
Early Morphological Changes of the Rectus Femoris Muscle and Deep Fascia in Ullrich Congenital Muscular Dystrophy

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
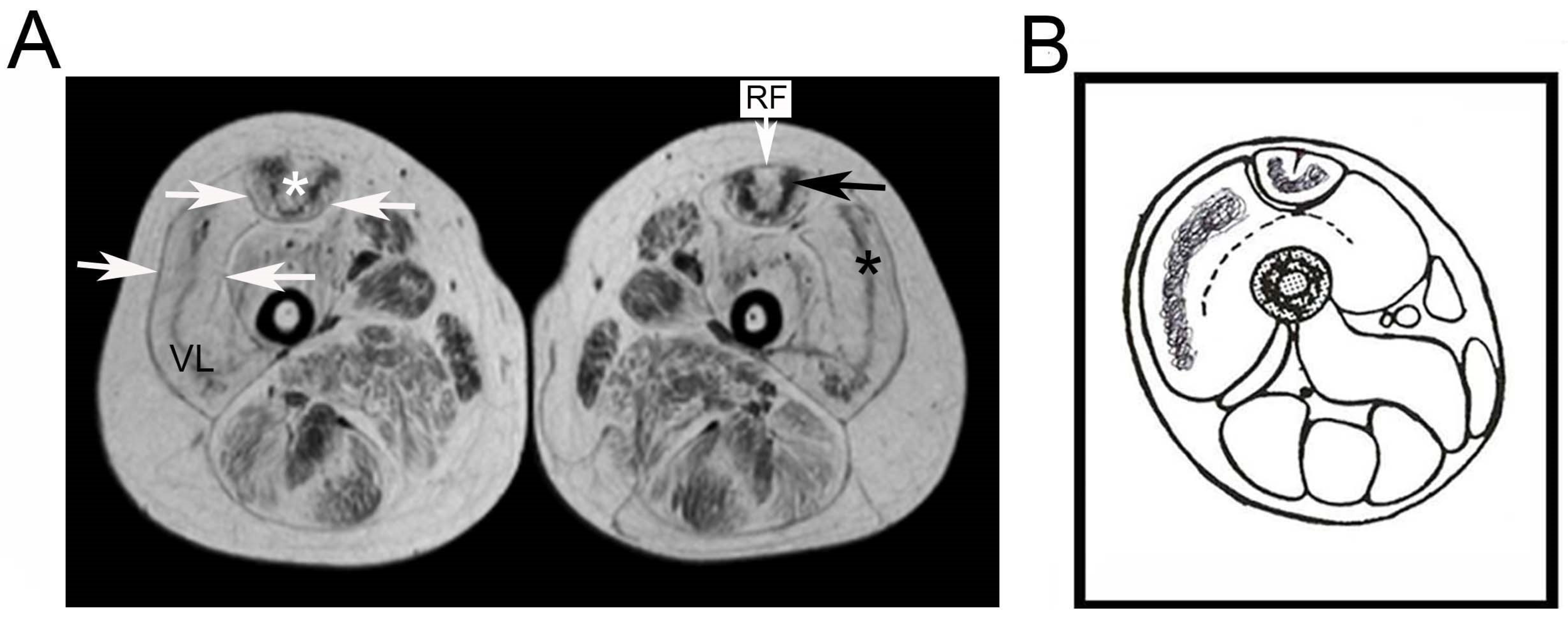
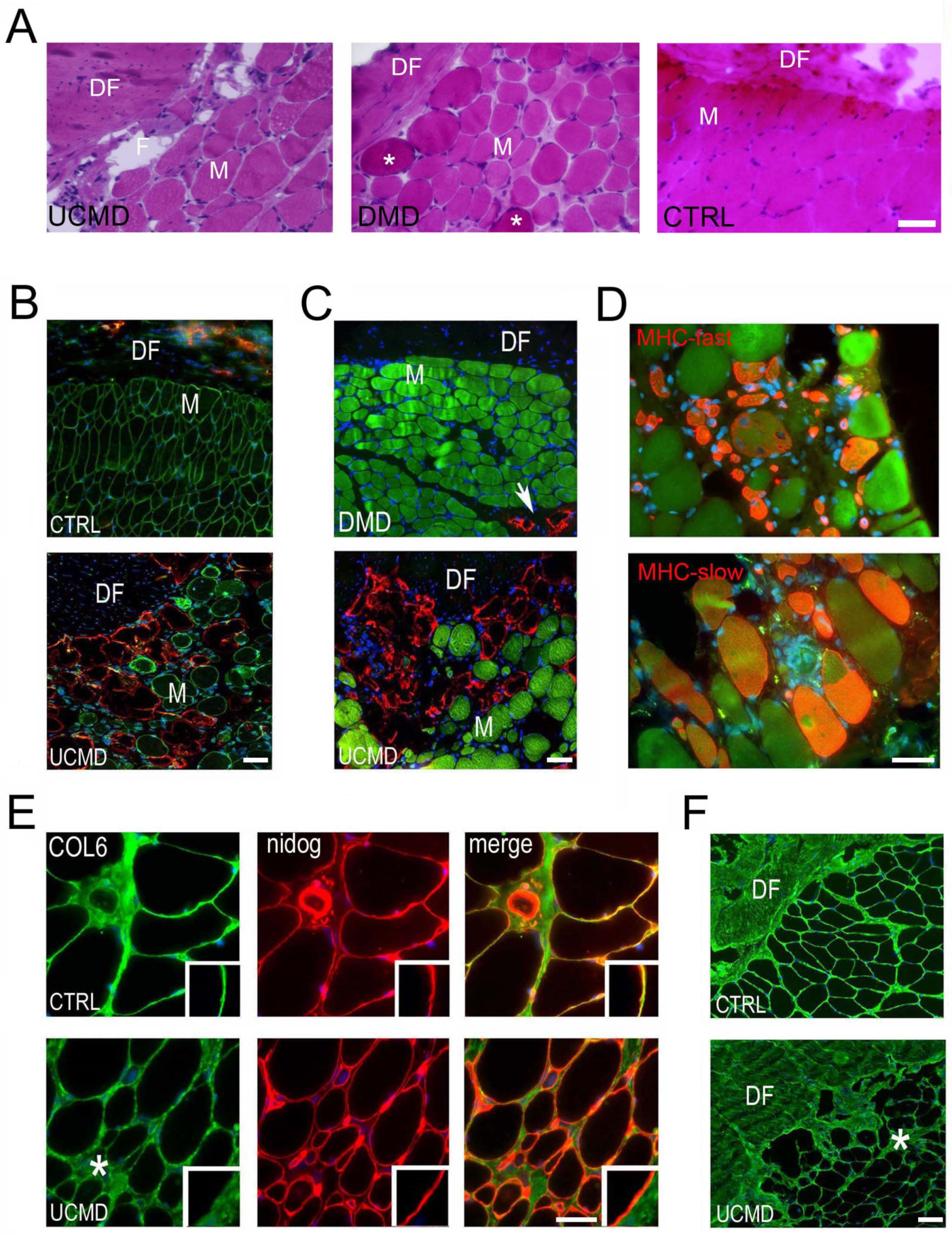
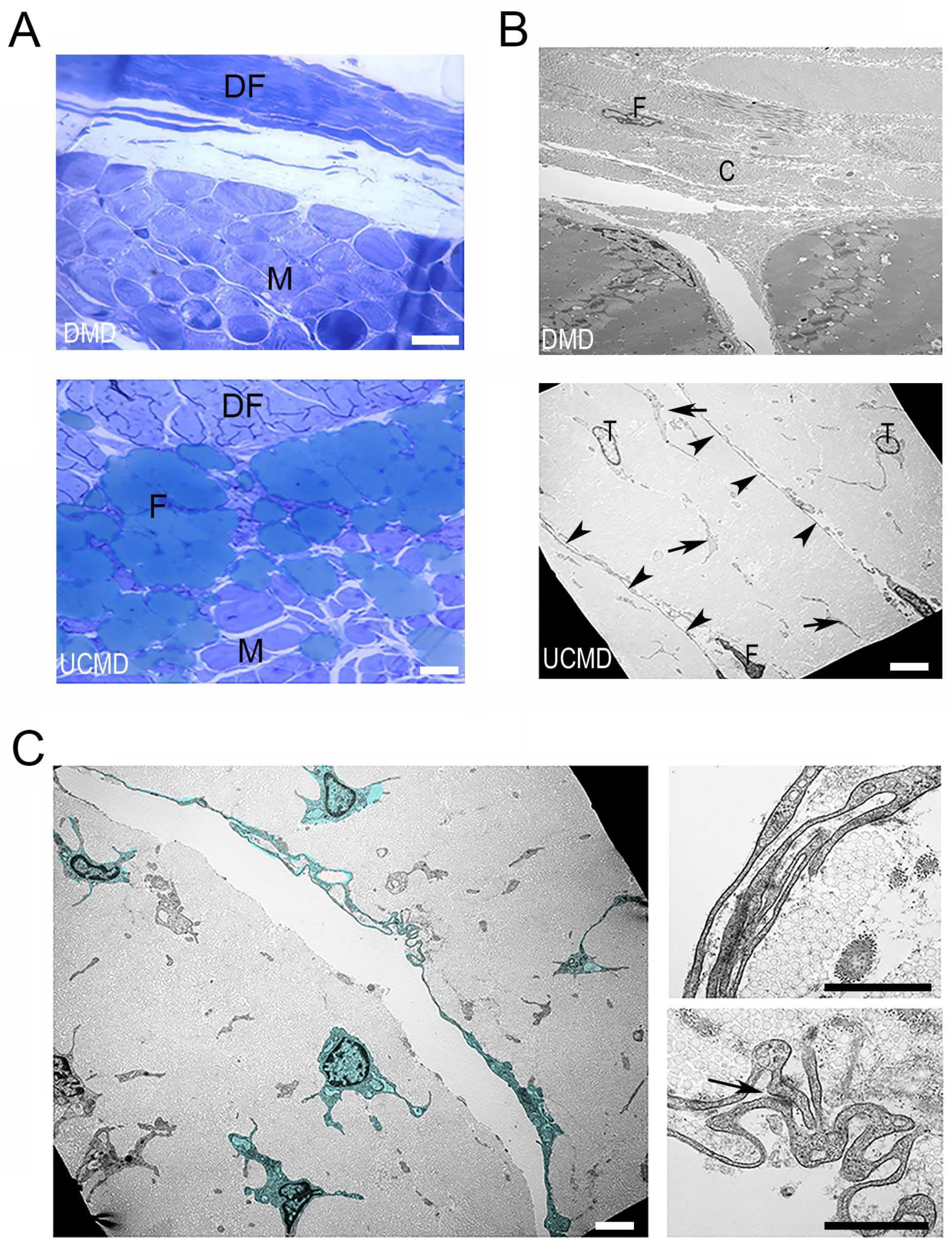
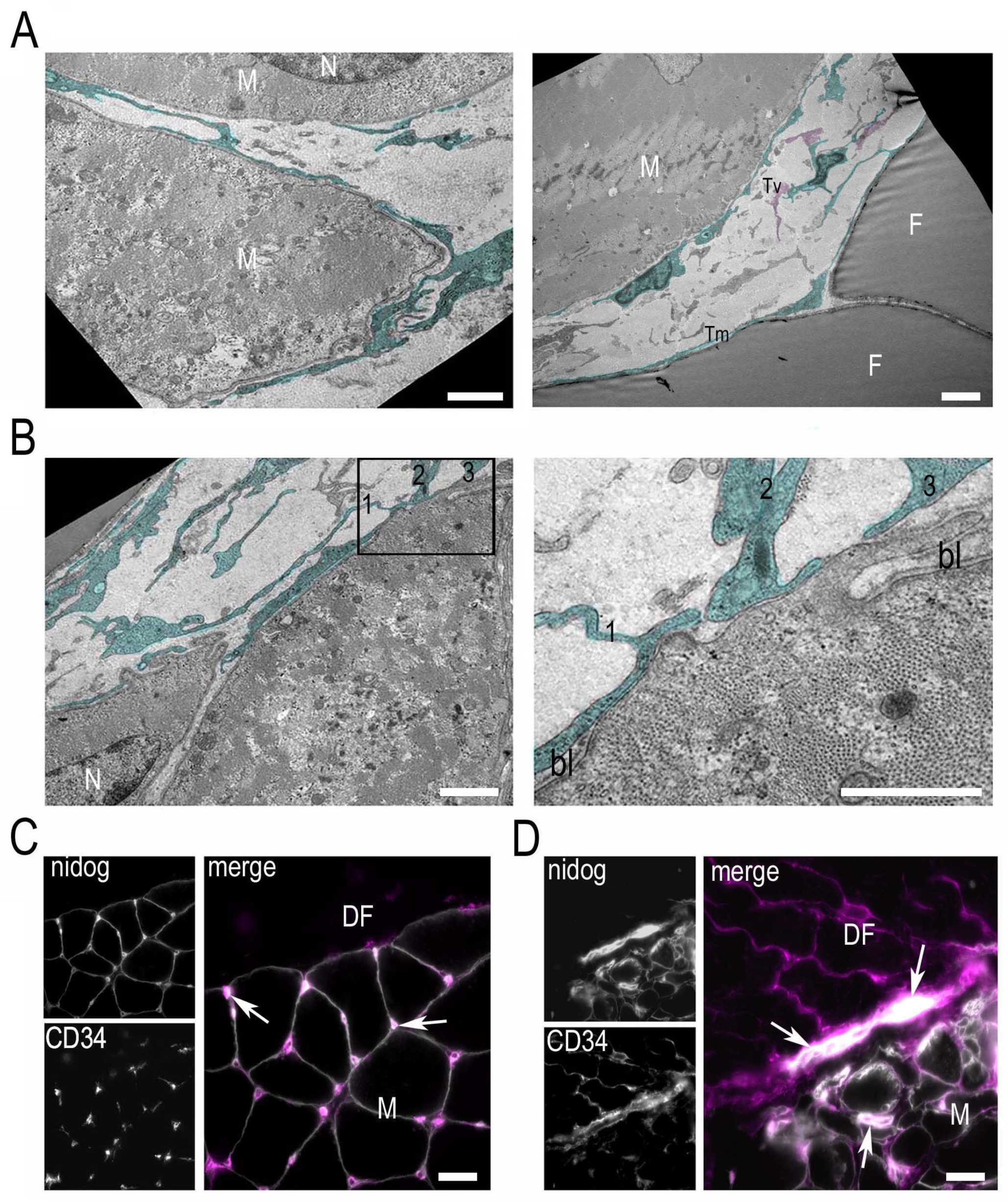
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cescon, M.; Gattazzo, F.; Chen, P.; Bonaldo, P. Collagen VI at a glance. J. Cell Sci. 2015, 128, 3525–3531. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, J.; Rich, C.; Zhou, F.H.; Hansen, U. Three novel collagen VI chains, alpha4(VI), alpha5(VI), and alpha6(VI). J. Biol. Chem. 2008, 283, 20170–20180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatelli, P.; Gualandi, F.; Gara, S.K.; Grumati, P.; Zamparelli, A.; Martoni, E.; Pellegrini, C.; Merlini, L.; Ferlini, A.; Bonaldo, P.; et al. Expression of collagen VI alpha5 and alpha6 chains in human muscle and in duchenne muscular dystrophy-related muscle fibrosis. Matrix Biol. 2012, 31, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruns, R.R.; Press, W.; Engvall, E.; Timpl, R.; Gross, J. Type VI collagen in extracellular, 100-nm periodic filaments and fibrils: Identification by immunoelectron microscopy. J. Cell Biol. 1986, 103, 393–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiberg, C.; Heinegard, D.; Wenglen, C.; Timpl, R.; Morgelin, M. Biglycan organizes collagen VI into hexagonal-like networks resembling tissue structures. J. Biol. Chem. 2002, 277, 49120–49126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlini, L.; Faldini, C.; Bonaldo, P. Editorial: Muscle-tendon-innervation unit: Degeneration and aging-pathophysiological and regeneration mechanisms. Front. Aging Neurosci. 2016, 8, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fede, C.; Pirri, C.; Fan, C.; Petrelli, L.; Guidolin, D.; de Caro, R.; Stecco, C. A closer look at the cellular and molecular components of the deep/muscular fasciae. Int. J. Mol. Sci. 2021, 22, 1411. [Google Scholar] [CrossRef] [PubMed]
- Gatt, A.; Agarwal, S.; Zito, P.M. Anatomy, Fascia Layers. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Pirri, C.; Fede, C.; Petrelli, L.; Guidolin, D.; Fan, C.; de Caro, R.; Stecco, C. An anatomical comparison of the fasciae of the thigh: A macroscopic, microscopic and ultrasound imaging study. J. Anat. 2021, 238, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Sardone, F.; Santi, S.; Tagliavini, F.; Traina, F.; Merlini, L.; Squarzoni, S.; Cescon, M.; Wagener, R.; Maraldi, N.M.; Bonaldo, P.; et al. Collagen VI-NG2 axis in human tendon fibroblasts under conditions mimicking injury response. Matrix Biol. 2016, 55, 90–105. [Google Scholar] [CrossRef]
- Kumka, M.; Bonar Fascia, J. A morphological description and classification system based on a literature review. J. Can. Chiropr. Assoc. 2012, 56, 179–191. [Google Scholar] [PubMed]
- Merlini, L.; Bernardi, P. Therapy of collagen vi-related myopathies (Bethlem and Ullrich). Neurotherapeutics 2008, 5, 613–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, A.K.; Bushby, K.M. Collagen VI related muscle disorders. J. Med. Genet. 