
Effects of Sulfide Input on Arsenate Bioreduction and Its Reduction Product Formation in Sulfidic Groundwater


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Sediment Samples
2.2. Bacterial Isolation and Identification
2.3. Batch Experiments
2.3.1. Bioreduction Products of Arsenate in the Presence of Sulfide
2.3.2. Effect of Initial Sulfide Concentration and pH on Arsenate Bioreduction
2.3.3. Effect of the Ratio of Sulfur to Arsenic on Arsenate Bioreduction Products
2.4. Chemical Analysis
3. Results
3.1. Phylogenetic Analysis
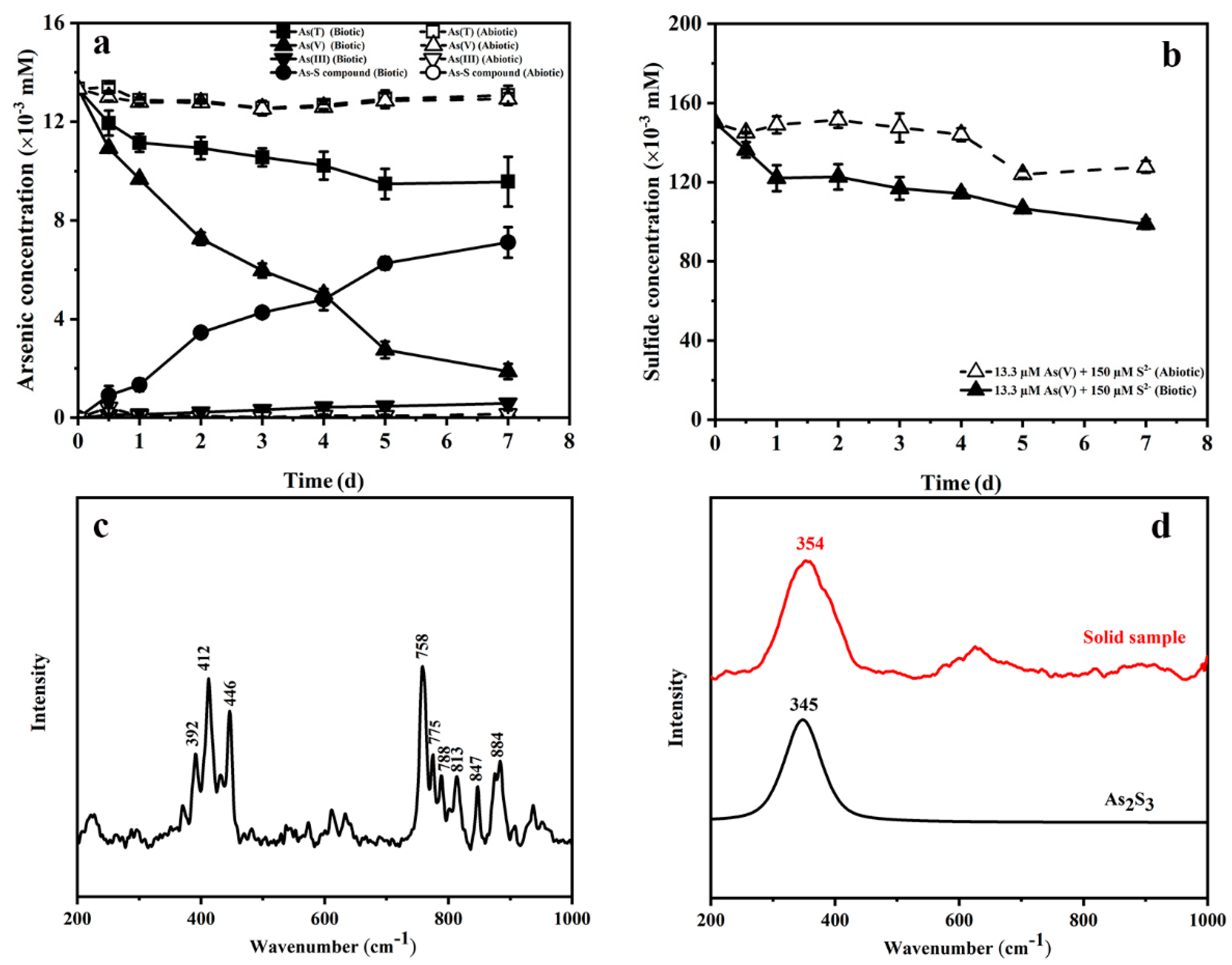
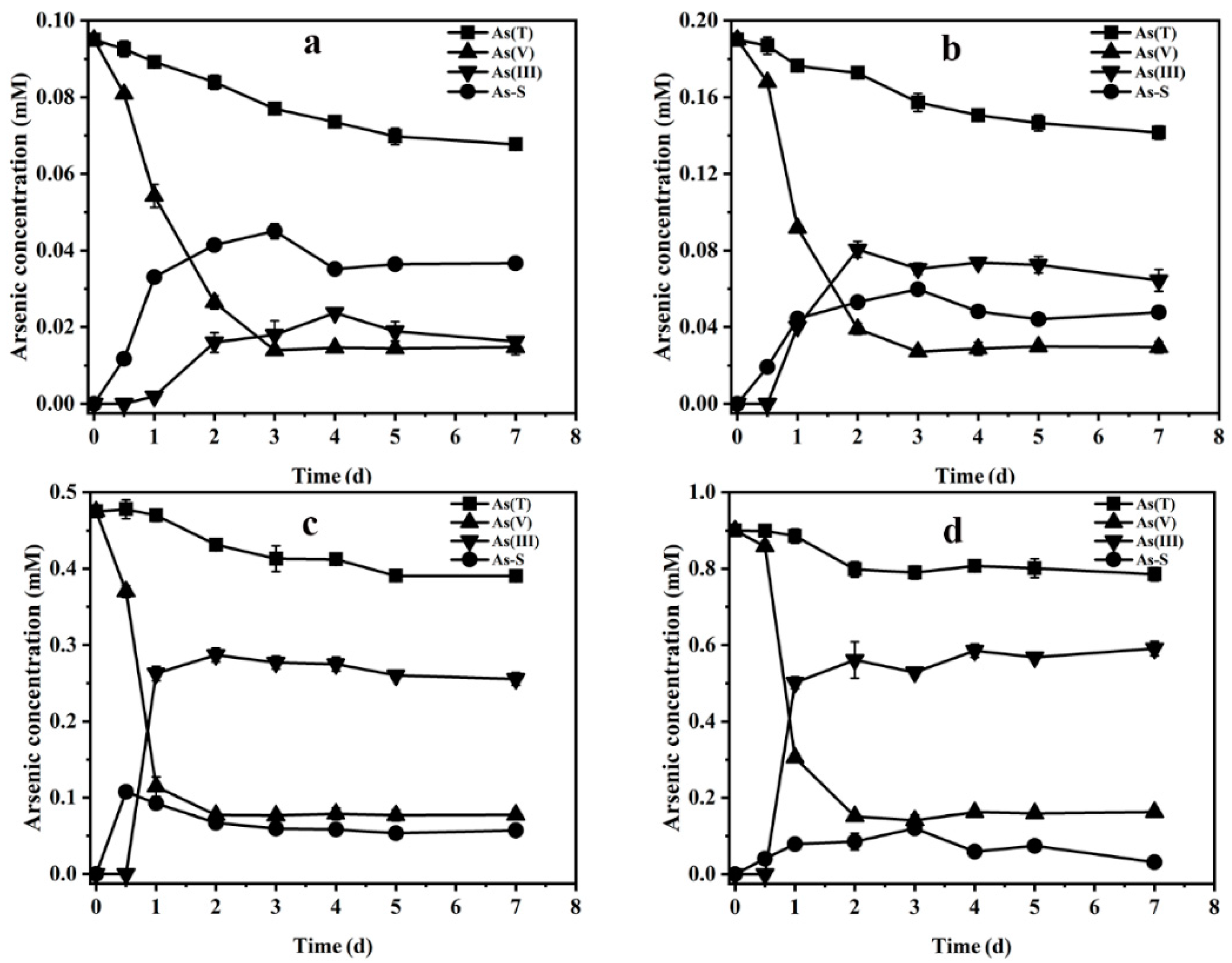
3.2. Bioreduction Products of Arsenate in the Presence of Sulfide
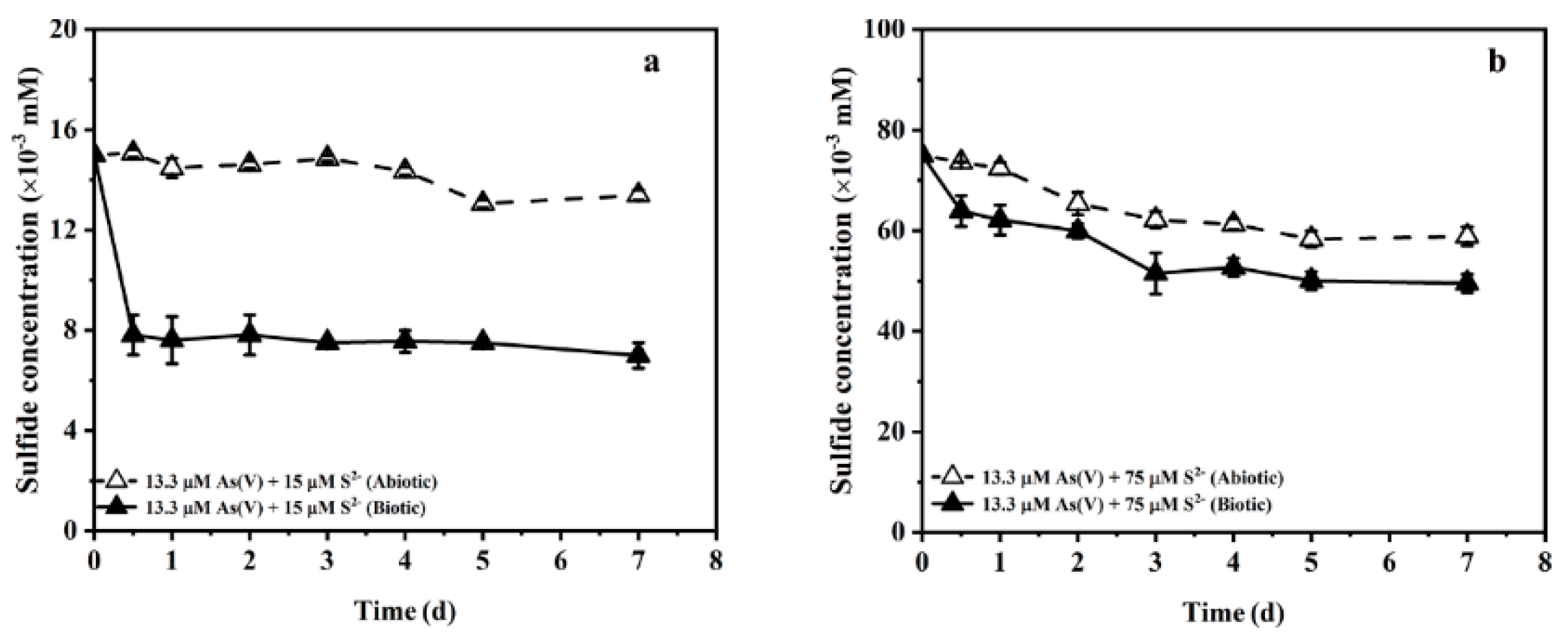
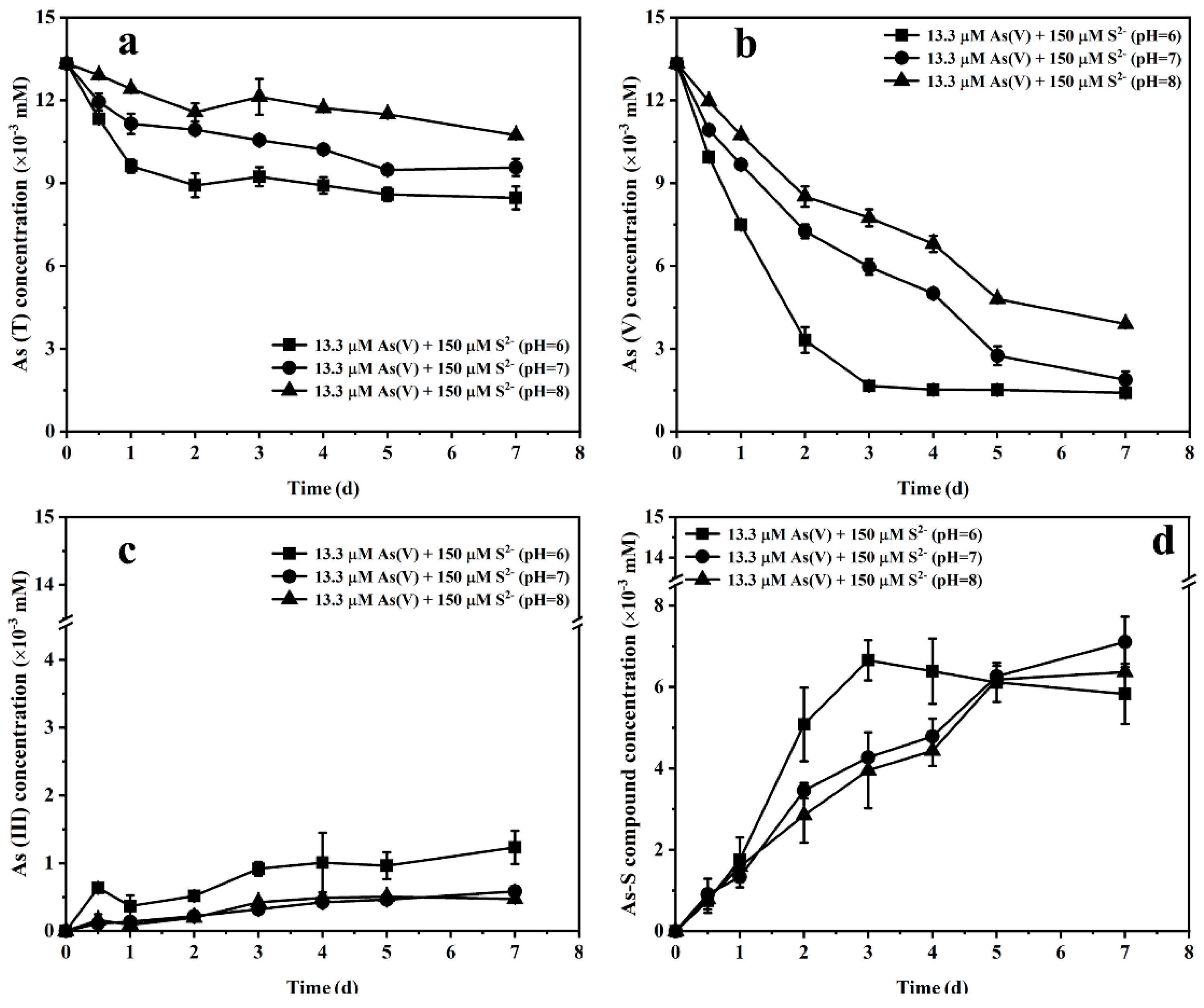
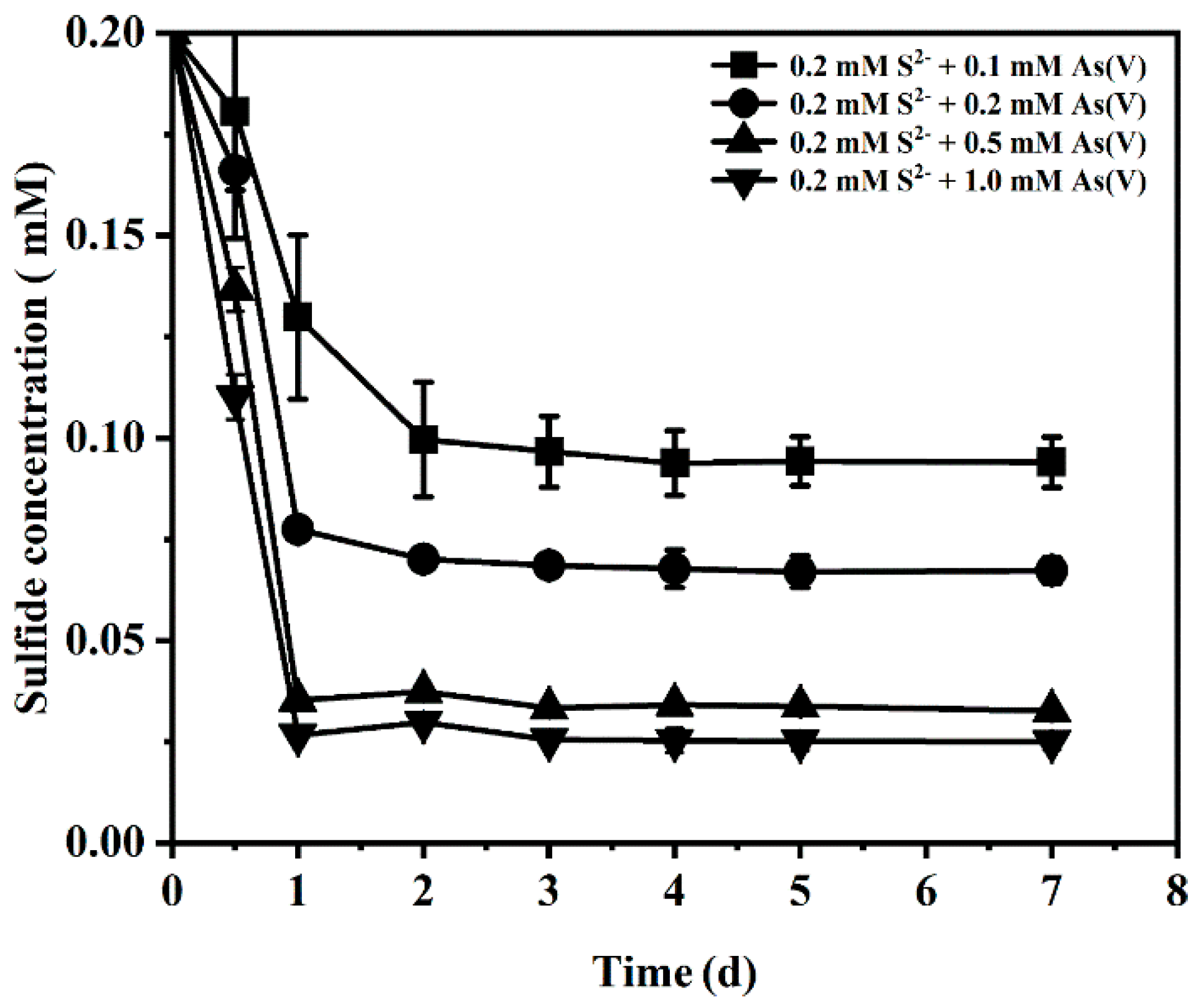
3.3. Effect of Initial Sulfide Concentration and pH on Arsenate Bioreduction
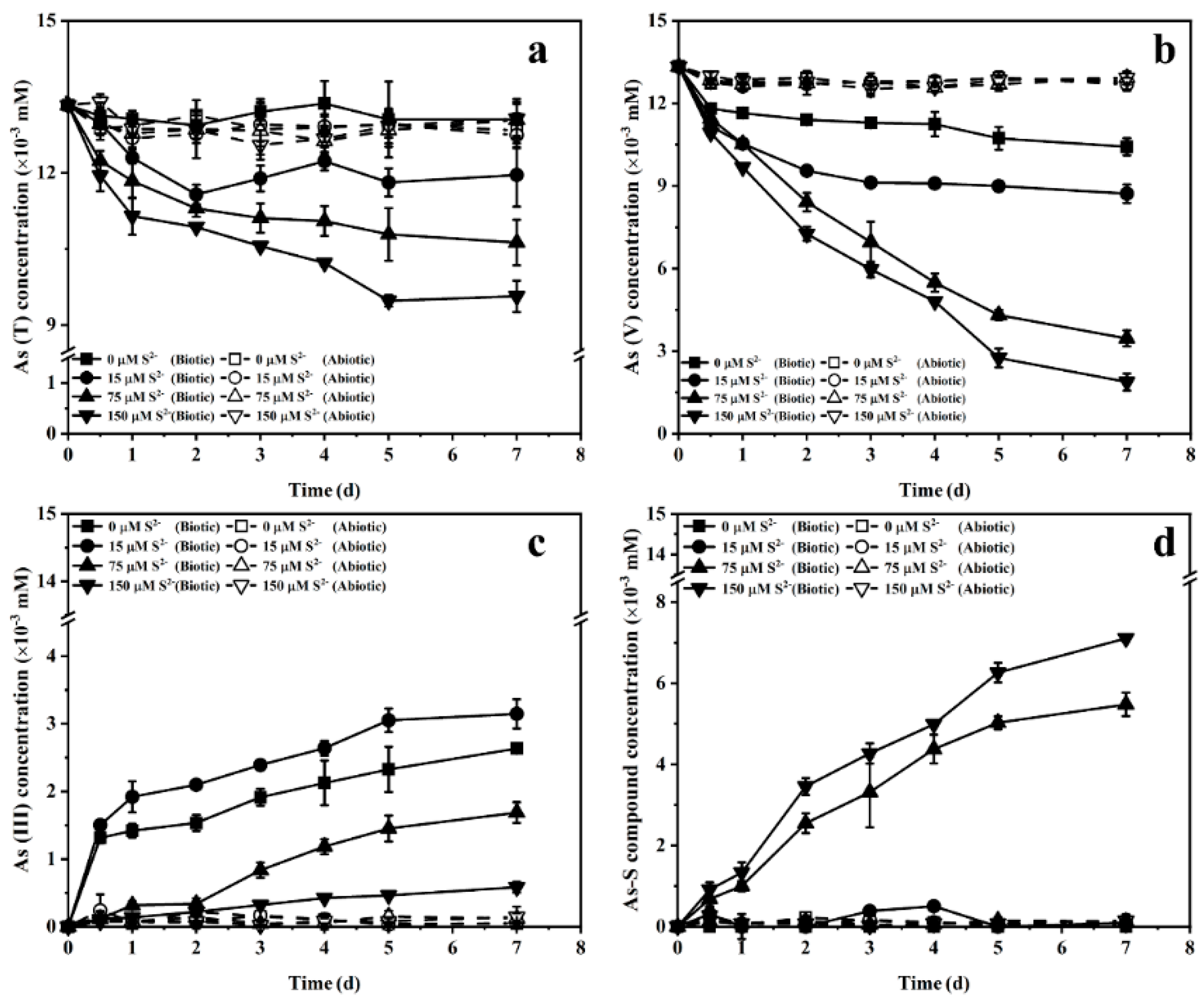
3.4. Effect of the Ratio of Sulfur to Arsenic on Arsenate Bioreduction Products
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhowmick, S.; Pramanik, S.; Singh, P.; Mondal, P.; Chatterjee, D.; Nriagu, J. Arsenic in groundwater of West Bengal, India: A review of human health risks and assessment of possible intervention options. Sci. Total Environ. 2018, 612, 148–169. [Google Scholar] [CrossRef] [PubMed]
- Palma-Lara, I.; Martínez-Castillo, M.; Quintana-Pérez, J.; Arellano-Mendoza, M.; Tamay-Cach, F.; Valenzuela-Limón, O.; García-Montalvo, E.; Hernández-Zavala, A. Arsenic exposure: A public health problem leading to several cancers. Regul. Toxicol. Pharm. 2020, 110, 104539. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, P.; Tu, J.; Zhang, R.; Wei, D.; Li, B.; Wang, Y.; Jiang, Z. Evidence of arsenic mobilization mediated by an indigenous iron reducing bacterium from high arsenic groundwater aquifer in Hetao Basin of Inner Mongolia, China. Int. Biodeterior. Biodegrad. 2018, 128, 22–27. [Google Scholar] [CrossRef]
