
Effects of Acute Yohimbine Hydrochloride Supplementation on Repeated Supramaximal Sprint Performance


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Supplementation
2.4. Blood Collection and Analysis (Lactate, Epinephrine, and Norepinephrine)
2.5. Procedures
2.6. Data Analysis
3. Results
3.1. Mean Power, Peak Power, Total Work, and Fatigue Index
3.2. Heart Rate (HR) and Rate of Perceived Exertion (RPE)
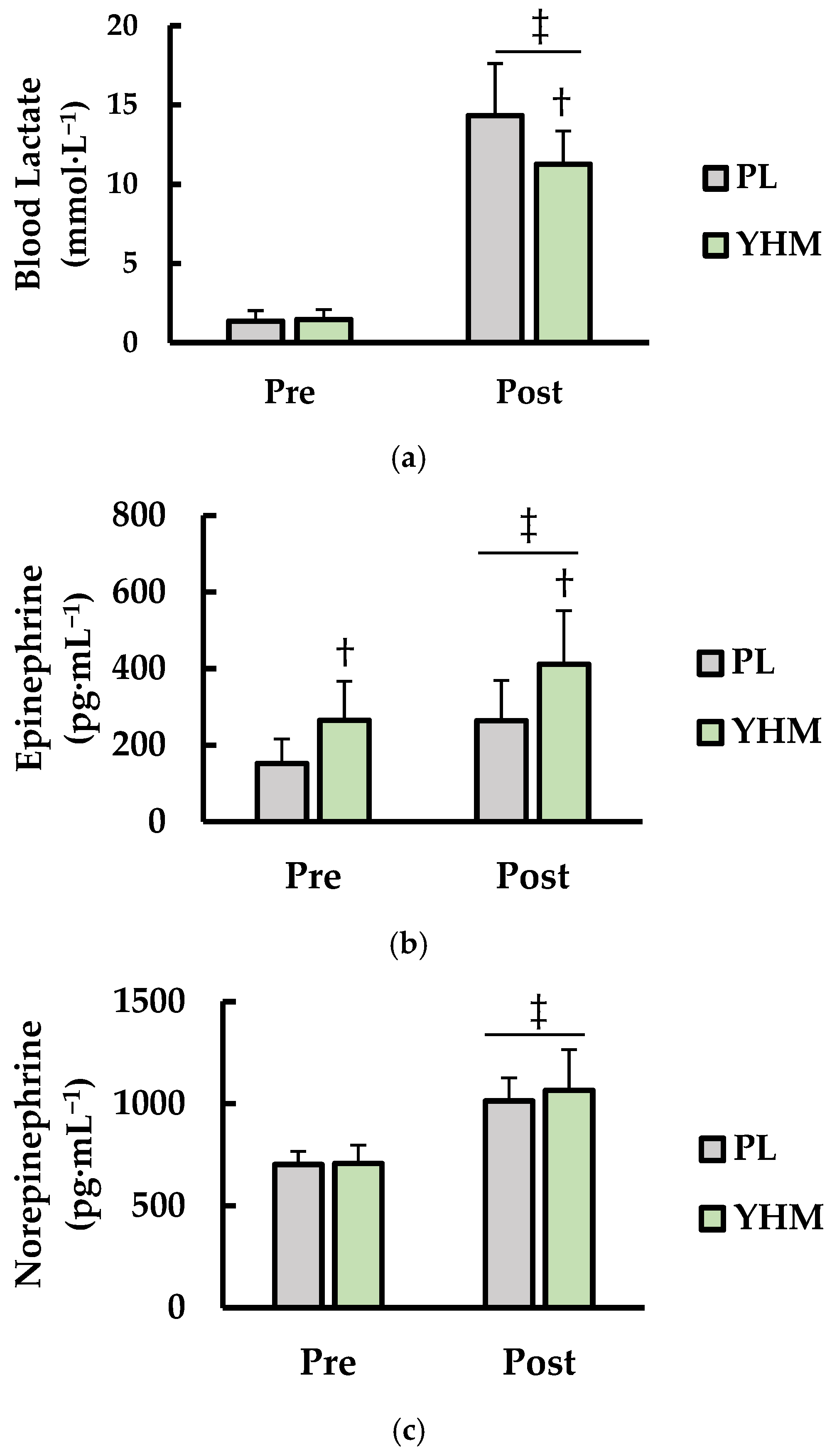
3.3. Blood Lactate (LA), Norepinephrine (NE), and Epinephrine (EPI)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tam, S.W.; Worcel, M.; Wyllie, M. Yohimbine: A clinical review. Pharm. Ther. 2001, 91, 215–243. [Google Scholar] [CrossRef]
- Fu, W.-Q.; Li, W.; Wang, J.-H.; Du, G.-H. Yohimbine. In Natural Small Molecule Drugs from Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 167–171. [Google Scholar]
- Cohen, P.A.; Wang, Y.H.; Maller, G.; DeSouza, R.; Khan, I.A. Pharmaceutical quantities of yohimbine found in dietary supplements in the USA. Drug Test. Anal. 2016, 8, 357–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostojic, S.M. Yohimbine: The effects on body composition and exercise performance in soccer players. Res. Sports Med. 2006, 14, 289–299. [Google Scholar] [CrossRef]
- Cameron, O.G.; Zubieta, J.K.; Grunhaus, L.; Minoshima, S. Effects of yohimbine on cerebral blood flow, symptoms, and physiological functions in humans. Psychosom. Med. 2000, 62, 549–559. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Pre-exercise administration of yohimbine may enhance the efficacy of exercise training as a fat loss strategy by boosting lipolysis. Med. Hypotheses 2002, 58, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.; Nakatsu, S.; Fenemore, J.; Condra, M.; Surridge, D.; Morales, A. The pharmacokinetics of yohimbine in man. Eur. J. Clin. Pharmacol. 1987, 32, 577–582. [Google Scholar] [CrossRef]
- Schwartz, D.; Clark, T. Selectivity of atipamezole, yohimbine and tolazoline for alpha-2 adrenergic receptor subtypes: Implications for clinical reversal of alpha-2 adrenergic receptor mediated sedation in sheep. J. Vet. Pharmacol. Ther. 1998, 21, 342–347. [Google Scholar] [CrossRef]
- Ehlert, U.; Erni, K.; Hebisch, G.; Nater, U. Salivary α-amylase levels after yohimbine challenge in healthy men. J. Clin. Endocrinol. Metab. 2006, 91, 5130–5133. [Google Scholar] [CrossRef]
- Murburg, M.M.; Villacres, E.C.; Ko, G.N.; Veith, R.R. Effects of yohimbine on human sympathetic nervous system function. J. Clin. Endocrinol. Metab. 1991, 73, 861–865. [Google Scholar] [CrossRef]
