
Sulfidic Habitats in the Gypsum Karst System of Monte Conca (Italy) Host a Chemoautotrophically Supported Invertebrate Community

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
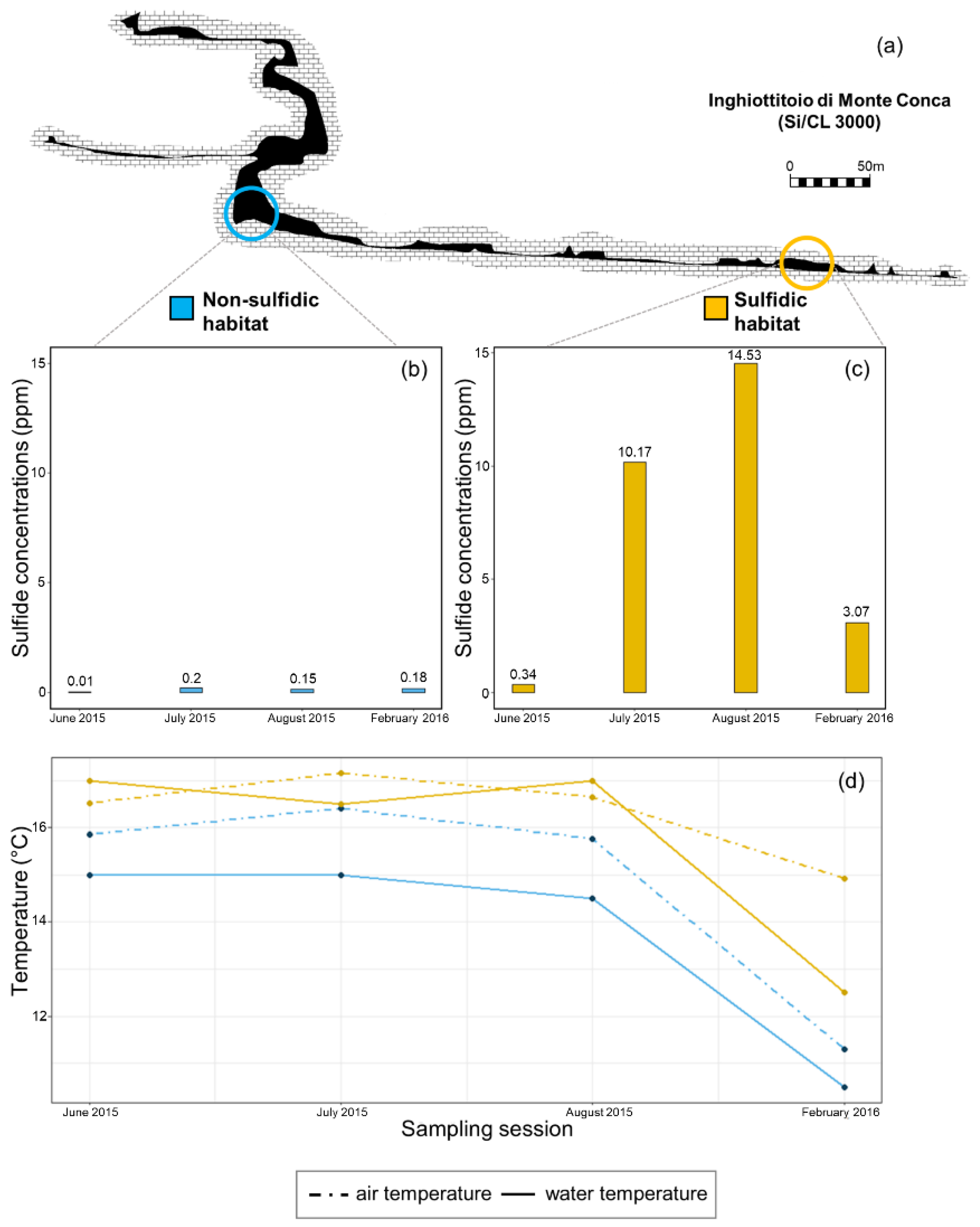
2.1. Site Description
2.2. Habitat Characterization
2.3. Biological Survey
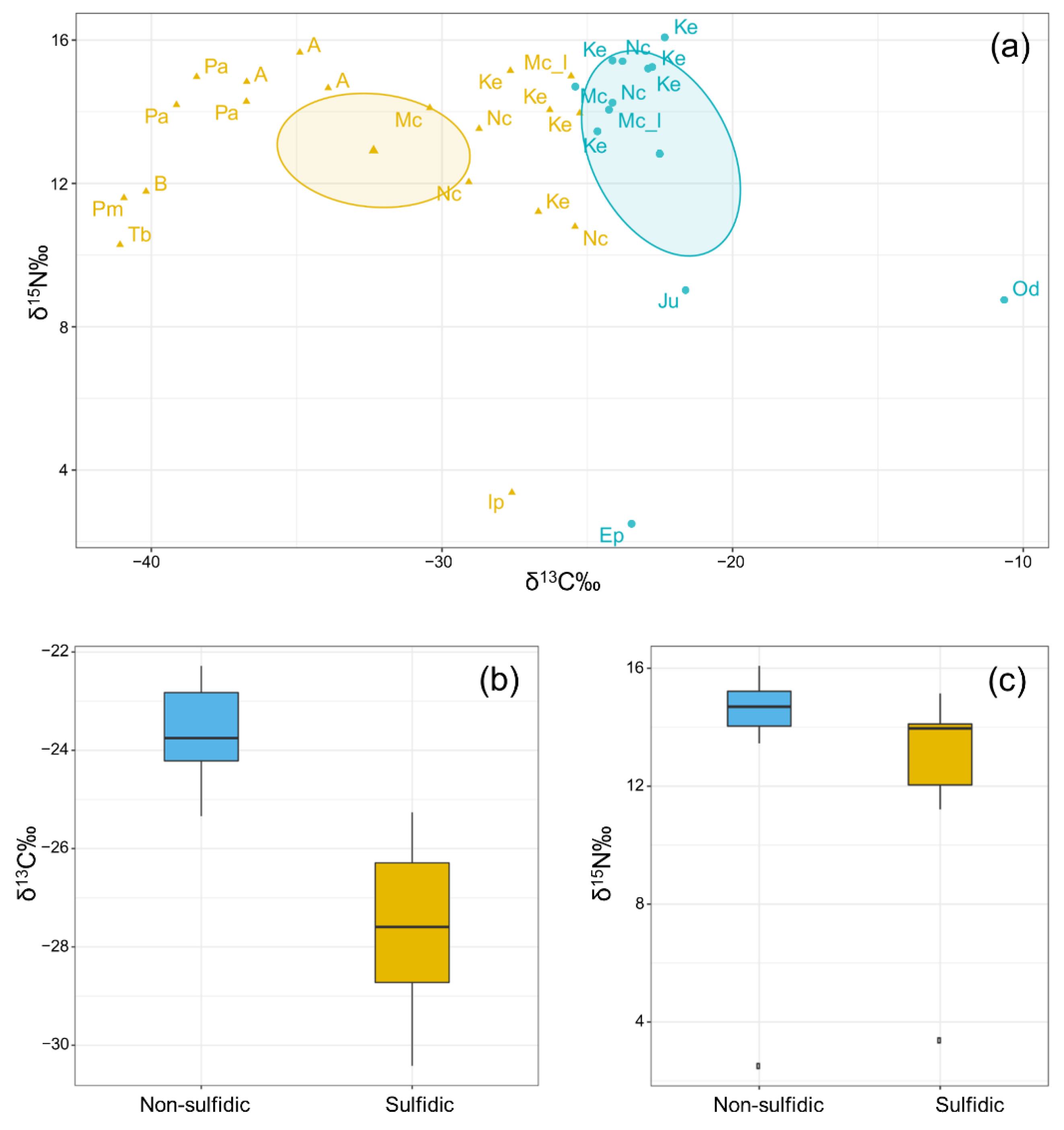
2.4. Stable Isotope Analysis
3. Results
3.1. Habitat Characterization
3.2. Biological Survey
3.3. Stable Isotope Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Culver, D.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2019; 336p. [Google Scholar] [CrossRef] [Green Version]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Simon, K.S.; Benfield, E.F.; Macko, S.A. Food web structure and the role of epilithic biofilms in cave streams. Ecology 2003, 84, 2395–2406. [Google Scholar] [CrossRef] [Green Version]
- Hüppop, K. Adaptation to low food. In Encyclopedia of Cave; Culver, D.C., White, B., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 4–10. [Google Scholar]
- Fenolio, D.B.; Graening, G.O.; Collier, B.A.; Stout, J.F. Coprophagy in a cave–Adapted salamander; the importance of bat guano examined through nutritional and stable isotope analyses. Proc. R. Soc. B 2006, 273, 439–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fišer, Z. Adaptation to low food. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Elsevier Academic Press: London, UK, 2019; pp. 1–7. [Google Scholar] [CrossRef]
- Sarbu, S.M.; Kane, T.C.; Kinkle, B.K. A chemoautotrophically based cave ecosystem. Science 1996, 272, 1953–1955. [Google Scholar] [CrossRef]
- Stevens, T. Lithoautotrophy in the subsurface. FEMS Microbiol. Rev. 1997, 20, 327–337. [Google Scholar] [CrossRef]