- Zheng, T.; Deng, Y.; Wang, Y.; Jiang, H.; Xie, X.; Gan, Y. Microbial sulfate reduction facilitates seasonal variation of arsenic concentration in groundwater of Jianghan Plain, Central China. Sci. Total Environ. 2020, 735, 139327. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wen, D.; Liu, Z.; Jia, Y.; Guo, Q. A review of high arsenic groundwater in Mainland and Taiwan, China: Distribution, characteristics and geochemical processes. Appl. Geochem. 2014, 41, 196–217. [Google Scholar] [CrossRef]
- Islam, F.S.; Gault, A.G.; Boothman, C.; Polya, D.A.; Charnock, J.M.; Chatterjee, D.; Lloyd, J.R. Role of metal-reducing bacteria in arsenic release from Bengal delta sediments. Nature 2004, 430, 68–71. [Google Scholar] [CrossRef]
- Guo, H.; Liu, Z.; Ding, S.; Hao, C.; Xiu, W.; Hou, W. Arsenate reduction and mobilization in the presence of indigenous aerobic bacteria obtained from high arsenic aquifers of the Hetao basin, Inner Mongolia. Environ. Pollut. 2015, 203, 50–59. [Google Scholar] [CrossRef]
- Jia, M.R.; Tang, N.; Cao, Y.; Chen, Y.; Han, Y.H.; Ma, L.Q. Efficient arsenate reduction by As-resistant bacterium Bacillus sp. strain PVR-YHB1-1: Characterization and genome analysis. Chemosphere 2019, 218, 1061–1070. [Google Scholar] [CrossRef]
- Bhattacharjee, H.; Rosen, B.P. Arsenic metabolism in prokaryotic and eukaryotic microbes. In Molecular Microbiology of Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2007; pp. 371–406. [Google Scholar] [CrossRef]
- Oremland, R.S.; Stolz, J.F. The ecology of arsenic. Science 2003, 300, 939–944. [Google Scholar] [CrossRef]
