
Cytotoxicity and Genotoxicity of Senecio vulgaris L. Extracts: An In Vitro Assessment in HepG2 Liver Cells

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents, and Media
2.2. Plant Material
2.3. Extraction Methods
2.3.1. Chloroform Extract
2.3.2. Aqueous Extract
2.4. Cell Line and Culture Conditions
2.5. Cell Count and Viability: AO/DAPI Double Staining
2.6. Genotoxicity Testing: Comet Assay
3. Results
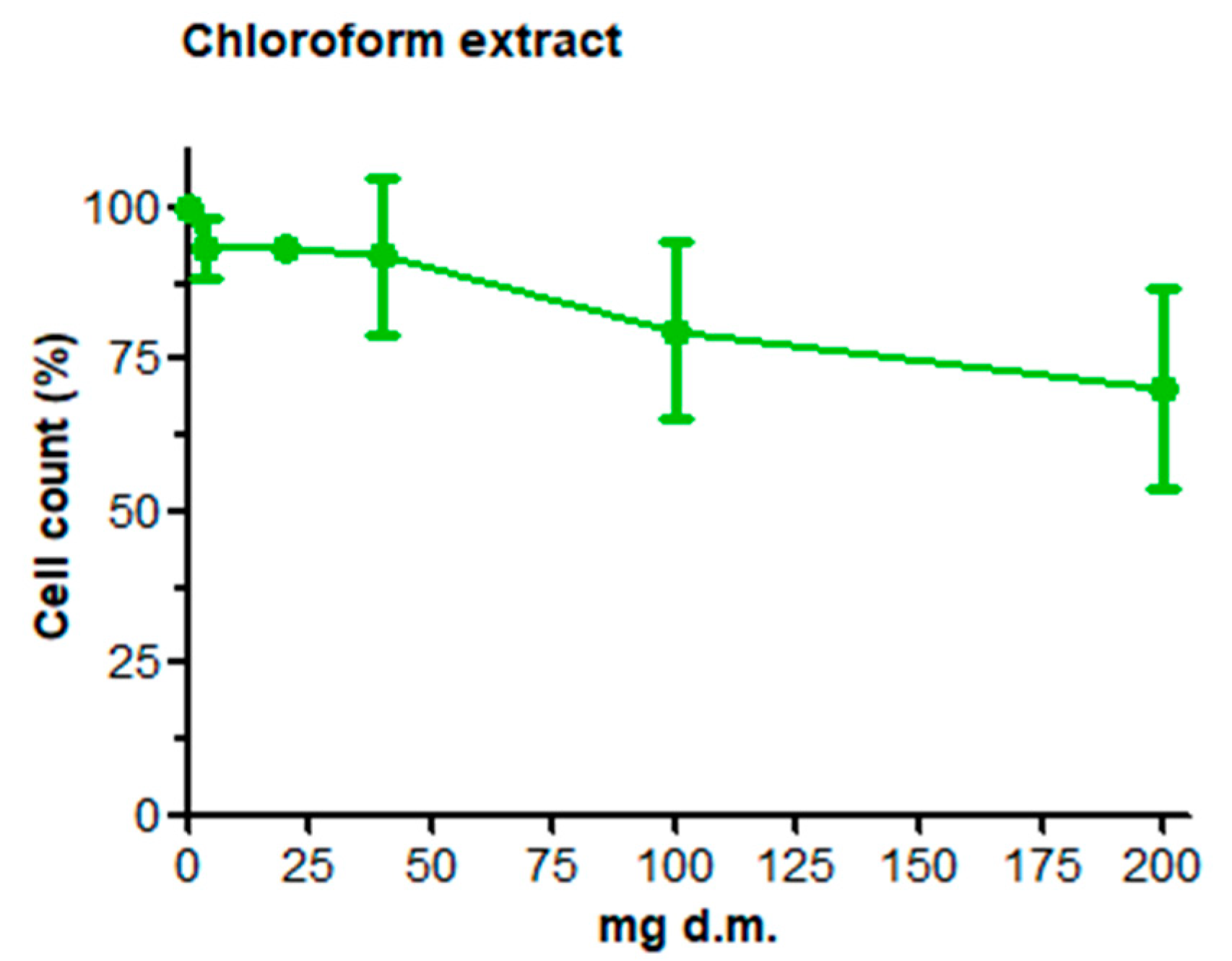
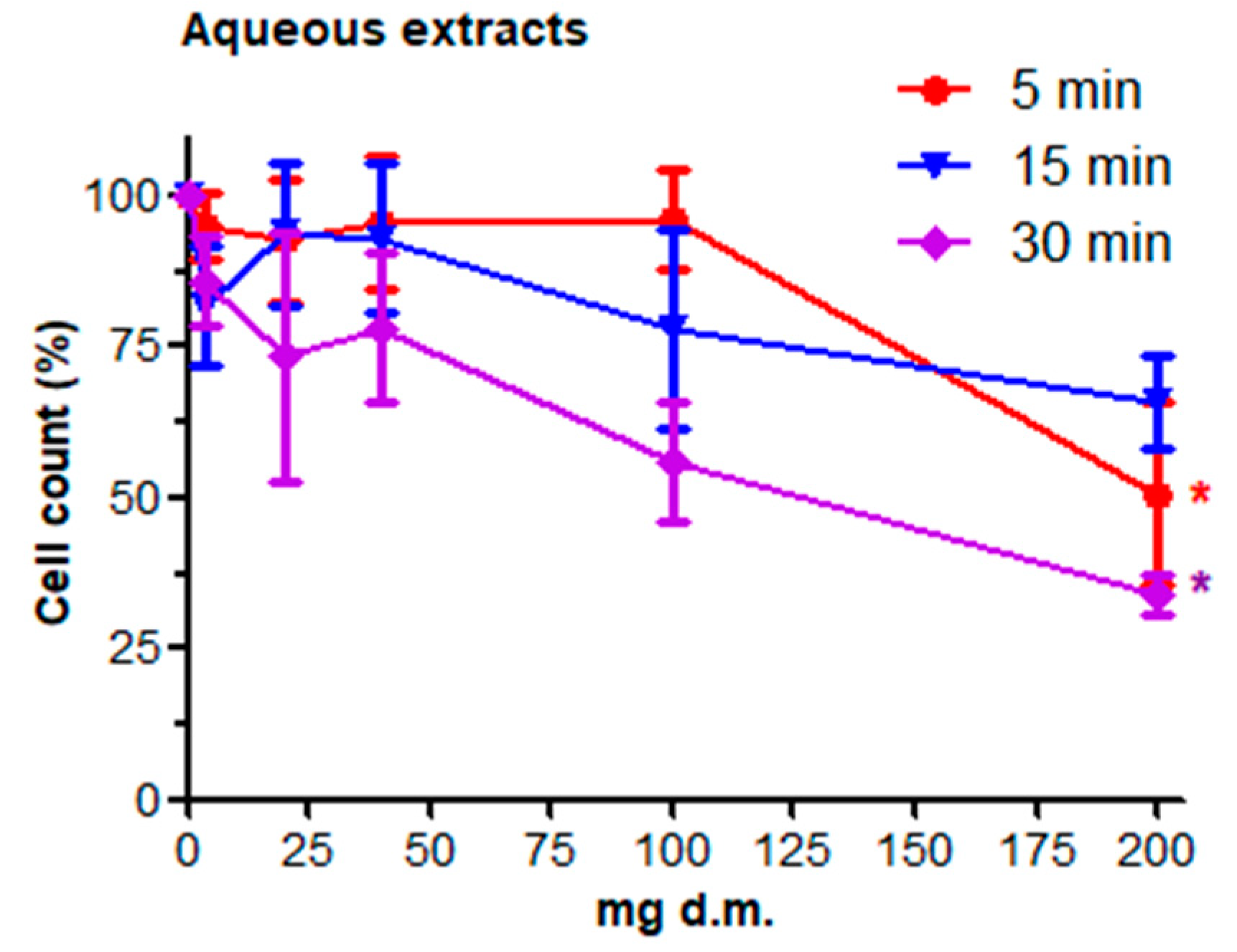
3.1. Cell Count and Viability: AO/DAPI Double Staining
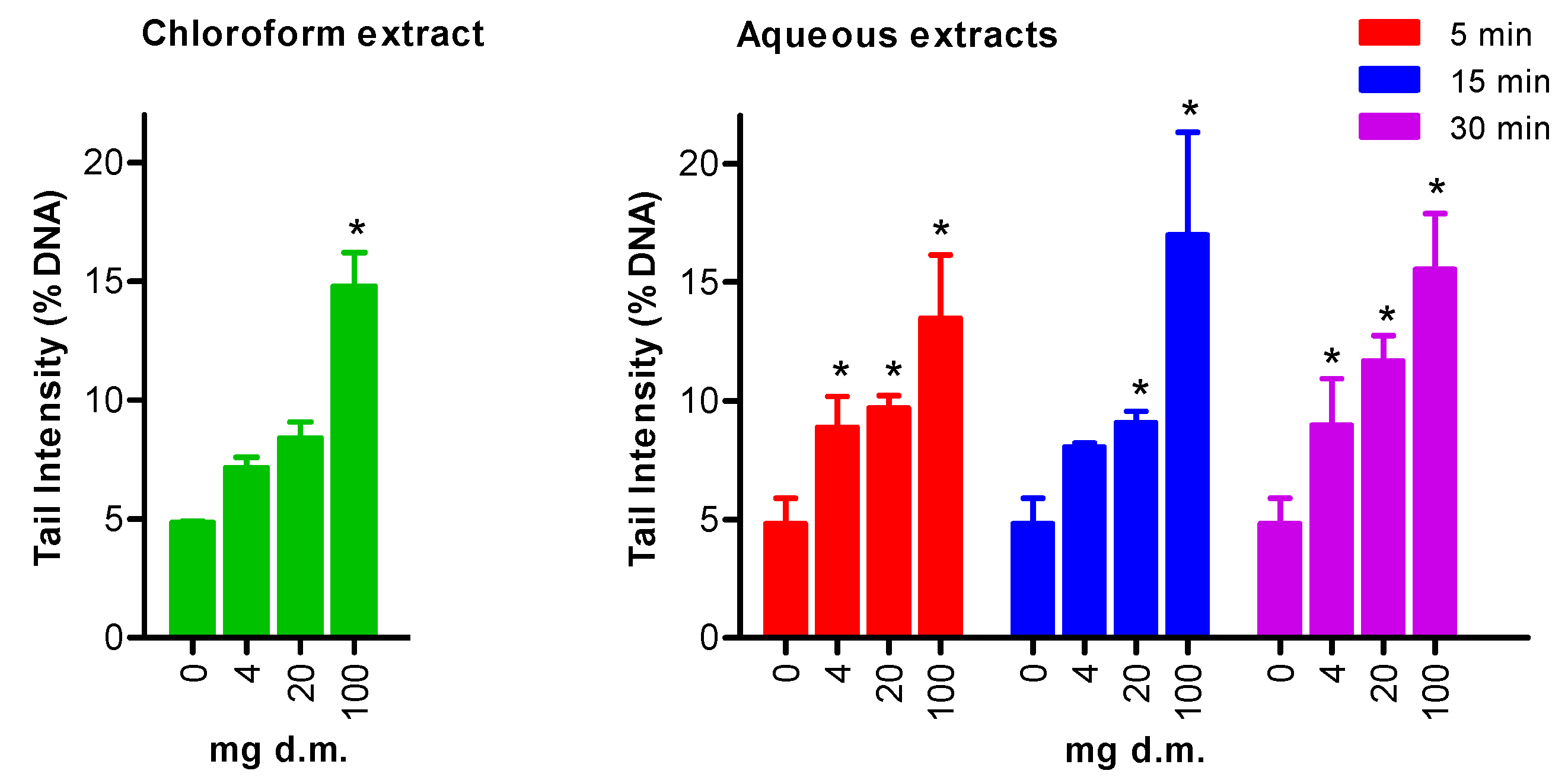
3.2. Genotoxicity Testing: Comet Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heywood, V.; Harborne, J.; Turner, B.; Linnean Society of London; Phytochemical Group. The Biology and Chemistry of the Compositae; Academic Press: London, UK, 1977. [Google Scholar]
- Gledhill, D. The Names of Plants, 4th ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Nicolini, G. Enciclopedia Botanica Motta; Federico Motta Editore: Milano, Italy, 1960; Volume 3. [Google Scholar]
- Von Linnaeus, C. Species Plantarum; Imprensis Laurentii Salvii: Stockholm, Sweden, 1753. [Google Scholar]
- Robinson, D.E.; O’Donovan, J.T.; Sharma, M.P.; Doohan, D.J.; Figueroa, R. The Biology of Canadian Weeds. 123. Senecio vulgaris L. Can. J. Plant Sci. 2003, 83, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Mitich, L.W. Common Groundsel (Senecio vulgaris). Weed Technol. 1995, 9, 209–211. [Google Scholar] [CrossRef]
- Simpson, J.; Weiner, E. The Oxford English Dictionary, 2nd ed.; Claredon Press: Oxford, UK, 1989. [Google Scholar]
- Mansour, R.M.; Saleh, N.A. Flavonoids of Three Local Senecio Species. Phytochemistry 1981, 20, 1180–1181. [Google Scholar] [CrossRef]
- Stuart, M. The Encyclopedia of Herbs and Herbalism; Orbis Books: London, UK, 1979. [Google Scholar]
