. 2014, 24, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Lu, Y.; Shao, Y.; Prophete, C.; Horton, L.; Sisco, M.; Lee, H.W.; Kluz, T.; Sun, H.; Costa, M.; et al. Longitudinal impact on rat cardiac tissue transcriptomic profiles due to acute intratracheal inhalation exposures to isoflurane. PLoS ONE 2021, 16, e0257241. [Google Scholar] [CrossRef]
- Lauterstein, D.E.; Tijerina, P.B.; Corbett, K.; Akgol Oksuz, B.; Shen, S.S.; Gordon, T.; Klein, C.B.; Zelikoff, J.T. Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages. Int. J. Environ. Res. Public Health 2016, 13, 417. [Google Scholar] [CrossRef] [Green Version]
- Laulicht, F.; Brocato, J.; Cartularo, L.; Vaughan, J.; Wu, F.; Kluz, T.; Sun, H.; Oksuz, B.A.; Shen, S.; Peana, M.; et al. Tungsten-induced carcinogenesis in human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2015, 288, 33–39. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.A.; Flecknell, P.A. Anaesthesia and post-operative analgesia following experimental surgery in laboratory rodents: Are we making progress? Altern. Lab. Anim. 2005, 33, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Cesarovic, N.; Nicholls, F.; Rettich, A.; Kronen, P.; Hassig, M.; Jirkof, P.; Arras, M. Isoflurane and sevoflurane provide equally effective anaesthesia in laboratory mice. Lab. Anim. 2010, 44, 329–336. [Google Scholar] [CrossRef]
- Rossaint, R.; Reyle-Hahn, M.; Schulte Am Esch, J.; Scholz, J.; Scherpereel, P.; Vallet, B.; Giunta, F.; Del Turco, M.; Erdmann, W.; Tenbrinck, R.; et al. Multicenter ran.ndomized comparison of the efficacy and safety of xenon and isoflurane in patients undergoing elective surgery. Anesthesiology 2003, 98, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Huneke, R.; Fassl, J.; Rossaint, R.; Luckhoff, A. Effects of volatile anesthetics on cardiac ion channels. Acta Anaesthesiol. Scand. 2004, 48, 547–561. [Google Scholar] [CrossRef]
- Uhlig, C.; Bluth, T.; Schwarz, K.; Deckert, S.; Heinrich, L.; De Hert, S.; Landoni, G.; Serpa Neto, A.; Schultz, M.J.; Pelosi, P.; et al. Effects of Volatile Anesthetics on Mortality and Postoperative Pulmonary and Other Complications in Patients Undergoing Surgery: A Systematic Review and Meta-analysis. Anesthesiology 2016, 124, 1230–1245. [Google Scholar] [CrossRef]
- Tarnow, J.; Bruckner, J.B.; Eberlein, H.J.; Hess, W.; Patschke, D.; Weymar, A. Cardiovascular effects of the new inhalation anaesthetic isoflurane in man (author’s transl). Anaesthesist 1975, 24, 425–431. [Google Scholar]
- Stachnik, J. Inhaled anesthetic agents. Am. J. Health Syst. Pharm. 2006, 63, 623–634. [Google Scholar] [CrossRef]
- Constantinides, C.; Mean, R.; Janssen, B.J. Effects of isoflurane anesthesia on the cardiovascular function of the C57BL/6 mouse. ILAR J. 2011, 52, e21–e31. [Google Scholar] [PubMed]
- Wong, D.; Makowska, I.J.; Weary, D.M. Rat aversion to isoflurane versus carbon dioxide. Biol. Lett. 2013, 9, 20121000. [Google Scholar] [CrossRef] [PubMed]
- Van Allen, N.R.; Krafft, P.R.; Leitzke, A.S.; Applegate, R.L., 2nd; Tang, J.; Zhang, J.H. The role of Volatile Anesthetics in Cardioprotection: A systematic review. Med. Gas. Res. 2012, 2, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, P.; Teng, J.; Zou, J.; Fang, Y.; Zhang, X.; Bosnjak, Z.J.; Liang, M.; Ding, X. miR-21 contributes to xenon-conferred amelioration of renal ischemia-reperfusion injury in mice. Anesthesiology 2013, 119, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Lang, X.E.; Wang, X.; Zhang, K.R.; Lv, J.Y.; Jin, J.H.; Li, Q.S. Isoflurane preconditioning confers cardioprotection by activation of ALDH2. PLoS ONE 2013, 8, e52469. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Feng, C.; Li, L.; Zuo, Z. Contribution of microRNA-203 to the isoflurane preconditioning-induced neuroprotection. Brain Res. Bull. 2012, 88, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.M.; Yan, Y.; Bai, X.; Ge, Z.D.; Liang, M.; Kriegel, A.J.; Twaroski, D.M.; Bosnjak, Z.J. Up-regulation of microRNA-21 mediates isoflurane-induced protection of cardiomyocytes. Anesthesiology 2015, 122, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.J.; Liu, B. Inhibition of MicroRNA-23 Contributes to the Isoflurane-Mediated Cardioprotection Against Oxidative Stress. Cardiovasc. Toxicol. 2018, 18, 450–458. [Google Scholar] [CrossRef]
- Qiao, S.; Olson, J.M.; Paterson, M.; Yan, Y.; Zaja, I.; Liu, Y.; Riess, M.L.; Kersten, J.R.; Liang, M.; Warltier, D.C.; et al. MicroRNA-21 Mediates Isoflurane-induced Cardioprotection against Ischemia-Reperfusion Injury via Akt/Nitric Oxide Synthase/Mitochondrial Permeability Transition Pore Pathway. Anesthesiology 2015, 123, 786–798. [Google Scholar] [CrossRef] [Green Version]
- Bakar, A.M.; Park, S.W.; Kim, M.; Lee, H.T. Isoflurane protects against human endothelial cell apoptosis by inducing sphingosine kinase-1 via ERK MAPK. Int. J. Mol. Sci. 2012, 13, 977–993. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Y. Isoflurane reduces endotoxin-induced oxidative, inflammatory, and apoptotic responses in H9c2 cardiomyocytes. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3976–3987. [Google Scholar] [CrossRef]
- Sloan, N.L.; Shapiro, M.Z.; Sabra, A.; Dasaro, C.R.; Crane, M.A.; Harrison, D.J.; Luft, B.J.; Moline, J.M.; Udasin, I.G.; Todd, A.C.; et al. Cardiovascular disease in the World Trade Center Health Program General Responder Cohort. Am. J. Ind. Med. 2021, 64, 97–107. [Google Scholar] [CrossRef]
- Pulliam, D.A.; Deepa, S.S.; Liu, Y.; Hill, S.; Lin, A.L.; Bhattacharya, A.; Shi, Y.; Sloane, L.; Viscomi, C.; Zeviani, M.; et al. Complex IV-deficient Surf1(-/-) mice initiate mitochondrial stress responses. Biochem. J. 2014, 462, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Z.H.; Vittinghoff, E.; Musone, S.L.; Lin, F.; Whiteman, D.; Pawlikowska, L.; Kwok, P.Y.; Olgin, J.E.; Aouizerat, B.E. Association of TGFBR2 polymorphism with risk of sudden cardiac arrest in patients with coronary artery disease. Heart Rhythm. 2009, 6, 1745–1750. [Google Scholar] [CrossRef] [Green Version]
- Robson, A.; Allinson, K.R.; Anderson, R.H.; Henderson, D.J.; Arthur, H.M. The TGFbeta type II receptor plays a critical role in the endothelial cells during cardiac development. Dev. Dyn. 2010, 239, 2435–2442. [Google Scholar] [CrossRef]
- Erdmann, J.; Kessler, T.; Munoz Venegas, L.; Schunkert, H. A decade of genome-wide association studies for coronary artery disease: The challenges ahead. Cardiovasc. Res. 2018, 114, 1241–1257. [Google Scholar] [CrossRef]
- Landry, N.; Kavosh, M.S.; Filomeno, K.L.; Rattan, S.G.; Czubryt, M.P.; Dixon, I.M.C. Ski drives an acute increase in MMP-9 gene expression and release in primary cardiac myofibroblasts. Physiol. Rep. 2018, 6, e13897. [Google Scholar] [CrossRef]