- Saltikov, C.W.; Wildman, R.A.; Newman, D.K. Expression dynamics of arsenic respiration and detoxification in Shewanella sp. strain ANA-3. J. Bacteriol. 2005, 187, 7390–7396. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xie, Z.; Wei, X.; Chen, M.; Luo, Y.; Wang, Y. An indigenous bacterium Bacillus XZM for phosphate enhanced transformation and migration of arsenate. Sci. Total Environ. 2020, 719, 137183. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Meng, X.; Jing, C. Influence of sulfur on the mobility of arsenic and antimony during oxic-anoxic cycles: Differences and competition. Geochim. Cosmochim. Acta. 2020, 288, 51–67. [Google Scholar] [CrossRef]
- Qiu, Y.Y.; Guo, J.H.; Zhang, L.; Chen, G.H.; Jiang, F. A high-rate sulfidogenic process based on elemental sulfur reduction: Cost-effectiveness evaluation and microbial community analysis. Biochem. Eng. J. 2017, 128, 26–32. [Google Scholar] [CrossRef]
- Sun, J.; Quicksall, A.N.; Chillrud, S.N.; Mailloux, B.J.; Bostick, B.C. Arsenic mobilization from sediments in microcosms under sulfate reduction. Chemosphere 2016, 153, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; He, J.; Xie, X.; Wang, Y.; Li, J.; Qian, K.; Deng, Y.; Gan, Y. The effect of groundwater velocities on sulfidation of arsenic-bearing ferrihydrite: Insight from column experiments. J. Hazard. Mater. 2020, 586, 124827. [Google Scholar] [CrossRef]
- Pi, K.; Wang, Y.; Xie, X.; Ma, T.; Su, C.; Liu, Y. Role of sulfur redox cycling on arsenic mobilization in aquifers of Datong Basin, northern China. Appl. Geochem. 2017, 77, 31–43. [Google Scholar] [CrossRef]
- Alam, R.; McPhedran, K. Applications of biological sulfate reduction for remediation of arsenic–A review. Chemosphere 2019, 222, 932–944. [Google Scholar] [CrossRef]
