
Hemocytes: A Useful Tool for Assessing the Toxicity of Microplastics, Heavy Metals, and Pesticides on Aquatic Invertebrates

 ,
,  ,
, 

Abstract
:1. Introduction
1.1. Hemolymph
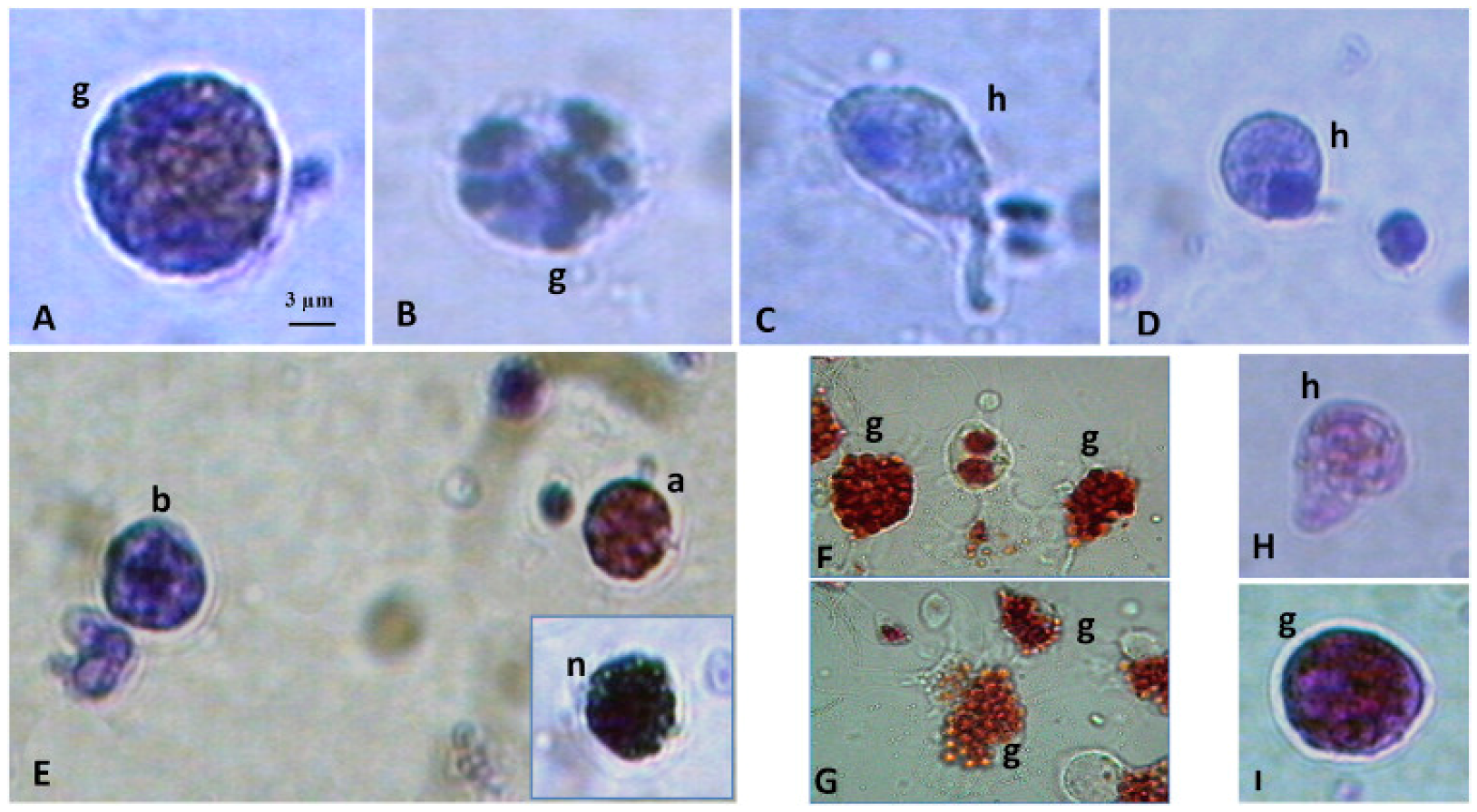
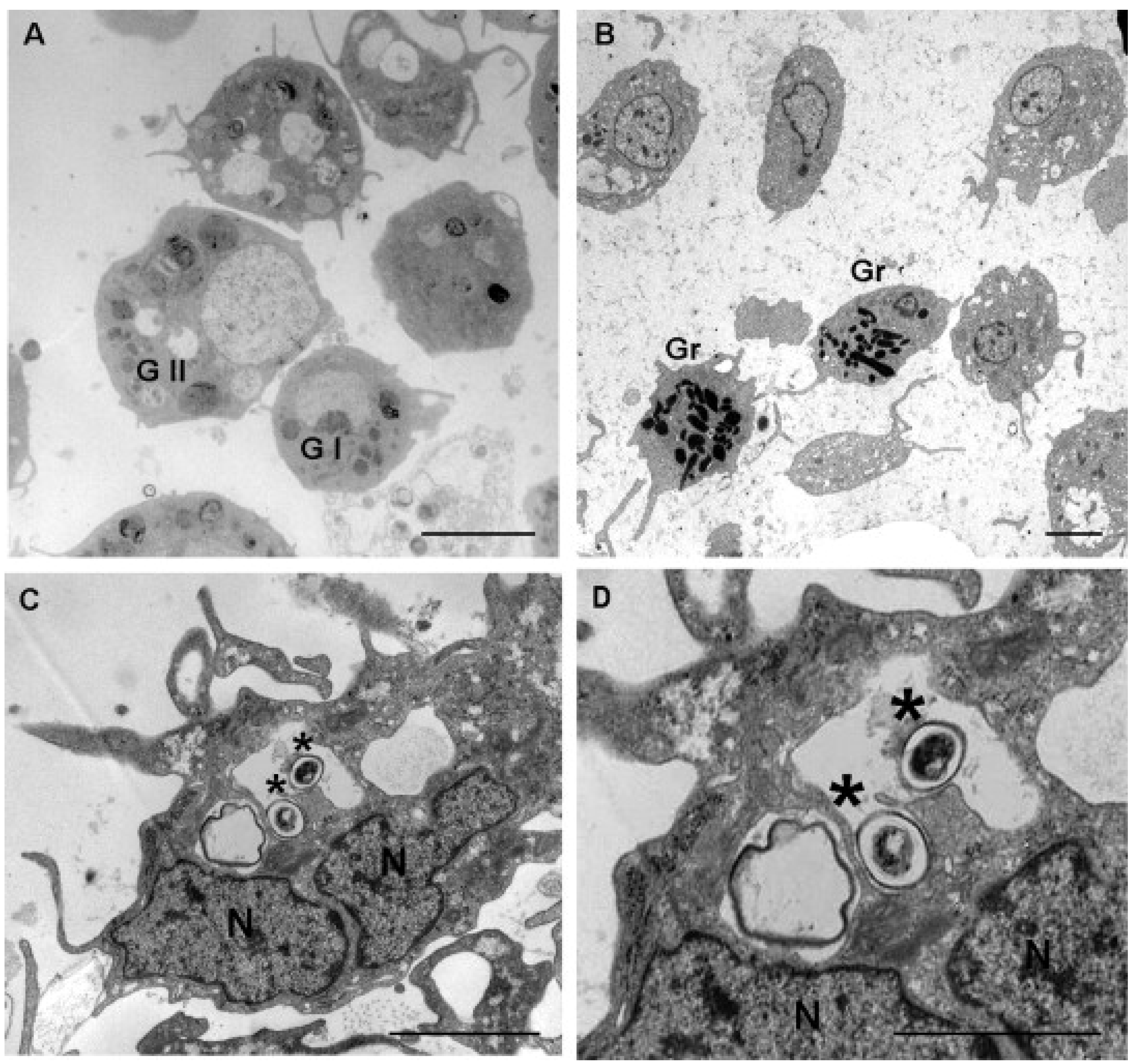
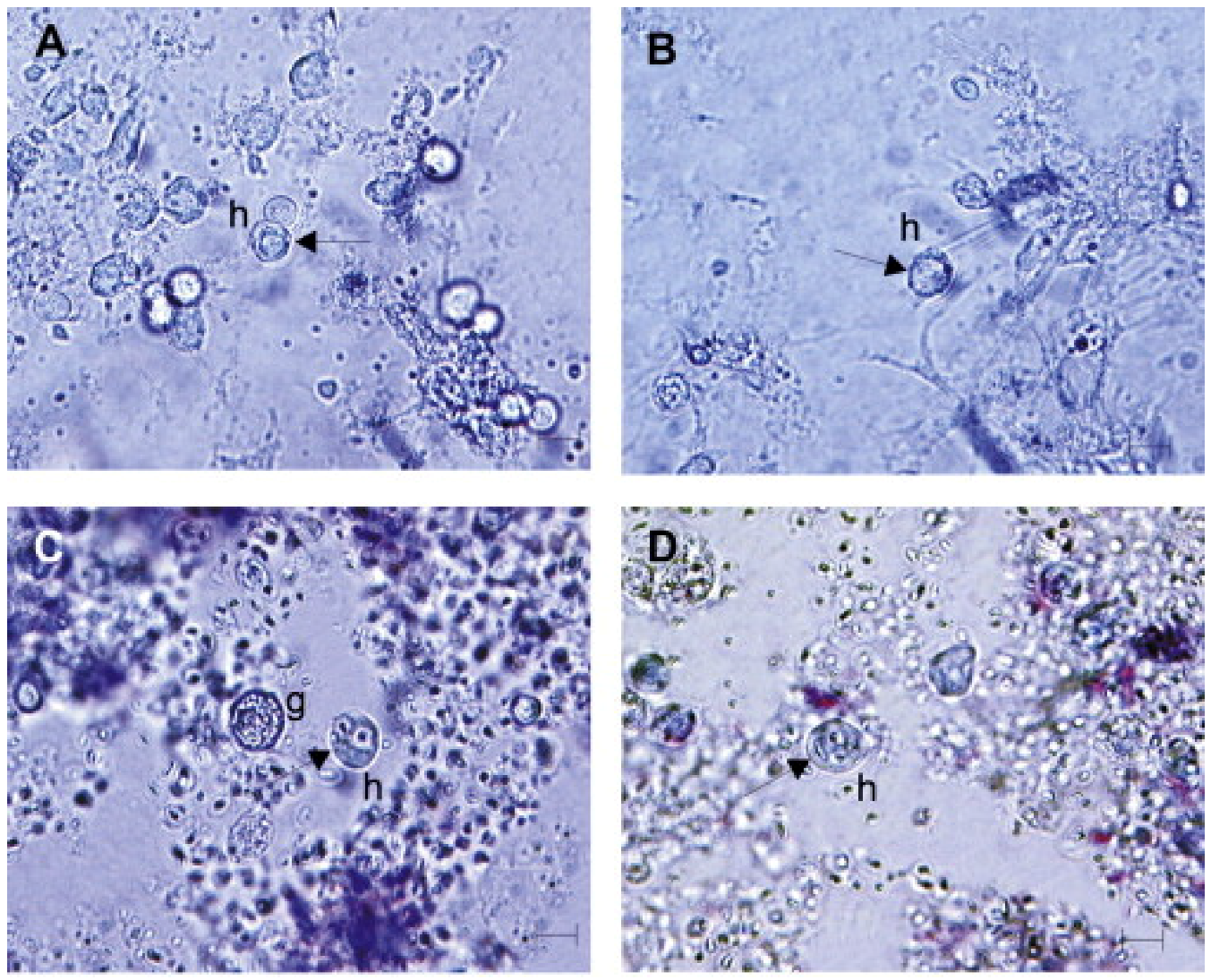
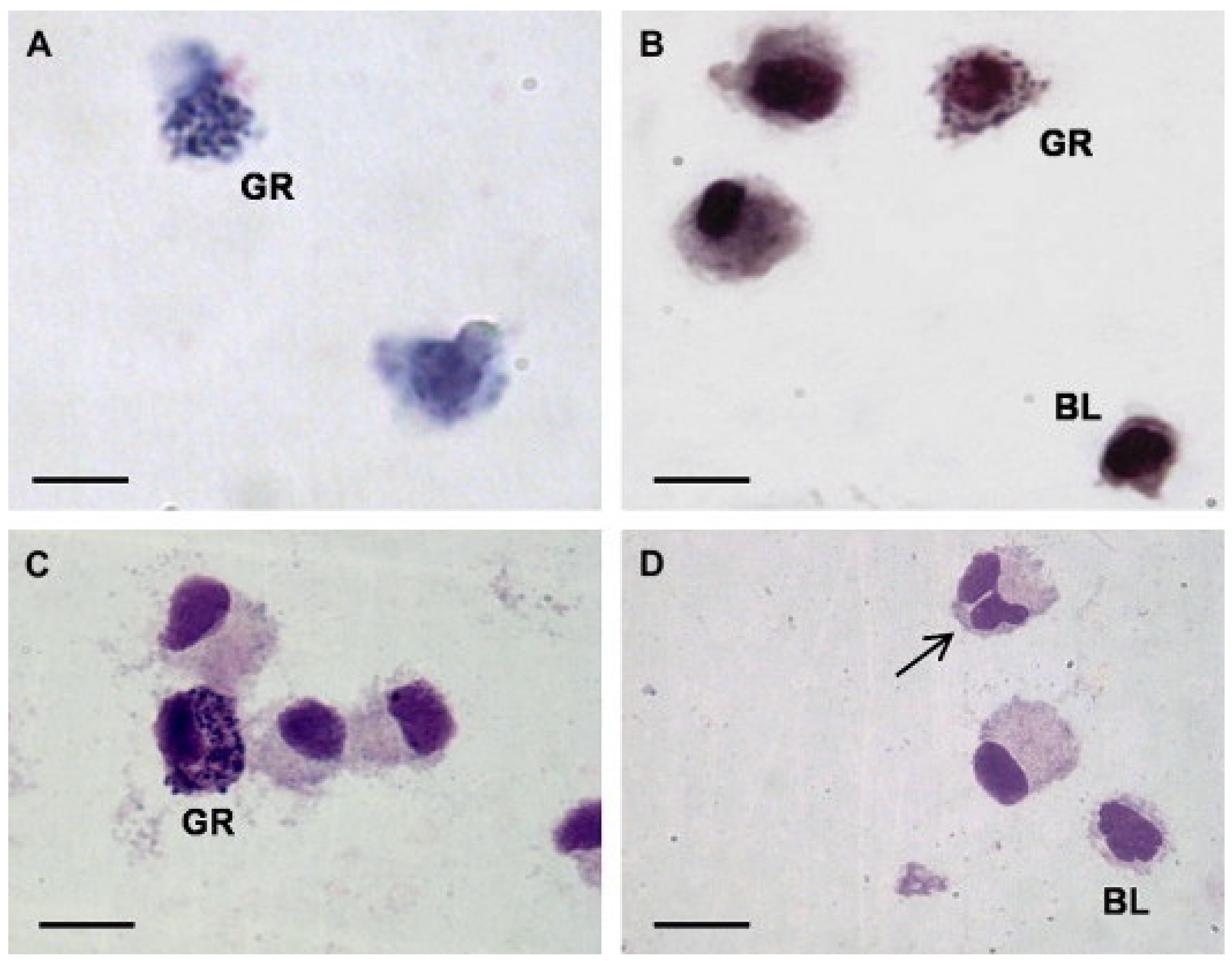
1.2. Hemocytes
1.3. Model Organisms
2. Hemocytes’ Response to Xenobiotics
2.1. Heavy Metals
2.2. Pesticides
2.3. Hydrocarbons and Oil Spills
2.4. Micro- and Nano-Plastics
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maciorowski, H.D.; Clarke, R. Advantages and Disadvantages of Using Invertebrates in Toxicity Testing. In Aquatic Invertebrate Bioassays, ASTM STP 715; American Society for Testing and Materials: Philadelphia, PA, USA, 1980; pp. 36–47. [Google Scholar]
- Wilson-Sanders, S.E. Invertebrate Models for Biomedical Research, Testing, and Education. ILAR J. 2011, 52, 126–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.E.; Heatley, J.J.; Chitty, J. Clinical techniques of invertebrates. Vet. Clin. Exot. Anim. Pract. 2006, 9, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, M.; Cornillie, P.; Dierick, M.; Adriaens, D.; Nevejan, N.; Bossier, P.; Van den Broeck, W.; Sorgeloos, P.; Defoirdt, T.; Declercq, A.M. The blue mussel inside: 3D visualization and description of the vascular-related anatomy of Mytilus edulis to unravel hemolymph extraction. Sci. Rep. 2020, 10, 6773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stara, A.; Pagano, M.; Albano, M.; Savoca, S.; Di Bella, G.; Albergamo, A.; Koutkova, Z.; Sandova, M.; Velisek, J.; Fabrello, J.; et al. Effects of long-term exposure of Mytilus galloprovincialis to thiacloprid: A multibiomarker approach. Environ. Pollut. 2021, 289, 117892. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.; Porcino, C.; Briglia, M.; Fiorino, E.; Vazzana, M.; Silvestro, S.; Faggio, C. The Influence of Exposure of Cadmium Chloride and Zinc Chloride on Haemolymph and Digestive Gland Cells from Mytilus galloprovincialis. Int. J. Environ. Res. 2017, 11, 207–216. [Google Scholar] [CrossRef]
- Torre, A.; Trischitta, F.; Faggio, C. Effect of CdCl2 on Regulatory Volume Decrease (RVD) in Mytilus galloprovincialis digestive cells. Toxicol. In Vitro 2013, 27, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Albano, M.; Zuskova, E.; Velisek, J.; Matozzo, V.; Faggio, C. Acute effects of neonicotinoid insecticides on Mytilus galloprovincialis: A case study with the active compound thiacloprid and the commercial formulation calypso 480 SC. Ecotoxicol. Environ. Saf. 2020, 203, 110980. [Google Scholar] [CrossRef]