- Galdenzi, S.; Sarbu, S.M. Chemiosintesi e speleogenesi in un ecosistema ipogeo: I Rami Sulfurei delle Grotte di Frasassi (Italia centrale). Le Grotte D’italia 2000, 1, 3–18. [Google Scholar]
- Krumholz, L.R. Microbial communities in the deep subsurface. Hydrogeol. J. 2000, 8, 4–10. [Google Scholar]
- Por, F.D.; Dimentman, C.; Frumkin, A.; Naaman, I. Animal life in the chemoautotrophic ecosystem of the hypogenic groundwater cave of Ayyalon (Israel): A summing up. Nat. Sci. 2013, 5, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Popa, I.; Brad, T.; Vaxevanopoulos, M.; Giurginca, A.; Baba, S.; Iepure, S.; Plaiasu, R.; Sarbu, S.M. Rich and diverse subterranean invertebrate communities inhabiting Melissoreypa cave in central Greece. Trav. Inst. Spéol. 2019, 58, 65–78. [Google Scholar]
- Brad, T.; Iepure, S.; Sarbu, S.M. The Chemoautotrophically Based Movile Cave Groundwater Ecosystem, a Hotspot of Subterranean Biodiversity. Diversity 2021, 13, 128. [Google Scholar] [CrossRef]
- Hose, L.D.; Pisarowicz, J.A. Cueva de Villa Luz, Tabasco, Mexico: Reconnaissance Study of an Active Sulfur Spring Cave and Ecosystem. J. Cave Karst Stud. 1999, 61, 13–21. [Google Scholar]
- Sarbu, S.M. Movile Cave: A chemoautotrophically based groundwater ecosystem. In Ecosystems of the World: Subterranean Ecosystems; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Elsevier: New York, NY, USA, 2001; pp. 319–344. [Google Scholar]
- Engel, A.S. Observations on the biodiversity of sulfidic karst habitats. J. Cave Karst Stud. 2007, 69, 187–206. [Google Scholar]