2005, 42, 673–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho Vanegas, O.; Bertini, E.; Zhang, R.Z.; Petrini, S.; Minosse, C.; Sabatelli, P.; Giusti, B.; Chu, M.L.; Pepe, G. Ullrich scleroatonic muscular dystrophy is caused by recessive mutations in collagen type VI. Proc. Natl. Acad. Sci. USA 2001, 98, 7516–7521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepe, G.; Bertini, E.; Bonaldo, P.; Bushby, K.; Giusti, B.; de Visser, M.; Guicheney, P.; Lattanzi, G.; Merlini, L.; Muntoni, F.; et al. Bethlem myopathy (BETHLEM) and Ullrich scleroatonic muscular dystrophy: 100th ENMC international workshop, 23–24 November 2001, Naarden, the Netherlands. Neuromuscul. Disord. 2002, 12, 984–993. [Google Scholar] [CrossRef]
- Gualandi, F.; Urciuolo, A.; Martoni, E.; Sabatelli, P.; Squarzoni, S.; Bovolenta, M.; Messina, S.; Mercuri, E.; Franchella, A.; Ferlini, A.; et al. Autosomal recessive Bethlem myopathy. Neurology 2009, 73, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Sugie, K.; Murayama, K.; Awaya, A.; Suzuki, Y.; Noguchi, S.; Hayashi, Y.K.; Nonaka, I.; Nishino, I. Ullrich disease due to deficiency of collagen VI in the sarcolemma. Neurology 2004, 62, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zhang, R.Z.; Sabatelli, P.; Chu, M.L.; Bonnemann, C.G. Muscle interstitial fibroblasts are the main source of collagen VI synthesis in skeletal muscle: Implications for congenital muscular dystrophy types Ullrich and Bethlem. J. Neuropathol. Exp. Neurol. 2008, 67, 144–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Mallebrera, C.; Maioli, M.A.; Kim, J.; Brown, S.C.; Feng, L.; Lampe, A.K.; Bushby, K.; Hicks, D.; Flanigan, K.M.; Bonnemann, C.; et al. A comparative analysis of collagen VI production in muscle, skin and fibroblasts from 14 Ullrich congenital muscular dystrophy patients with dominant and recessive COL6A mutations. Neuromuscul. Disord. 2006, 16, 571–582. [Google Scholar] [CrossRef]
- Pepe, G.; de Visser, M.; Bertini, E.; Bushby, K.; Vanegas, O.C.; Chu, M.L.; Lattanzi, G.; Merlini, L.; Muntoni, F.; Urtizberea, A. Bethlem Myopathy (BETHLEM) In Proceedings of the 86th ENMC International Workshop, Naarden, The Netherlands, 10–11 November 2000.