- Balbi, T.; Auguste, M.; Cortese, K.; Montagna, M.; Borello, A.; Pruzzo, C.; Vezzulli, L.; Canesi, L. Responses of Mytilus galloprovincialis to Challenge with the Emerging Marine Pathogen Vibrio Coralliilyticus. Fish Shellfish Immunol. 2019, 84, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, L.L.; Stoskopf, M.K.; Bogan, A.; Showers, W.; Kwak, T.; Hanlon, S.; Levine, J.F. Evaluation of a nonlethal technique for hemolymph collection in Elliptio complanata, a freshwater bivalve (Mollusca: Unionidae). Dis. Aquat. Org. 2005, 65, 159–165. [Google Scholar] [CrossRef]
- Fredrick, W.S.; Ravichandran, S. Hemolymph proteins in marine crustaceans. Asian Pac. J. Trop. Biomed. 2012, 2, 496–502. [Google Scholar] [CrossRef]
- Vazquez, L.; Alpuche, J.; Maldonado, G.; Agundis, C.; Pereyra-Morales, A.; Zenteno, E. Review: Immunity mechanisms in crustaceans. Innate Immun. 2009, 15, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Matozzo, V.; Marin, M.G. The role of haemocytes from the crab Carcinus aestuarii (Crustacea, Decapoda) in immune responses: A first survey. Fish Shellfish Immunol. 2010, 28, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Travers, M.-A.; da Silva, P.M.; Le Goïc, N.; Marie, D.; Donval, A.; Huchette, S.; Koken, M.; Paillard, C. Morphologic, cytometric and functional characterisation of abalone (Haliotis tuberculata) haemocytes. Fish Shellfish Immunol. 2008, 24, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Carballal, M.; Lopez, M.; Azevedo, C.; Villalba, A. Hemolymph Cell Types of the Mussel Mytilus galloprovincialis. Dis. Aquat. Org. 1997, 29, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Matozzo, V.; Pagano, M.; Spinelli, A.; Caicci, F.; Faggio, C. Pinna nobilis: A Big Bivalve with Big Haemocytes? Fish Shellfish Immunol. 2016, 55, 529–534. [Google Scholar] [CrossRef]
- Accorsi, A.; Bucci, L.; de Eguileor, M.; Ottaviani, E.; Malagoli, D. Comparative analysis of circulating hemocytes of the freshwater snail Pomacea canaliculata. Fish Shellfish Immunol. 2013, 34, 1260–1268. [Google Scholar] [CrossRef]
- Chiarelli, R.; Roccheri, M.C. Marine Invertebrates as Bioindicators of Heavy Metal Pollution. Open J. Met. 2014, 4, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Cima, F. Microscopy Methods for Morpho-Functional Characterisation of Marine Invertebrate Haemocytes. In Microscopy: Science, Technology, Applications and Education; Formatex Research Center: Badajoz, Spain, 2010; Volume 2, pp. 1100–1107. [Google Scholar]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The natural indicator of environmental pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Knakievicz, T. Planarians as invertebrate bioindicators in freshwater environmental quality: The biomarkers approach. Ecotoxicol. Environ. Contam. 2014, 9, 1–12. [Google Scholar] [CrossRef]
- Waykar, B.; Deshmukh, G. Evaluation of Bivalves as Bioindicators of Metal Pollution in Freshwater. Bull. Environ. Contam. Toxicol. 2012, 88, 48–53. [Google Scholar] [CrossRef]