- Goldberg, M.R.; Hollister, A.S.; Robertson, D. Influence of yohimbine on blood pressure, autonomic reflexes, and plasma catecholamines in humans. Hypertension 1983, 5, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Grossman, E.; Rosenthal, T.; Peleg, E.; Holmes, C.; Goldstein, D.S. Oral yohimbine increases blood pressure and sympathetic nervous outflow in hypertensive patients. J. Cardiovasc. Pharmacol. 1993, 22, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.M.; Critchley, H.D.; Duka, T. The impact of Yohimbine-induced arousal on facets of behavioural impulsivity. Psychopharmacology 2019, 236, 1783–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, E.; Pittler, M. Yohimbine for erectile dysfunction: A systematic review and meta-analysis of randomized clinical trials. J. Urol. 1998, 159, 433–436. [Google Scholar] [CrossRef]
- McMahon, S.; Jenkins, D. Factors affecting the rate of phosphocreatine resynthesis following intense exercise. Sports Med. 2002, 32, 761–784. [Google Scholar] [CrossRef]
- Stallknecht, B.; Vissing, J.; Galbo, H. Lactate production and clearance in exercise. Effects of training. A mini-review. Scand. J. Med. Sci. Sports 1998, 8, 127–131. [Google Scholar] [CrossRef]
- Giampreti, A.; Lonati, D.; Locatelli, C.; Rocchi, L.; Campailla, M.T. Acute neurotoxicity after yohimbine ingestion by a body builder. Clin. Toxicol. 2009, 47, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Anderson, D.; Harre, N.; Wade, N. Case study: Two fatal case reports of acute yohimbine intoxication. J. Anal. Toxicol. 2013, 37, 611–614. [Google Scholar] [CrossRef] [Green Version]
- M Al-Kuraishy, H.; AN Abood, H.; Al-Gareeb, I.A. Ergogenic Effects of Yohimbine: Standardized Cycling Clinical Study. Kerbala J. Med. 2014, 7, 1850–1855. [Google Scholar]
- Stahlnecker, A.C., IV; Brown, G.A. Acute Effects of a Weight Loss Supplement on Resting Metabolic Rate and Anaerobic Exercise Performance. Med. Sci. Sports Exerc. 2006, 38, S403. [Google Scholar] [CrossRef]
- Cimolai, N.; Cimolai, T. Yohimbine use for physical enhancement and its potential toxicity. J. Diet. Suppl. 2011, 8, 346–354. [Google Scholar] [CrossRef]
- Ballmann, C.G.; Maze, S.B.; Wells, A.C.; Marshall, M.M.; Rogers, R.R. Effects of short-term Rhodiola Rosea (Golden Root Extract) supplementation on anaerobic exercise performance. J. Sports Sci. 2019, 37, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Riebe, D.; Ehrman, J.K.; Liguori, G.; Magal, M.; American College of Sports Medicine. ACSM’s Guidelines for Exercise Testing and Prescription; Wolters Kluwer: Macquarie Park, Australia, 2018. [Google Scholar]
- Ballmann, C.G.; Maynard, D.J.; Lafoon, Z.N.; Marshall, M.R.; Williams, T.D.; Rogers, R.R. Effects of Listening to Preferred versus Non-Preferred Music on Repeated Wingate Anaerobic Test Performance. Sports 2019, 7, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karow, M.C.; Rogers, R.R.; Pederson, J.A.; Williams, T.D.; Marshall, M.R.; Ballmann, C.G. Effects of Preferred and Nonpreferred Warm-Up Music on Exercise Performance. Percept. Mot. Ski. 2020, 127, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.D.; Langley, H.N.; Roberson, C.C.; Rogers, R.R.; Ballmann, C.G. Effects of Short-Term Golden Root Extract (Rhodiola rosea) Supplementation on Resistance Exercise Performance. Int. J. Environ. Res. Public Health 2021, 18, 6953. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bae, S.; Kim, Y.; Cho, C.-H.; Kim, S.J.; Kim, Y.-J.; Lee, S.-P.; Kim, H.-R.; Hwang, Y.-I.; Kang, J.S. Vitamin C prevents stress-induced damage on the heart caused by the death of cardiomyocytes, through down-regulation of the excessive production of catecholamine, TNF-α, and ROS production in Gulo (−/−) Vit C-insufficient mice. Free Radic. Biol. Med. 2013, 65, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Bada, A.; Svendsen, J.; Secher, N.; Saltin, B.; Mortensen, S. Peripheral vasodilatation determines cardiac output in exercising humans: Insight from atrial pacing. J. Physiol. 2012, 590, 2051–2060. [Google Scholar] [CrossRef] [PubMed]
- Dumar, A.M.; Huntington, A.F.; Rogers, R.R.; Kopec, T.J.; Williams, T.D.; Ballmann, C.G. Acute Beetroot Juice Supplementation Attenuates Morning-Associated Decrements in Supramaximal Exercise Performance in Trained Sprinters. Int. J. Environ. Res. Public Health 2021, 18, 412. [Google Scholar] [CrossRef]
- Fritz, C.O.; Morris, P.E.; Richler, J.J. Effect size estimates: Current use, calculations, and interpretation. J. Exp. Psychol. Gen. 2012, 141, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum Associates: Hillsdale, MI, USA, 1988. [Google Scholar]
- Kemp, G.; Taylor, D.; Radda, G. Control of phosphocreatine resynthesis during recovery from exercise in human skeletal muscle. NMR Biomed. 1993, 6, 66–72. [Google Scholar] [CrossRef]
- Balfour, D. Neural mechanisms underlying nicotine dependence. Addiction 1994, 89, 1419–1423. [Google Scholar] [CrossRef]
- Lara, B.; Ruiz-Moreno, C.; Salinero, J.J.; Del Coso, J. Time course of tolerance to the performance benefits of caffeine. PLoS ONE 2019, 14, e0210275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, T.; Schwartz, S.; Correll, C.U. Alpha-2 agonists for attention-deficit/hyperactivity disorder in youth: A systematic review and meta-analysis of monotherapy and add-on trials to stimulant therapy. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Constantin-Teodosiu, D.; Howell, S.; Hultman, E.; Greenhaff, P. Creatine ingestion favorably affects performance and muscle metabolism during maximal exercise in humans. Am. J. Physiol. Endocrinol. Metab. 1996, 271, E31–E37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, I.; Wendelin-Saarenhovi, M.; Kaskinoro, K.; Knuuti, J.; Scheinin, M.; Kalliokoski, K.K. Inhibition of α-adrenergic tone disturbs the distribution of blood flow in the exercising human limb. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H163–H172. [Google Scholar] [CrossRef] [Green Version]