- Molyneux, R.J.; Johnson, A.E.; Olsen, J.D.; Baker, D.C. Toxicity of Pyrrolizidine Alkaloids from Riddell Groundsel (Senecio riddellii) to Cattle. Am. J. Vet. Res. 1991, 52, 146–151. [Google Scholar] [PubMed]
- Baker, H.G. The Evolution of Weeds. In Review of Ecology and Systematics; Annual Reviews: San Mateo, CA, USA, 1974; Volume 5, pp. 1–24. [Google Scholar]
- Steenkamp, V.; Stewart, M.J.; Zuckerman, M. Clinical and Analytical Aspects of Pyrrolizidine Poisoning Caused by South African Traditional Medicines. Ther. Drug Monit. 2000, 22, 302–306. [Google Scholar] [CrossRef]
- The Plant List 2022. Available online: https://www.dixon.org/events/event/23544/ (accessed on 20 September 2022).
- Chen, L.; Mulder, P.P.J.; Louisse, J.; Peijnenburg, A.; Wesseling, S.; Rietjens, I.M.C.M. Risk Assessment for Pyrrolizidine Alkaloids Detected in (Herbal) Teas and Plant Food Supplements. Regul. Toxicol. Pharmacol. 2017, 86, 292–302. [Google Scholar] [CrossRef]
- Teuscher, E.; Lindequist, U. Biogene Gifte: Biologie, Chemie, Pharmakologie, Toxikologie; Wissenschaftliche Verlagsgesellschaft: Stuttgart, Germany, 2010. [Google Scholar]
- Bodi, D.; Ronczka, S.; Gottschalk, C.; Behr, N.; Skibba, A.; Wagner, M.; Lahrssen-Wiederholt, M.; Preiss-Weigert, A.; These, A. Determination of Pyrrolizidine Alkaloids in Tea, Herbal Drugs and Honey. Food Addit. Contam. Part A 2014, 31, 1886–1895. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for Human Health Related to the Presence of Pyrrolizidine Alkaloids in Honey, Tea, Herbal Infusions and Food Supplements. EFSA J. 2017, 15, e04908. [Google Scholar] [CrossRef] [Green Version]
- WHO/IPCS. Pyrrolizidine Alkaloids; Environmental Health Criteria; World Health Organization/International Programme on Chemical Safety: Geneva, Switzerland, 1988; Volume 80. [Google Scholar]
- Doorduin, L.J.; Vrieling, K. A Review of the Phytochemical Support for the Shifting Defence Hypothesis. Phytochem. Rev. 2011, 10, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Mei, N.; Fu, P.P. Genotoxicity of Pyrrolizidine Alkaloids. J. Appl. Toxicol. 2010, 30, 183–196. [Google Scholar] [CrossRef]