- Matozzo, V.; Chinellato, A.; Munari, M.; Finos, L.; Bressan, M.; Marin, M.G. First Evidence of Immunomodulation in Bivalves under Seawater Acidification and Increased Temperature. PLoS ONE 2012, 7, e33820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnaire, B.; Thomas-Guyon, H.; Burgeot, T.; Renault, T. Pollutant effects on Pacific oyster, Crassostrea gigas (Thunberg), hemocytes: Screening of 23 molecules using flow cytometry. Cell Biol. Toxicol. 2006, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Berthod, C.; Bénard-Déraspe, M.-H.; Laplante, J.-F.; Lemaire, N.; Nadeau, M.; Toupoint, N.; Triffault-Bouchet, G.; Saint-Louis, R. Hemocyte Health Status Based on Four Biomarkers to Assess Recovery Capacity in American Lobster (Homarus americanus) after Exposure to Marine Diesel and Diluted Bitumen. J. Mar. Sci. Eng. 2021, 9, 370. [Google Scholar] [CrossRef]
- Parisi, M.G.; Pirrera, J.; La Corte, C.; Dara, M.; Parrinello, D.; Cammarata, M. Effects of organic mercury on Mytilus galloprovincialis hemocyte function and morphology. J. Comp. Physiol. B 2020, 191, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Welbourn, P. Effects of mercury exposure on ionic regulation in the crayfish Orconectes propinquus. Environ. Pollut. 1993, 82, 139–142. [Google Scholar] [CrossRef]
- Saravanan, R.; Sugumar, V.; Mahin, M.I.B. Heavy metal stress induced hyperglycemia in blue swimmer crab, Portunus pelagicus. Acta Oceanol. Sin. 2018, 37, 47–53. [Google Scholar] [CrossRef]
- Rault, P.; Fortier, M.; Pédelucq, J.; Lacaze, E.; Brousseau, P.; Auffret, M.; Fournier, M. Immunotoxicity of Heavy Metals (Silver, Cadmium, Mercury and Lead) on Marine Bivalve Mytilus Edulis: In Vitro Exposure of Hemocytes. J. Xenobiotics 2013, 3, e8. [Google Scholar] [CrossRef] [Green Version]
- Matozzo, L.B.V.; Ballarin, L.; Pampanin, D.M.; Marin, M.G. Effects of Copper and Cadmium Exposure on Functional Responses of Hemocytes in the Clam, Tapes philippinarum. Arch. Environ. Contam. Toxicol. 2001, 41, 163–170. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Alfaro, A.C.; Merien, F.; Lulijwa, R.; Young, T. Copper-induced immunomodulation in mussel (Perna canaliculus) haemocytes. Metallomics 2018, 10, 965–978. [Google Scholar] [CrossRef]
- Huang, X.; Jiang, X.; Sun, M.; Dupont, S.; Huang, W.; Hu, M.; Li, Q.; Wang, Y. Effects of copper on hemocyte parameters in the estuarine oyster Crassostrea rivularis under low pH conditions. Aquat. Toxicol. 2018, 203, 61–68. [Google Scholar] [CrossRef]