- Buckwalter, J.B.; Clifford, P.S. α-Adrenergic vasoconstriction in active skeletal muscles during dynamic exercise. Am. J. Physiol. Heart Circ. Physiol. 1999, 277, H33–H39. [Google Scholar] [CrossRef]
- Murakami, M.; Katsumura, T.; Hamaoka, T.; Osada, T.; Sako, T.; Higuchi, H.; Esaki, K.; Kime, R.; Shimomitsu, T. Effects of epinephrine and lactate on the increase in oxygen consumption of non-exercising skeletal muscle after aerobic exercise. J. Biomed. Opt. 2000, 5, 406–410. [Google Scholar] [CrossRef]
- Musso, N.R.; Vergassola, C.; Pende, A.; Lotti, G. Yohimbine effects on blood pressure and plasma catecholamines in human hypertension. Am. J. Hypertens 1995, 8, 565–571. [Google Scholar] [CrossRef]
- Costello, J.T.; Bieuzen, F.; Bleakley, C.M. Where are all the female participants in Sports and Exercise Medicine research? Eur. J. Sport Sci. 2014, 14, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Cowley, E.S.; Olenick, A.A.; McNulty, K.L.; Ross, E.Z. “Invisible sportswomen”: The sex data gap in sport and exercise science research. Women Sport Phys. Act. J. 2021, 29, 146–151. [Google Scholar] [CrossRef]
- Convertino, V.A. Gender differences in autonomic functions associated with blood pressure regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, R1909–R1920. [Google Scholar] [CrossRef]
- Gratas-Delamarche, A.; Le Cam, R.; Delamarche, P.; Monnier, M.; Koubi, H. Lactate and catecholamine responses in male and female sprinters during a Wingate test. Eur. J. Appl. Physiol. Occup. Physiol. 1994, 68, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Toth, M.J.; Gardner, A.W.; Arciero, P.J.; Calles-Escandon, J.; Poehlman, E.T. Gender differences in fat oxidation and sympathetic nervous system activity at rest and during submaximal exercise in older individuals. Clin. Sci. 1998, 95, 59–66. [Google Scholar] [CrossRef]
- Dart, A.M.; Du, X.-J.; Kingwell, B.A. Gender, sex hormones and autonomic nervous control of the cardiovascular system. Cardiovasc. Res. 2002, 53, 678–687. [Google Scholar] [CrossRef]
- Trudeau, F.; Brisson, G.; Beauregard, F.; Peronnet, F. Exercise-induced increase of plasma lactate is abolished by a pre-exercise epinephrine infusion. Arch. Int. Physiol. Biochim. Biophys. 1994, 102, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Downey, J.M. Regulation of Venous Return and Cardiac Output; Elsevier Academic Press: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Wang, Y.; Yu, X.; Wang, F.; Wang, Y.; Wang, Y.; Li, H.; Lv, X.; Lu, D.; Wang, H. Yohimbine promotes cardiac NE release and prevents LPS-induced cardiac dysfunction via blockade of presynaptic α2A-adrenergic receptor. PLoS ONE 2013, 8, e63622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuptsova, A.; Zaripova, R.; Hisamieva, L.; Biktemirova, R.; Ziyatdinova, N.; Zefirov, T. Yohimbine influence on myocardium contractile activity among newborn rats. Int. J. Adv. Biotech. Res. 2016, 7, 1305–1309. [Google Scholar]