- Briones-Gallardo, R.; Escot-Espinoza, V.M.; Cervantes-González, E. Removing arsenic and hydrogen sulfide production using arsenic-tolerant sulfate-reducing bacteria. Int. J. Environ. Sci. Technol. 2017, 14, 609–622. [Google Scholar] [CrossRef]
- Le Pape, P.; Battaglia-Brunet, F.; Parmentier, M.; Joulian, C.; Gassaud, C.; Fernandez-Rojo, L.; Morin, G. Complete removal of arsenic and zinc from a heavily contaminated acid mine drainage via an indigenous SRB consortium. J. Hazard. Mater. 2017, 321, 764–772. [Google Scholar] [CrossRef]
- Deng, Y.; Zheng, T.; Wang, Y.; Liu, L.; Jiang, H.; Ma, T. Effect of microbially mediated iron mineral transformation on temporal variation of arsenic in the Pleistocene aquifers of the central Yangtze River basin. Sci. Total Environ. 2018, 619, 1247–1258. [Google Scholar] [CrossRef]
- Wan, M.; Schröder, C.; Peiffer, S. Fe (III): S (-II) concentration ratio controls the pathway and the kinetics of pyrite formation during sulfidation of ferric hydroxides. Geochim. Cosmochim. Acta 2017, 217, 334–348. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Chatterjee, S.; Gupta, D.K. Potential role of microbes in bioremediation of arsenic. Arsenic Contamination in the Environment; Springer: Cham, Switzerland, 2017; pp. 195–213. [Google Scholar] [CrossRef]
- Kulp, T.R.; Hoeft, S.E.; Asao, M.; Madigan, M.T.; Hollibaugh, J.T.; Fisher, J.C.; Stolz, J.F.; Culbertson, C.W.; Miller, L.G.; Oremland, R.S. Arsenic (III) fuels anoxygenic photosynthesis in hot spring biofilms from Mono Lake, California. Science 2008, 321, 967–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeft, S.E.; Kulp, T.R.; Stolz, J.F.; Hollibaugh, J.T.; Oremland, R.S. Dissimilatory arsenate reduction with sulfide as electron donor: Experiments with Mono Lake water and isolation of strain MLMS-1, a chemoautotrophic arsenate respirer. Appl. Environ. Microb. 