- Moyano, M.R.; García, A.; Rueda, A.; Molina, A.M.; Mendez, A.; Infante, F. Echium vulgare and Senecio vulgaris Poisoning in Fighting Bulls. J. Vet. Med. Ser. A 2006, 53, 24–25. [Google Scholar] [CrossRef] [PubMed]
- Stegelmeier, B.L.; Edgar, J.A.; Colegate, S.M.; Gardner, D.R.; Schoch, T.K.; Coulombe, R.A.; Molyneux, R.J. Pyrrolizidine Alkaloid Plants, Metabolism and Toxicity. J. Nat. Toxins 1999, 8, 95–116. [Google Scholar] [PubMed]
- Wiedenfeld, H.; Edgar, J. Toxicity of Pyrrolizidine Alkaloids to Humans and Ruminants. Phytochem. Rev. 2011, 10, 137–151. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Gardner, D.L.; Colegate, S.M.; Edgar, J.A. Pyrrolizidine Alkaloid Toxicity in Livestock: A Paradigm for Human Poisoning? Food Addit. Contam. Part A 2011, 28, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Mulder, P.P.J.; López, P.; Castellari, M.; Bodi, D.; Ronczka, S.; Preiss-Weigert, A.; These, A. Occurrence of Pyrrolizidine Alkaloids in Animal- and Plant-Derived Food: Results of a Survey across Europe. Food Addit. Contam. Part A 2018, 35, 118–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohabbat, O.; Younos, M.S.; Merzad, A.A.; Srivastava, R.N.; Sediq, G.G.; Aram, G.N. An Outbreak of Hepatic Veno-Occlusive Disease in North-Western Afghanistan. Lancet 1976, 2, 269–271. [Google Scholar] [CrossRef]
- Tandon, B.N.; Tandon, H.D.; Tandon, R.K.; Narndranathan, M.; Joshi, Y.K. An Epidemic of Veno-Occlusive Disease of Liver in Central India. Lancet 1976, 2, 271–272. [Google Scholar] [CrossRef]
- Van den Berg, S.J.P.L.; Restani, P.; Boersma, M.G.; Delmulle, L.; Rietjens, I.M.C.M. Levels of Genotoxic and Carcinogenic Ingredients in Plant Food Supplements and Associated Risk Assessment. Food Nutr. Sci. 2011, 2, 989–1010. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, S.J.P.L.; Serra-Majem, L.; Coppens, P.; Rietjens, I.M.C.M. Safety Assessment of Plant Food Supplements (PFS). Food Funct. 2011, 2, 760–768. [Google Scholar] [CrossRef] [Green Version]