- Bonnemann, C.G.; Brockmann, K.; Hanefeld, F. Muscle ultrasound in Bethlem myopathy. Neuropediatrics 2003, 34, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Liu, A.; Wei, C.; Yang, H.; Chang, X.; Wang, S.; Yuan, Y.; Bonnemann, C.; Wu, Q.; Wu, X.; et al. Genetic and clinical findings in a Chinese cohort of patients with collagen VI-related myopathies. Clin. Genet. 2018, 93, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Shin, H.Y.; Park, H.J.; Kim, S.H.; Kim, S.M.; Choi, Y.C. Clinical, pathologic, and genetic features of collagen VI-related myopathy in Korea. J. Clin. Neurol. 2017, 13, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercuri, E.; Bushby, K.; Ricci, E.; Birchall, D.; Pane, M.; Kinali, M.; Allsop, J.; Nigro, V.; Saenz, A.; Nascimbeni, A.; et al. Muscle MRI findings in patients with limb girdle muscular dystrophy with calpain 3 deficiency (LGMD2A) and early contractures. Neuromuscul. Disord. 2005, 15, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Panades-de Oliveira, L.; Rodriguez-Lopez, C.; Montenegro, D.C.; Toledano, M.D.M.M.; Fernandez-Marmiesse, A.; Perez, J.E.; Lain, A.H.; Dominguez-Gonzalez, C. Bethlem myopathy: A series of 16 patients and description of seven new associated mutations. J. Neurol. 2019, 266, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Yonekawa, T.; Nishino, I. Ullrich congenital muscular dystrophy: Clinicopathological features, natural history and pathomechanism(s). J. Neurol. Neurosurg. Psychiatry 2015, 86, 280–287. [Google Scholar] [CrossRef]
- Irwin, W.A.; Bergamin, N.; Sabatelli, P.; Reggiani, C.; Megighian, A.; Merlini, L.; Braghetta, P.; Columbaro, M.; Volpin, D.; Bressan, G.M.; et al. Mitochondrial dysfunction and apoptosis in myopathic mice with collagen VI deficiency. Nat. Genet. 2003, 35, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Grumati, P.; Coletto, L.; Sabatelli, P.; Cescon, M.; Angelin, A.; Bertaggia, E.; Blaauw, B.; Urciuolo, A.; Tiepolo, T.; Merlini, L.; et al. Autophagy is defective in collagen vi muscular dystrophies, and its reactivation rescues myofiber degeneration. Nat. Med. 2010, 16, 1313–1320. [Google Scholar] [CrossRef]
- Zulian, A.; Rizzo, E.; Schiavone, M.; Palma, E.; Tagliavini, F.; Blaauw, B.; Merlini, L.; Maraldi, N.M.; Sabatelli, P.; Braghetta, P.; et al. NIM811, a cyclophilin inhibitor without immunosuppressive activity, is beneficial in collagen VI congenital muscular dystrophy models. Hum. Mol. Genet. 2014, 23, 5353–5363. [Google Scholar] [CrossRef] [Green Version]
- Angelin, A.; Tiepolo, T.; Sabatelli, P.; Grumati, P.; Bergamin, N.; Golfieri, C.; Mattioli, E.; Gualandi, F.; Ferlini, A.; Merlini, L.; et al. Mitochondrial dysfunction in the pathogenesis of Ullrich congenital muscular dystrophy and prospective therapy with cyclosporins. Proc. Natl. Acad. Sci. USA 2007, 104, 991–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnaro, S.; Pellegrini, C.; Pellegrini, M.; Chrisam, M.; Sabatelli, P.; Toni, S.; Grumati, P.; Ripamonti, C.; Pratelli, L.; Maraldi, M.N.; et al. Autophagy activation in COL6 myopathic patients by a low-protein-diet pilot trial. Autophagy 2016, 12, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Mercuri, E.; Lampe, A.; Allsop, J.; Knight, R.; Pane, M.; Kinali, M.; Bonnemann, C.; Flanigan, K.; Lapini, I.; Bushby, K.; et al. Muscle MRI in Ullrich congenital muscular dystrophy and Bethlem myopathy. Neuromuscul. Disord. 2005, 15. [Google Scholar] [CrossRef] [PubMed]