2004, 70, 2741–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollibaugh, J.T.; Carini, S.; Gürleyük, H.; Jellison, R.; Joye, S.B.; LeCleir, G.; Meile, C.; Vasquez, L.; Wallschläger, D. Arsenic speciation in Mono Lake, California: Response to seasonal stratification and anoxia. Geochim. Cosmochim. Acta 2005, 69, 1925–1937. [Google Scholar] [CrossRef]
- Rochette, E.A.; Bostick, B.C.; Li, G.; Fendorf, S. Kinetics of arsenate reduction by dissolved sulfide. Environ. Sci. Technol. 2000, 34, 4714–4720. [Google Scholar] [CrossRef]
- Bostick, B.C.; Fendorf, S.; Brown Jr, G.E. In situ analysis of thioarsenite complexes in neutral to alkaline arsenic sulphide solutions. Mineral. Mag. 2005, 69, 781–795. [Google Scholar] [CrossRef]
- Planer-Friedrich, B.; Suess, E.; Scheinost, A.C.; Wallschläger, D. Arsenic speciation in sulfidic waters: Reconciling contradictory spectroscopic and chromatographic evidence. Anal. Chem. 2010, 82, 10228–10235. [Google Scholar] [CrossRef]
- Xie, Z.; Luo, Y.; Wang, Y.; Xie, X.; Su, C. Arsenic resistance and bioaccumulation of an indigenous bacterium isolated from aquifer sediments of Datong Basin, Northern China. Geomicrobiol. J. 2013, 30, 549–556. [Google Scholar] [CrossRef]
- Le, X.C.; Yalcin, S.; Ma, M. Speciation of submicrogram per liter levels of arsenic in water: On-site species separation integrated with sample collection. Environ. Sci. Technol. 2000, 34, 2342–2347. [Google Scholar] [CrossRef]
- Reese, B.K.; Finneran, D.W.; Mills, H.J.; Zhu, M.X.; Morse, J.W. Examination and refinement of the determination of aqueous hydrogen sulfide by the methylene blue method. Aquat. Geochem. 2011, 17, 567–582. [Google Scholar] [CrossRef]