- Evariste, L.; Rioult, D.; Brousseau, P.; Geffard, A.; David, E.; Auffret, M.; Fournier, M.; Betoulle, S. Differential Sensitivity to Cadmium of Immunomarkers Measured in Hemocyte Subpopulations of Zebra Mussel Dreissena Polymorpha. Ecotoxicol. Environ. Saf. 2017, 137, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Evans, S.; Hughes, F. Cadmium-induced apoptosis in oyster hemocytes involves disturbance of cellular energy balance but no mitochondrial permeability transition. J. Exp. Biol. 2004, 207, 3369–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boissel, P.G.J.; Fournier, M.; Rodriguez-Lecompte, J.C.; McKenna, P.; Kibenge, F.; Siah, A. Functional and molecular responses of the blue mussel Mytilus edulis’ hemocytes exposed to cadmium–An in vitro model and transcriptomic approach. Fish Shellfish Immunol. 2017, 67, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, T.; Lei, W.-W.; Liu, D.-M.; Li, Y.-J.; Xuan, R.-J.; Ma, J.-J. Cadmium-Induced Oxidative Stress and Apoptotic Changes in the Testis of Freshwater Crab, Sinopotamon henanense. PLoS ONE 2011, 6, e27853. [Google Scholar] [CrossRef] [Green Version]
- Lorenzon, S.; Francese, M.; Smith, V.; Ferrero, E. Heavy metals affect the circulating haemocyte number in the shrimp Palaemon elegans. Fish Shellfish Immunol. 2001, 11, 459–472. [Google Scholar] [CrossRef]
- Guria, S. Lead (Pb) Induces Paraptosis like Cell Death in Hemocytes of Lamellidens Sp.: A Preliminary Study. World Sci. News 2018, 94, 163–172. [Google Scholar]
- Mosher, S.; Cope, W.G.; Weber, F.X.; Shea, D.; Kwak, T.J. Effects of Lead on Na+, K+-ATPase and Hemolymph Ion Concentrations in the Freshwater Mussel Elliptio Complanata. Environ. Toxicol. 2010, 27, 268–276. [Google Scholar] [CrossRef]
- Ray, M.; Bhunia, A.S.; Bhunia, N.S.; Ray, S. Density shift, morphological damage, lysosomal fragility and apoptosis of hemocytes of Indian molluscs exposed to pyrethroid pesticides. Fish Shellfish Immunol. 2013, 35, 499–512. [Google Scholar] [CrossRef]
- de Mello, G.L.; Junior, H.A.; Garcia, S.; Vinatea, L. Acute Toxicity of Pyrazosulfuron-Ethyl and Permethrin to Juvenile Litopenaeus Vannamei. Acta Sci. Biol. Sci. 2011, 33, 1–6. [Google Scholar]
- Matozzo, V.; Zampieri, C.; Munari, M.; Marin, M.G. Glyphosate Affects Haemocyte Parameters in the Clam Ruditapes Philippinarum. Mar. Environ. Res. 2019, 146, 66–70. [Google Scholar] [CrossRef]
- Mottier, A.; Serpentini, A.; Dallas, L.; James, A.; Lebel, J.-M.; Costil, K. In vitro effects of glyphosate-based herbicides and related adjuvants on primary culture of hemocytes from Haliotis tuberculata. Fish Shellfish Immunol. 2020, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Yang, X.; Yan, G.; Huang, Y.; Zuo, F.; Shen, Y.; Ding, Y.; Cheng, Y. Effects of glyphosate on immune responses and haemocyte DNA damage of Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2017, 71, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.I.; Gnatyshyna, L.L.; Stoliar, O.B. In situ exposure history modulates the molecular responses to carbamate fungicide Tattoo in bivalve mollusk. Ecotoxicology 2013, 22, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Gnatyshyna, L.; Falfushynska, H.; Stoliar, O.; Dallinger, R. Preliminary Study of Multiple Stress Response Reactions in the Pond Snail Lymnaea stagnalis Exposed to Trace Metals and a Thiocarbamate Fungicide at Environmentally Relevant Concentrations. Arch. Environ. Contam. Toxicol. 2020, 79, 89–100. [Google Scholar] [CrossRef]