- Xiong, B.; Jha, V.; Min, J.K.; Cho, J. Protein disulfide isomerase in cardiovascular disease. Exp. Mol. Med. 2020, 52, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Schroen, B.; Leenders, J.J.; van Erk, A.; Bertrand, A.T.; van Loon, M.; van Leeuwen, R.E.; Kubben, N.; Duisters, R.F.; Schellings, M.W.; Janssen, B.J.; et al. Lysosomal integral membrane protein 2 is a novel component of the cardiac intercalated disc and vital for load-induced cardiac myocyte hypertrophy. J. Exp. Med. 2007, 204, 1227–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhang, M.; Wu, H.; Ding, X.; Li, D.; Dong, X.; Hu, X.; Su, S.; Shang, W.; Wu, J.; et al. SAIL: A new conserved anti-fibrotic lncRNA in the heart. Basic. Res. Cardiol. 2021, 116, 15. [Google Scholar] [CrossRef] [PubMed]
- Peghaire, C.; Bats, M.L.; Sewduth, R.; Jeanningros, S.; Jaspard, B.; Couffinhal, T.; Duplaa, C.; Dufourcq, P. Fzd7 (Frizzled-7) Expressed by Endothelial Cells Controls Blood Vessel Formation Through Wnt/beta-Catenin Canonical Signaling. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2369–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Elmagd, M.; Mulvaney, J.; Wheeler, G.N. Frizzled-7 is required for Xenopus heart development. Biol. Open 2017, 6, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Combs, M.D.; Yutzey, K.E. Heart valve development: Regulatory networks in development and disease. Circ. Res. 2009, 105, 408–421. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Georges, A.; Tucker, N.R.; Kyryachenko, S.; Toomer, K.; Schott, J.J.; Delling, F.N.; Fernandez-Friera, L.; Solis, J.; Ellinor, P.T.; et al. Genome-Wide Association Study-Driven Gene-Set Analyses, Genetic, and Functional Follow-Up Suggest GLIS1 as a Susceptibility Gene for Mitral Valve Prolapse. Circ. Genom. Precis. Med. 2019, 12, e002497. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Guo, H.; Yang, X.; Zhou, L.; Zhang, X.; Cheng, L.; Zeng, H.; Hu, F.B.; Tanguay, R.M.; Wu, T. Genetic variations in HSPA8 gene associated with coronary heart disease risk in a Chinese population. PLoS ONE 2010, 5, e9684. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, Y.; Urashima, T.; Kawachi, F.; Akaike, T.; Kusakari, Y.; Ida, H.; Minamisawa, S. Pulmonary hypertension due to left heart disease causes intrapulmonary venous arterialization in rats. J. Thorac. Cardiovasc. Surg. 2017, 154, 1742–1753.e8. [Google Scholar] [CrossRef] [Green Version]
- Nagai-Okatani, C.; Minamino, N. Aberrant Glycosylation in the Left Ventricle and Plasma of Rats with Cardiac Hypertrophy and Heart Failure. PLoS ONE 2016, 11, e0150210. [Google Scholar] [CrossRef] [Green Version]
- Finck, B.N. The PPAR regulatory system in cardiac physiology and disease. Cardiovasc. Res. 2007, 73, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Kim, J. Peroxisome Proliferator-Activated Receptors and the Heart: Lessons from the Past and Future Directions. PPAR Res. 2015, 2015, 271983. [Google Scholar] [CrossRef] [Green Version]
- Hibuse, T.; Maeda, N.; Nakatsuji, H.; Tochino, Y.; Fujita, K.; Kihara, S.; Funahashi, T.; Shimomura, I. The heart requires glycerol as an energy substrate through aquaporin 7, a glycerol facilitator. Cardiovasc. Res. 2009, 83, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkerk, A.O.; Lodder, E.M.; Wilders, R. Aquaporin Channels in the Heart-Physiology and Pathophysiology. Int. J. Mol. Sci. 2019, 20, 2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; Boluyt, M.O.; O’Neill, L.; Zheng, J.S.; Wu, G.; Nitta, Y.K.; Crow, M.T.; Lakatta, E.G. Myocardial retinoid X receptor, thyroid hormone receptor, and myosin heavy chain gene expression in the rat during adult aging. J. Gerontol. A Biol. Sci. Med. Sci. 1999, 54, B23–B27. [Google Scholar] [CrossRef] [Green Version]