- Tagliavini, F.; Pellegrini, C.; Sardone, F.; Squarzoni, S.; Paulsson, M.; Wagener, R.; Gualandi, F.; Trabanelli, C.; Ferlini, A.; Merlini, L.; et al. Defective collagen VI alpha6 chain expression in the skeletal muscle of patients with collagen VI-related myopathies. Biochim. Biophys. Acta 2014, 1842, 1604–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schessl, J.; Goemans, N.M.; Magold, A.I.; Zou, Y.; Hu, Y.; Kirschner, J.; Sciot, R.; Bonnemann, C.G. Predominant fiber atrophy and fiber type disproportion in early Ullrich disease. Muscle Nerve 2008, 38, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Paco, S.; Ferrer, I.; Jou, C.; Cusi, V.; Corbera, J.; Torner, F.; Gualandi, F.; Sabatelli, P.; Orozco, A.; Gomez-Foix, A.M.; et al. Muscle fiber atrophy and regeneration coexist in collagen VI-deficient human muscle: Role of calpain-3 and nuclear factor-kappaB signaling. J. Neuropathol. Exp. Neurol. 2012, 71, 894–906. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, I.; Horikiri, T.; Niiyama, T.; Suehara, M.; Shiraishi, T.; Hu, J.; Uchida, Y.; Saito, A.; Nakagawa, M.; Arimura, K.; et al. Pathological characteristics of skeletal muscle in Ullrich’s disease with collagen VI deficiency. Neuromuscul. Disord. 2003, 13, 310–316. [Google Scholar] [CrossRef]
- Wang, Y.; Pessin, J.E. Mechanisms for fiber-type specificity of skeletal muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Grumati, P.; Coletto, L.; Schiavinato, A.; Castagnaro, S.; Bertaggia, E.; Sandri, M.; Bonaldo, P. Physical exercise stimulates autophagy in normal skeletal muscles but is detrimental for collagen VI-deficient muscles. Autophagy 2011, 7, 1415–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urciuolo, A.; Quarta, M.; Morbidoni, V.; Gattazzo, F.; Molon, S.; Grumati, P.; Montemurro, F.; Tedesco, F.S.; Blaauw, B.; Cossu, G.; et al. Collagen VI regulates satellite cell self-renewal and muscle regeneration. Nat. Commun. 2013, 4, 1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlini, L.; Angelin, A.; Tiepolo, T.; Braghetta, P.; Sabatelli, P.; Zamparelli, A.; Ferlini, A.; Maraldi, N.M.; Bonaldo, P.; Bernardi, P. Cyclosporin A corrects mitochondrial dysfunction and muscle apoptosis in patients with collagen VI myopathies. Proc. Natl. Acad. Sci. USA 2008, 105, 5225–5229. [Google Scholar] [CrossRef] [Green Version]
- Merlini, L.; Sabatelli, P.; Armaroli, A.; Gnudi, S.; Angelin, A.; Grumati, P.; Michelini, M.E.; Franchella, A.; Gualandi, F.; Bertini, E.; et al. Cyclosporine A in Ullrich congenital muscular dystrophy: Long-term results. Oxid. Med. Cell. Longev. 2011, 2011, 139194. [Google Scholar] [CrossRef] [PubMed]