- Chinonso, U.C.; Olatunji, M.R.; Omolara, L.A.A. Sublethal Effects of Organophosphate Chlorpyrifos on Hemato-Immunological Parameters of the Gercacinid Crab, Cardiosoma Armatum (Herklots, 1851). J. Biol. Res. Biotechnol. 2021, 19, 1185–1191. [Google Scholar]
- Garate, O.F.; Gazzaniga, S.; Cochón, A.C. A comparative study of enzymatic and immunological parameters in Planorbarius corneus and Biomphalaria glabrata exposed to the organophosphate chlorpyrifos. Aquat. Toxicol. 2020, 225, 105544. [Google Scholar] [CrossRef]
- Maharajan, A.; Narayanaswamy, Y.; Ganapiriya, V. Haematological Changes of Fresh Water Crab, Paratelphusa Jacquemontii in Response to the Combination of Chlorpyrifos and Cypermethrin (Nurocombi) Insecticide. Ann. Aquac. Res. 2017, 4, 1041. [Google Scholar]
- Bado-Nilles, A.; Gagnaire, B.; Thomas-Guyon, H.; Le Floch, S.; Renault, T. Effects of 16 pure hydrocarbons and two oils on haemocyte and haemolymphatic parameters in the Pacific oyster, Crassostrea gigas (Thunberg). Toxicol. In Vitro 2008, 22, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Donaghy, L.; Hong, H.-K.; Lee, H.-J.; Jun, J.-C.; Park, Y.-J.; Choi, K.-S. Hemocyte parameters of the Pacific oyster Crassostrea gigas a year after the Hebei Spirit oil spill off the west coast of Korea. Helgol. Mar. Res. 2010, 64, 349–355. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, L.; Liu, J. Immunotoxicity Effect of Benzo[α]Pyrene on Scallop Chlamys Farreri. J. Ocean Univ. China 2009, 8, 89–94. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Thilagam, H.; Huang, W.-B.; Wang, K.-J. Immunomodulation in the marine gastropod Haliotis diversicolor exposed to benzo(a)pyrene. Chemosphere 2009, 75, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Capolupo, M.; Valbonesi, P.; Fabbri, E. A Comparative Assessment of the Chronic Effects of Micro- and Nano-Plastics on the Physiology of the Mediterranean Mussel Mytilus galloprovincialis. Nanomaterials 2021, 11, 649. [Google Scholar] [CrossRef]
- Paul-Pont, I.; Lacroix, C.; González Fernández, C.; Hégaret, H.; Lambert, C.; Le Goïc, N.; Frère, L.; Cassone, A.-L.; Sussarellu, R.; Fabioux, C.; et al. Exposure of Marine Mussels Mytilus Spp. to Polystyrene Microplastics: Toxicity and Influence on Fluoranthene Bioaccumulation. Environ. Pollut. 2016, 216, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, M.A.; Dissanayake, A.; Galloway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested Microscopic Plastic Translocates to the Circulatory System of the Mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Liddle, C.; Consolandi, G.; Drago, C.; Hird, C.; Lindeque, P.K.; Galloway, T.S. Microplastics, Microfibres and Nanoplastics Cause Variable Sub-Lethal Responses in Mussels (Mytilus Spp.). Mar. Pollut. Bull. 2020, 160, 111552. [Google Scholar] [CrossRef] [PubMed]