- Blum, J.S.; Hernandez-Maldonado, J.; Redford, K.; Sing, C.; Bennett, S.C.; Saltikov, C.W.; Oremland, R.S. Arsenate-dependent growth is independent of an ArrA mechanism of arsenate respiration in the termite hindgut isolate Citrobacter sp. strain TSA-1. Can. J. Microbiol. 2018, 64, 619–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suess, E.; Planer-Friedrich, B. Thioarsenate formation upon dissolution of orpiment and arsenopyrite. Chemosphere 2012, 89, 1390–1398. [Google Scholar] [CrossRef]
- Sun, J.; Hong, Y.; Guo, J.; Yang, J.; Huang, D.; Lin, Z.; Jiang, F. Arsenite removal without thioarsenite formation in a sulfidogenic system driven by sulfur reducing bacteria under acidic conditions. Water Res. 2019, 151, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Huang, L.; Jiang, H.; Peng, Y.E.; Guo, W.; Chen, Z.; She, W.; Guo, Q.; Dong, H. Thioarsenate formation coupled with anaerobic arsenite oxidation by a sulfate-reducing bacterium isolated from a hot spring. Front. Microbiol. 2017, 8, 1336. [Google Scholar] [CrossRef] [Green Version]
- Helz, G.R.; Tossell, J.A.; Charnock, J.M.; Pattrick, R.A.; Vaughan, D.J.; Garner, C.D. Oligomerization in As (III) sulfide solutions: Theoretical constraints and spectroscopic evidence. Geochim. Cosmochim. Acta 1995, 59, 4591–4604. [Google Scholar] [CrossRef]
- Wood, S.A.; Tait, C.D.; Janecky, D.R. A Raman spectroscopic study of arsenite and thioarsenite species in aqueous solution at 25 C. Geochemical Trans. 2002, 3, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Cassone, G.; Chillè, D.; Giacobello, F.; Giuffrè, O.; Mollica Nardo, V.; Ponterio, R.C.; Saija, F.; Sponer, J.; Trusso, S.; Foti, C. Interaction between As (III) and simple thioacids in water: An experimental and ab initio molecular dynamics investigation. J. Phys. Chem. B 2019, 123, 6090–6098. [Google Scholar] [CrossRef]