- Fiacco, E.; Castagnetti, F.; Bianconi, V.; Madaro, L.; de Bardi, M.; Nazio, F.; D’Amico, A.; Bertini, E.; Cecconi, F.; Puri, P.L.; et al. Autophagy regulates satellite cell ability to regenerate normal and dystrophic muscles. Cell Death Differ. 2016, 23, 1839–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.E.; Bareja, A.; Bartlett, D.B.; White, J.P. Autophagy as a therapeutic target to enhance aged muscle regeneration. Cells 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawidowicz, J.; Szotek, S.; Matysiak, N.; Mielanczyk, L.; Maksymowicz, K. Electron microscopy of human fascia lata: Focus on telocytes. J. Cell. Mol. Med. 2015, 19, 2500–2506. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G. The telocytes: Ten years after their introduction in the scientific literature. An update on their morphology, distribution, and potential roles in the gut. Int. J. Mol. Sci. 2020, 21, 4478. [Google Scholar] [CrossRef]
- Rosa, I.; Marini, M.; Manetti, M. Telocytes: An emerging component of stem cell niche microenvironment. J. HistoChem. CytoChem. 2021, 69, 795–818. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Li, D.; Chen, Y.; Li, Y.; Huang, R.; Zhu, K.; Chen, H.; Yuan, Z.; Zheng, X.; Zhao, H.; et al. Early moderate exercise benefits myocardial infarction healing via improvement of inflammation and ventricular remodelling in rats. J. Cell. Mol. Med. 2019, 23, 8328–8342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manetti, M.; Tani, A.; Rosa, I.; Chellini, F.; Squecco, R.; Idrizaj, E.; Zecchi-Orlandini, S.; Ibba-Manneschi, L.; Sassoli, C. Morphological evidence for telocytes as stromal cells supporting satellite cell activation in eccentric contraction-induced skeletal muscle injury. Sci. Rep. 2019, 9, 14515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deconinck, N.; Dan, B. Pathophysiology of Duchenne muscular dystrophy: Current hypotheses. Pediatr. Neurol. 2007, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabatelli, P.; Merlini, L.; Di Martino, A.; Cenni, V.; Faldini, C. Early Morphological Changes of the Rectus Femoris Muscle and Deep Fascia in Ullrich Congenital Muscular Dystrophy. Int. J. Environ. Res. Public Health 2022, 19, 1252. https://doi.org/10.3390/ijerph19031252
Sabatelli P, Merlini L, Di Martino A, Cenni V, Faldini C. Early Morphological Changes of the Rectus Femoris Muscle and Deep Fascia in Ullrich Congenital Muscular Dystrophy. International Journal of Environmental Research and Public Health. 2022; 19(3):1252. https://doi.org/10.3390/ijerph19031252
Chicago/Turabian StyleSabatelli, Patrizia, Luciano Merlini, Alberto Di Martino, Vittoria Cenni, and Cesare Faldini. 2022. "Early Morphological Changes of the Rectus Femoris Muscle and Deep Fascia in Ullrich Congenital Muscular Dystrophy" International Journal of Environmental Research and Public Health 19, no. 3: 1252. https://doi.org/10.3390/ijerph19031252
APA StyleSabatelli, P., Merlini, L., Di Martino, A., Cenni, V., & Faldini, C. (2022). Early Morphological Changes of the Rectus Femoris Muscle and Deep Fascia in Ullrich Congenital Muscular Dystrophy. International Journal of Environmental Research and Public Health, 19(3), 1252. https://doi.org/10.3390/ijerph19031252