- Sendra, M.; Carrasco-Braganza, M.I.; Yeste, P.M.; Vila, M.; Blasco, J. Immunotoxicity of Polystyrene Nanoplastics in Different Hemocyte Subpopulations of Mytilus galloprovincialis. Sci. Rep. 2020, 10, 8637. [Google Scholar] [CrossRef] [PubMed]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Le Goïc, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canonico, B.; Papa, S.; Borello, A.; Vezzulli, L.; Canesi, L. Shift in Immune Parameters after Repeated Exposure to Nanoplastics in the Marine Bivalve Mytilus. Front. Immunol. 2020, 11, 426. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Balbi, T.; Salis, A.; Damonte, G.; Cortese, K.; Caratto, V.; Monopoli, M.P.; Dawson, K.; et al. Interactions of cationic polystyrene nanoparticles with marine bivalve hemocytes in a physiological environment: Role of soluble hemolymph proteins. Environ. Res. 2016, 150, 73–81. [Google Scholar] [CrossRef]
- Gheorghe, S.; Stoica, C.; Vasile, G.G.; Nita-Lazar, M.; Stanescu, E.; Lucaciu, I.E. Metals Toxic Effects in Aquatic Ecosystems: Modulators of Water Quality. In Water Quality; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.-J.; Liao, W.; Yan, Z.-F.; Bai, Y.-C.; Feng, C.-L.; Xu, Z.-X.; Xu, D.-Y. Progress in the Research of the Toxicity Effect Mechanisms of Heavy Metals on Freshwater Organisms and Their Water Quality Criteria in China. J. Chem. 2020, 2020, 9010348. [Google Scholar] [CrossRef]
- Baby, J.; Raj, J.S.; Biby, E.T.; Sankarganesh, P.; Jeevitha, M.V.; Ajisha, S.U.; Rajan, S. Toxic effect of heavy metals on aquatic environment. Int. J. Biol. Chem. Sci. 2011, 4, 939–952. [Google Scholar] [CrossRef] [Green Version]
- Banaee, M.; Impellitteri, F.; Samani, H.E.-Z.; Piccione, G.; Faggio, C. Dietary Arthrospira platensis in Rainbow Trout (Oncorhynchus mykiss): A Means to Reduce Threats Caused by CdCl2 Exposure? Toxics 2022, 10, 731. [Google Scholar] [CrossRef]
- Tresnakova, N.; Famulari, S.; Zicarelli, G.; Impellitteri, F.; Pagano, M.; Presti, G.; Filice, M.; Caferro, A.; Gulotta, E.; Guiliano, S.; et al. Multi-characteristic toxicity of enantioselective chiral fungicide tebuconazole to a model organism Mediterranean mussel Mytilus galloprovincialis Lamarck, 1819 (Bivalve: Mytilidae). Sci. Total Environ. 2022, 160874. [Google Scholar] [CrossRef]
- Covert, S.A.; Shoda, M.E.; Stackpoole, S.M.; Stone, W.W. Pesticide mixtures show potential toxicity to aquatic life in U.S. streams, water years 2013–2017. Sci. Total Environ. 2020, 745, 141285. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.; Burgeot, T.; Renault, T. Pacific Oyster (Crassostrea Gigas) Hemocyte Are Not Affected by a Mixture of Pesticides in Short-Term in Vitro Assays. Environ. Sci. Pollut. Res. 2014, 21, 4940–4949. [Google Scholar] [CrossRef]