- Burton, E.D.; Johnston, S.G.; Planer-Friedrich, B. Coupling of arsenic mobility to sulfur transformations during microbial sulfate reduction in the presence and absence of humic acid. Chem. Geol. 2013, 343, 12–24. [Google Scholar] [CrossRef]
- Price, R.; Planer-Friedrich, B.; Pichler, T.; Jean, J.; Bundschuh, J.; Bhattacharya, P. Arsenic cycling, thioarsenates and orpiment precipitation at a shallow sea hydrothermal system, Milos Island, Greece. In Arsenic in Geosphere and Human Diseases: As 2010; CRC Press: Boca Raton, FL, USA, 2010; pp. 66–68. [Google Scholar] [CrossRef]
- Stauder, S.; Raue, B.; Sacher, F. Thioarsenates in sulfidic waters. Environ. Sci. Technol. 2005, 39, 5933–5939. [Google Scholar] [CrossRef]
- Oremland, R.S.; Kulp, T.R.; Blum, J.S.; Hoeft, S.E.; Baesman, S.; Miller, L.G.; Stolz, J.F. A microbial arsenic cycle in a salt-saturated, extreme environment. Science 2005, 308, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Ter Heijne, A.; de Rink, R.; Liu, D.; Klok, J.B.; Buisman, C.J. Bacteria as an electron shuttle for sulfide oxidation. Environ. Sci. Technol. Lett. 2018, 5, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocar, B.D.; Borch, T.; Fendorf, S. Arsenic repartitioning during biogenic sulfidization and transformation of ferrihydrite. Geochim. Cosmochim. Acta 2010, 74, 980–994. [Google Scholar] [CrossRef]
- Wilkin, R.T.; Wallschläger, D.; Ford, R.G. Speciation of arsenic in sulfidic waters. Geochem. Trans. 2003, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, X.; Wang, S.; Xiao, F.; Zhang, D.; Yao, S.; Jia, Y. The adsorption behavior of thioarsenite on magnetite and ferrous sulfide. Chem. Geol. 2018, 492, 1–11. [Google Scholar] [CrossRef]