- Schrenk, D.; Gao, L.; Lin, G.; Mahony, C.; Mulder, P.P.J.; Peijnenburg, A.; Pfuhler, S.; Rietjens, I.M.C.M.; Rutz, L.; Steinhoff, B.; et al. Pyrrolizidine Alkaloids in Food and Phytomedicine: Occurrence, Exposure, Toxicity, Mechanisms, and Risk Assessment—A Review. Food Chem. Toxicol. 2020, 136, 111107. [Google Scholar] [CrossRef]
- Cheng, D.; Nguyen, V.-T.; Ndihokubwayo, N.; Ge, J.; Mulder, P.P.J. Pyrrolizidine Alkaloid Variation in Senecio Vulgaris Populations from Native and Invasive Ranges. PeerJ 2017, 5, e3686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flade, J.; Beschow, H.; Wensch-Dorendorf, M.; Plescher, A.; Wätjen, W. Occurrence of Nine Pyrrolizidine Alkaloids in Senecio vulgaris L. Depending on Developmental Stage and Season. Plants 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, U. BBCH Monograph. In Growth Stages of Mono- and Dicotyledonous Plants; Open Agrar Repositorium: Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Mattocks, A.R. Relation of Structural Features to Pyrrolic Metabolites in Livers of Rats given Pyrrolizidine Alkaloids and Derivatives. Chem. Biol. Interact. 1981, 35, 301–310. [Google Scholar] [CrossRef]
- Vollmer, J.J.; Steiner, N.C.; Larsen, G.Y.; Muirhead, K.M.; Molyneux, R.J. Pyrrolizidine Alkaloids: Testing for Toxic Constituents of Comfrey. J. Chem. Educ. 1987, 64, 1027. [Google Scholar] [CrossRef]
- Aden, D.P.; Fogel, A.; Plotkin, S.; Damjanov, I.; Knowles, B.B. Controlled Synthesis of HBsAg in a Differentiated Human Liver Carcinoma-Derived Cell Line. Nature 1979, 282, 615–616. [Google Scholar] [CrossRef]
- Knowles, B.B.; Howe, C.C.; Aden, D.P. Human Hepatocellular Carcinoma Cell Lines Secrete the Major Plasma Proteins and Hepatitis B Surface Antigen. Science 1980, 209, 497–499. [Google Scholar] [CrossRef]