- Roatta, S.; Farina, D. Sympathetic actions on the skeletal muscle. Exerc. Sport Sci. Rev. 2010, 38, 31–35. [Google Scholar] [CrossRef]
- Roatta, S.; Arendt-Nielsen, L.; Farina, D. Sympathetic-induced changes in discharge rate and spike-triggered average twitch torque of low-threshold motor units in humans. J. Physiol. 2008, 586, 5561–5574. [Google Scholar] [CrossRef]
- Tsampoukos, A.; Peckham, E.A.; James, R.; Nevill, M.E. Effect of menstrual cycle phase on sprinting performance. Eur. J. Appl. Physiol. 2010, 109, 659–667. [Google Scholar] [CrossRef]
- Smekal, G.; Von Duvillard, S.P.; Frigo, P.; Tegelhofer, T.; Pokan, R.; Hofmann, P.; Tschan, H.; Baron, R.; Wonisch, M.; Renezeder, K. Menstrual cycle: No effect on exercise cardiorespiratory variables or blood lactate concentration. Med. Sci. Sports Exerc. 2007, 39, 1098–1106. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Mean ± SD |
|---|---|
| Age (yrs) | 20.1 ± 0.5 |
| Height (cm) | 169.2 ± 6.3 |
| BM (kg) | 67.6 ± 9.1 |
| BMI (kg·m2) | 23.8 ± 7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnes, M.E.; Cowan, C.R.; Boag, L.E.; Hill, J.G.; Jones, M.L.; Nixon, K.M.; Parker, M.G.; Parker, S.K.; Raymond, M.V.; Sternenberg, L.H.; et al. Effects of Acute Yohimbine Hydrochloride Supplementation on Repeated Supramaximal Sprint Performance. Int. J. Environ. Res. Public Health 2022, 19, 1316. https://doi.org/10.3390/ijerph19031316
Barnes ME, Cowan CR, Boag LE, Hill JG, Jones ML, Nixon KM, Parker MG, Parker SK, Raymond MV, Sternenberg LH, et al. Effects of Acute Yohimbine Hydrochloride Supplementation on Repeated Supramaximal Sprint Performance. International Journal of Environmental Research and Public Health. 2022; 19(3):1316. https://doi.org/10.3390/ijerph19031316
Chicago/Turabian StyleBarnes, Megan E., Camryn R. Cowan, Lauren E. Boag, Julianne G. Hill, Morgan L. Jones, Kylie M. Nixon, Mckenzie G. Parker, Shelby K. Parker, Mary V. Raymond, Lillie H. Sternenberg, and et al. 2022. "Effects of Acute Yohimbine Hydrochloride Supplementation on Repeated Supramaximal Sprint Performance" International Journal of Environmental Research and Public Health 19, no. 3: 1316. https://doi.org/10.3390/ijerph19031316
APA StyleBarnes, M. E., Cowan, C. R., Boag, L. E., Hill, J. G., Jones, M. L., Nixon, K. M., Parker, M. G., Parker, S. K., Raymond, M. V., Sternenberg, L. H., Tidwell, S. L., Yount, T. M., Williams, T. D., Rogers, R. R., & Ballmann, C. G. (2022). Effects of Acute Yohimbine Hydrochloride Supplementation on Repeated Supramaximal Sprint Performance. International Journal of Environmental Research and Public Health, 19(3), 1316. https://doi.org/10.3390/ijerph19031316