- Wolthers, M.; Butler, I.B.; Rickard, D. Influence of arsenic on iron sulfide transformations. Chem. Geol. 2007, 236, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.J.; Chen, C.; Guo, H.L.; Wang, A.J.; Ren, N.Q.; Lee, D.J. Characterization of a newly isolated strain Pseudomonas sp. C27 for sulfide oxidation: Reaction kinetics and stoichiometry. Sci. Rep. 2016, 6, 21032. [Google Scholar] [CrossRef]
- Sun, S.; Xie, X.; Li, J.; Qian, K.; Chi, Z. Distribution and formation of thioarsenate in high arsenic groundwater from the Datong Basin, northern China. J. Hydrol. 2020, 590, 125268. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xie, X.; Chen, M.; Xie, Z.; Wang, J. Effects of Sulfide Input on Arsenate Bioreduction and Its Reduction Product Formation in Sulfidic Groundwater. Int. J. Environ. Res. Public Health 2022, 19, 16987. https://doi.org/10.3390/ijerph192416987
Yang Y, Xie X, Chen M, Xie Z, Wang J. Effects of Sulfide Input on Arsenate Bioreduction and Its Reduction Product Formation in Sulfidic Groundwater. International Journal of Environmental Research and Public Health. 2022; 19(24):16987. https://doi.org/10.3390/ijerph192416987
Chicago/Turabian StyleYang, Yang, Xi Xie, Mengna Chen, Zuoming Xie, and Jia Wang. 2022. "Effects of Sulfide Input on Arsenate Bioreduction and Its Reduction Product Formation in Sulfidic Groundwater" International Journal of Environmental Research and Public Health 19, no. 24: 16987. https://doi.org/10.3390/ijerph192416987
APA StyleYang, Y., Xie, X., Chen, M., Xie, Z., & Wang, J. (2022). Effects of Sulfide Input on Arsenate Bioreduction and Its Reduction Product Formation in Sulfidic Groundwater. International Journal of Environmental Research and Public Health, 19(24), 16987. https://doi.org/10.3390/ijerph192416987