- López-Terrada, D.; Cheung, S.W.; Finegold, M.J.; Knowles, B.B. Hep G2 Is a Hepatoblastoma-Derived Cell Line. Hum. Pathol. 2009, 40, 1512–1515. [Google Scholar] [CrossRef]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and Functional Characterization of Human Hepatoma HepG2 Cells. Methods Mol. Biol. 2015, 1250, 77–93. [Google Scholar] [CrossRef]
- Knasmüller, S.; Parzefall, W.; Sanyal, R.; Ecker, S.; Schwab, C.; Uhl, M.; Mersch-Sundermann, V.; Williamson, G.; Hietsch, G.; Langer, T.; et al. Use of Metabolically Competent Human Hepatoma Cells for the Detection of Mutagens and Antimutagens. Mutat. Res. 1998, 402, 185–202. [Google Scholar] [CrossRef]
- OECD. Guidance Document on Revisions to OECD Genetic Toxicology Test Guidelines; Organisation for Economic Co-Operation and Development: Paris, France, 2015. [Google Scholar]
- Villarini, M.; Pagiotti, R.; Dominici, L.; Fatigoni, C.; Vannini, S.; Levorato, S.; Moretti, M. Investigation of the Cytotoxic, Genotoxic, and Apoptosis-Inducing Effects of Estragole Isolated from Fennel (Foeniculum Vulgare). J. Nat. Prod. 2014, 77, 773–778. [Google Scholar] [CrossRef]
- Shah, D.; Naciri, M.; Clee, P.; Al-Rubeai, M. NucleoCounter—An Efficient Technique for the Determination of Cell Number and Viability in Animal Cell Culture Processes. Cytotechnology 2006, 51, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nunzio, M.; Valli, V.; Tomás-Cobos, L.; Tomás-Chisbert, T.; Murgui-Bosch, L.; Danesi, F.; Bordoni, A. Is Cytotoxicity a Determinant of the Different In Vitro and In Vivo Effects of Bioactives? Complement. Altern. Med. 2017, 17, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single Cell Gel/Comet Assay: Guidelines for in Vitro and in Vivo Genetic Toxicology Testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Kopp, T.; Abdel-Tawab, M.; Mizaikoff, B. Extracting and Analyzing Pyrrolizidine Alkaloids in Medicinal Plants: A Review. Toxins 2020, 12, 320. [Google Scholar] [CrossRef] [PubMed]