- Gan, N.; Martin, L.; Xu, W. Impact of Polycyclic Aromatic Hydrocarbon Accumulation on Oyster Health. Front. Physiol. 2021, 12, 734463. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.; Wedderburn, R.; Clarke, K.; McFadzen, I.; Lowe, D.; Readman, J. Emergent synergistic lysosomal toxicity of chemical mixtures in molluscan blood cells (hemocytes). Environ. Pollut. 2018, 235, 1006–1014. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hemolymph Parameters | Adductor Median | Ventricle Median | S | p |
|---|---|---|---|---|
| Cell count (µL) | 1070 | 710 | 4.0 | 0.046 |
| Na (mmol L–1) | 16 | 16 | 0.0 | 1.0 |
| K (mmol L–1) | 0.53 | 0.53 | 1.0 | 0.317 |
| Cl (mmol L–1) | 18 | 18 | 0.0 | 1.0 |
| P (mg dL–1) | 0.85 | 0.80 | 0.33 | 0.564 |
| Ca (mg dL–1) | 17.50 | 18.95 | 4.0 | 0.046 |
| Mg (mg dL–1) | 2.75 | 2.80 | 2.0 | 0.157 |
| NH3 (µmol L–1) | 36.3 | 46.1 | 1.0 | 0.317 |
| Protein (mg dL–1) | 68.2 | 67.9 | 0.0 | 1.0 |
| Class of Xenobiotics | Species | Ecosystem | LMS 1 | THC 1 | Phagocytosis | Biochemical Parameters | Hemocyte Viability and Apoptosis | Refs. |
|---|---|---|---|---|---|---|---|---|
| Heavy metals | Mytilus galloprovincialis | Marine ecosystem | ↓ | ↓ | [26] | |||
| Orconectes propinquus | Freshwater ecosystem | ↓ Na+, Ca2+ | [27] | |||||
| Portunus pelagicus | Marine ecosystem | ↑ Glucose (after 24 h) | [28] | |||||
| Mytilus edulis | Marine ecosystem | ↑ (10−9 to 10−7 M) | [29] | |||||
| Tapes philippinarum | Marine ecosystem | ↓ | ↓ | [30] | ||||
| Perna canaliculus | Marine ecosystem | Sign. altered 25 metabolites | [31] | |||||
| Crassostrea rivularis | Marine ecosystem | ↓ | ↑ | [32] | ||||
| Dreissena polymorpha | Brackish, freshwater ecosystem | ↓ | ↓ Cell viability | [33] | ||||
| Crassostrea virginica; | Marine ecosystem | ↓ Cell viability; ↑ Apoptosis; | [34] | |||||
| Mytilus edulis | Marine ecosystem | ↓ | ↓Cell viability | [35] | ||||
| Sinopotamon henanense | Freshwater ecosystem | ↓ | ↓ Protein content | [36] | ||||
| Palaemon elegans | Brackish, marine ecosystem | ↓ | [37] | |||||
| Lamellidens marginalis | Freshwater ecosystem | ↓ | ↓ Cell viability; ↑ Apoptosis; | [38] | ||||
| Elliptio complanata | Freshwater ecosystem | ↑ Ca2+; ↓ Na+ No effect on Cl- and K+ | [39] | |||||
| Pesticides | Bellamya bengalensis | Freshwater ecosystem | ↑ | [40] | ||||
| Litopenaeus vannamei | Brackish, marine ecosystem | Sign. difference | [41] | |||||
| Ruditapes philippinarum | Marine ecosystem | ↓ | [42] | |||||
| Haliotis tuberculate | Marine ecosystem | No effect | No effect | [43] | ||||
| Eriocheir sinensis | Marine, brackish, freshwater ecosystem | ↓ | ↓ | [44] | ||||