- Finck, B.N.; Chinetti, G.; Staels, B. PPARs/RXRs in Cardiovascular Physiology and Disease. PPAR Res. 2008, 2008, 173780. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Chang, J.P.; Lin, Y.S.; Pan, K.L.; Ho, W.C.; Liu, W.H.; Chang, T.H.; Huang, Y.K.; Fang, C.Y.; Chen, C.J. Deciphering the gene expression profile of peroxisome proliferator-activated receptor signaling pathway in the left atria of patients with mitral regurgitation. J. Transl. Med. 2016, 14, 157. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Sreenivasan, U.; Gong, D.W.; O’Connell, K.A.; Dabkowski, E.R.; Hecker, P.A.; Ionica, N.; Konig, M.; Mahurkar, A.; Sun, Y.; et al. Cardiomyocyte-specific perilipin 5 overexpression leads to myocardial steatosis and modest cardiac dysfunction. J. Lipid. Res. 2013, 54, 953–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WTC vs. ISO | WTC vs. Naïve | |||||
|---|---|---|---|---|---|---|
| Name | logFC | p | FDR | logFC | p | FDR |
| Surf1 | 0.63 | 9.45 × 10−6 | 1.53 × 10−2 | 0.47 | 2.64 × 10−4 | 3.48 × 10−2 |
| Tgfbr2 | −0.42 | 1.93 × 10−5 | 1.53 × 10−2 | −0.20 | 1.55 × 10−2 | 1.50 × 10−1 |
| Ski | −0.37 | 3.01 × 10−5 | 1.59 × 10−2 | −0.18 | 1.57 × 10−2 | 1.50 × 10−1 |
| Pola2 | 0.35 | 5.54 × 10−5 | 1.99 × 10−2 | 0.22 | 4.08 × 10−3 | 9.65 × 10−2 |
| P4hb | −0.24 | 2.12 × 10−4 | 3.58 × 10−2 | −0.16 | 5.29 × 10−3 | 1.07 × 10−1 |
| Rps18 | 0.54 | 4.28 × 10−4 | 3.96 × 10−2 | 0.75 | 1.36 × 10−5 | 1.08 × 10−2 |
| Mfsd1 | −0.21 | 4.77 × 10−4 | 4.17 × 10−2 | −0.14 | 1.29 × 10−2 | 1.41 × 10−1 |
| Scarb2 | −0.36 | 5.17 × 10−4 | 4.18 × 10−2 | −0.30 | 3.05 × 10−3 | 9.12 × 10−2 |
| Fzd7 | −0.59 | 7.53 × 10−4 | 4.92 × 10−2 | −0.52 | 2.49 × 10−3 | 8.65 × 10−2 |
| Isg20l2 | −0.53 | 8.52 × 10−4 | 5.03 × 10−2 | −0.58 | 3.76 × 10−4 | 4.29 × 10−2 |
| Safb | 0.22 | 9.69 × 10−4 | 5.16 × 10−2 | 0.25 | 2.34 × 10−4 | 3.47 × 10−2 |
| Glis1 | 0.64 | 1.98 × 10−3 | 7.06 × 10−2 | 0.77 | 4.35 × 10−4 | 4.53 × 10−2 |
| Hspa8_2 | 0.98 | 2.02 × 10−3 | 7.06 × 10−2 | 1.96 | 2.61 × 10−6 | 7.07 × 10−3 |
| LOC294154 | −0.24 | 2.07 × 10−3 | 7.19 × 10−2 | −0.30 | 3.24 × 10−4 | 3.83 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-H.; Lu, Y.; Shao, Y.; Prophete, C.; Horton, L.; Sisco, M.; Lee, H.-W.; Kluz, T.; Sun, H.; Costa, M.; et al. Longitudinal Impact of WTC Dust Inhalation on Rat Cardiac Tissue Transcriptomic Profiles. Int. J. Environ. Res. Public Health 2022, 19, 919. https://doi.org/10.3390/ijerph19020919
Park S-H, Lu Y, Shao Y, Prophete C, Horton L, Sisco M, Lee H-W, Kluz T, Sun H, Costa M, et al. Longitudinal Impact of WTC Dust Inhalation on Rat Cardiac Tissue Transcriptomic Profiles. International Journal of Environmental Research and Public Health. 2022; 19(2):919. https://doi.org/10.3390/ijerph19020919
Chicago/Turabian StylePark, Sung-Hyun, Yuting Lu, Yongzhao Shao, Colette Prophete, Lori Horton, Maureen Sisco, Hyun-Wook Lee, Thomas Kluz, Hong Sun, Max Costa, and et al. 2022. "Longitudinal Impact of WTC Dust Inhalation on Rat Cardiac Tissue Transcriptomic Profiles" International Journal of Environmental Research and Public Health 19, no. 2: 919. https://doi.org/10.3390/ijerph19020919
APA StylePark, S. -H., Lu, Y., Shao, Y., Prophete, C., Horton, L., Sisco, M., Lee, H. -W., Kluz, T., Sun, H., Costa, M., Zelikoff, J., Chen, L. -C., Gorr, M. W., Wold, L. E., & Cohen, M. D. (2022). Longitudinal Impact of WTC Dust Inhalation on Rat Cardiac Tissue Transcriptomic Profiles. International Journal of Environmental Research and Public Health, 19(2), 919. https://doi.org/10.3390/ijerph19020919