- Rutz, L.; Gao, L.; Küpper, J.-H.; Schrenk, D. Structure-Dependent Genotoxic Potencies of Selected Pyrrolizidine Alkaloids in Metabolically Competent HepG2 Cells. Arch. Toxicol. 2020, 94, 4159–4172. [Google Scholar] [CrossRef]
- Schrenk, D.; Fahrer, J.; Allemang, A.; Fu, P.; Lin, G.; Mahony, C.; Mulder, P.P.J.; Peijnenburg, A.; Pfuhler, S.; Rietjens, I.M.C.M.; et al. Novel Insights into Pyrrolizidine Alkaloid Toxicity and Implications for Risk Assessment: Occurrence, Genotoxicity, Toxicokinetics, Risk Assessment-A Workshop Report. Planta Med. 2022, 88, 98–117. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acito, M.; Russo, C.; Fatigoni, C.; Mercanti, F.; Moretti, M.; Villarini, M. Cytotoxicity and Genotoxicity of Senecio vulgaris L. Extracts: An In Vitro Assessment in HepG2 Liver Cells. Int. J. Environ. Res. Public Health 2022, 19, 14824. https://doi.org/10.3390/ijerph192214824
Acito M, Russo C, Fatigoni C, Mercanti F, Moretti M, Villarini M. Cytotoxicity and Genotoxicity of Senecio vulgaris L. Extracts: An In Vitro Assessment in HepG2 Liver Cells. International Journal of Environmental Research and Public Health. 2022; 19(22):14824. https://doi.org/10.3390/ijerph192214824
Chicago/Turabian StyleAcito, Mattia, Carla Russo, Cristina Fatigoni, Federica Mercanti, Massimo Moretti, and Milena Villarini. 2022. "Cytotoxicity and Genotoxicity of Senecio vulgaris L. Extracts: An In Vitro Assessment in HepG2 Liver Cells" International Journal of Environmental Research and Public Health 19, no. 22: 14824. https://doi.org/10.3390/ijerph192214824
APA StyleAcito, M., Russo, C., Fatigoni, C., Mercanti, F., Moretti, M., & Villarini, M. (2022). Cytotoxicity and Genotoxicity of Senecio vulgaris L. Extracts: An In Vitro Assessment in HepG2 Liver Cells. International Journal of Environmental Research and Public Health, 19(22), 14824. https://doi.org/10.3390/ijerph192214824