| Anodonta anatine; Lymnea stagnalis | Freshwater ecosystem | ↓ | [45,46] | |||||
| Cardisoma armatum | Terrestrial and marine ecosystem | ↓ | [47] | |||||
| Biomphalaria glabrata; Planobarius corneus | Freshwater ecosystem | No sign. effect | No sign. effect | ↑ | [48] | |||
| Paratelphusa jacquemontii | Freshwater ecosystem | ↓ | ↓ | [49] | ||||
| Mytilus galloprovincialis | Marine ecosystem | ↑ Ca2+, NH3, Mg2+, glucose; ↓ PHOS, urea, Cl−, K+ | [2,5] | |||||
| Hydrocarbons and oil spills | Crassostrea gigas | Marine ecosystem | ↓ | [50] | ||||
| Crassostrea gigas | Marine ecosystem | ↓ | ↓ | [51] | ||||
| Chlamys farreri | Marine ecosystem | ↓ | ↓ | [52] | ||||
| Haliotis diversicolor | Marine ecosystem | ↓ | ↓ | [53] | ||||
| Polystyrene microplastics | Mytilus galloprovincialis | Marine ecosystem | ↓ | ↓ | [54] | |||
| Mytilus galloprovincialis; Mytilus edulis | Marine ecosystem | ↓ | ↑ | [55] | ||||
| Mytilus edulis | Marine ecosystem | No sign. difference | No sign. difference | [56] | ||||
| Mytilus edulis | Marine ecosystem | No sign. effect | No sign. effect | No sign. effect | [57] | |||
| Mytilus galloprovincialis | Marine ecosystem | ↓ | ↓ Cell viability; ↑ Apoptosis; | [58] | ||||
| Crassostrea gigas | Marine ecosystem | ↑ Hemocytes size | [59] | |||||
| Amino-modified nanopolystyrene (PS-NH2) | Mytilus galloprovincialis | Marine ecosystem | No sign. effect | No sign. effect | [60] | |||
| Mytilus galloprovincialis | Marine ecosystem | ↓ | ↓ | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Impellitteri, F.; Curpăn, A.-S.; Plăvan, G.; Ciobica, A.; Faggio, C. Hemocytes: A Useful Tool for Assessing the Toxicity of Microplastics, Heavy Metals, and Pesticides on Aquatic Invertebrates. Int. J. Environ. Res. Public Health 2022, 19, 16830. https://doi.org/10.3390/ijerph192416830
Impellitteri F, Curpăn A-S, Plăvan G, Ciobica A, Faggio C. Hemocytes: A Useful Tool for Assessing the Toxicity of Microplastics, Heavy Metals, and Pesticides on Aquatic Invertebrates. International Journal of Environmental Research and Public Health. 2022; 19(24):16830. https://doi.org/10.3390/ijerph192416830
Chicago/Turabian StyleImpellitteri, Federica, Alexandrina-Stefania Curpăn, Gabriel Plăvan, Alin Ciobica, and Caterina Faggio. 2022. "Hemocytes: A Useful Tool for Assessing the Toxicity of Microplastics, Heavy Metals, and Pesticides on Aquatic Invertebrates" International Journal of Environmental Research and Public Health 19, no. 24: 16830. https://doi.org/10.3390/ijerph192416830
APA StyleImpellitteri, F., Curpăn, A. -S., Plăvan, G., Ciobica, A., & Faggio, C. (2022). Hemocytes: A Useful Tool for Assessing the Toxicity of Microplastics, Heavy Metals, and Pesticides on Aquatic Invertebrates. International Journal of Environmental Research and Public Health, 19(24), 16830. https://doi.org/10.3390/ijerph192416830