- Porter, M.L.; Engel, A.S.; Kane, T.C.; Kinkle, B.K. Productivity-diversity relationships from chemolithoautotrophically based sulfidic karst systems. Int. J. Speleol. 2009, 38, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Sarbu, S.M. The unusual fauna of a cave with thermomineral waters containing H2S from Southern Dobrogea, Romania. Mem. Biospeolog. 1990, 17, 191–195. [Google Scholar]
- Vlasceanu, L.; Popa, R.; Kinkle, B.K. Characterization of Thiobacillus thioparus LV43 and its distribution in a chemoautotrophically based groundwater ecosystem. Appl. Environ. Microbiol. 1997, 63, 3123–3127. [Google Scholar] [CrossRef] [Green Version]
- Lascu, C. Paleogeographical and hydrogeological hypothesis regarding the origin of a peculiar cave fauna. Misc. Speol. Rom Bucharest. 1989, 1, 13–18. [Google Scholar]
- Sarbu, S.M.; Popa, R. A unique chemoautotrophically based cave ecosystem. In The Natural History of Biospeleology; Camacho, A.I., Ed.; Editorial CSIC—CSIC Press: Madrid, Spain, 1992; pp. 637–666. [Google Scholar]
- Engel, A.S.; Porter, M.L.; Kinkle, B.K.; Kane, T.C. Ecological assessment and geological significance of microbial communities from Cesspool Cave, Virginia. Geomicrobiol. J. 2001, 18, 259–274. [Google Scholar] [CrossRef]
- Madonia, G.; Vattano, M. New knowledge on the Monte Conca gypsum karst system (central-western Sicily, Italy). Acta Carsol. 2011, 40, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Messina, M.; Grech, T.; Fiorenza, F.; Marletta, A.; Valenti, P.; Petralia, S. Sulfidic spring in the gypsum karst system of Monte Conca (Italy): Chemistry and microbiological evidences. Int. J. Speleol. 2015, 44, 2. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.C.; Messina, M.A.; Nicolosi, G.; Petralia, S.; Baker, M.D.; Mayne, C.K.S.; Dinon, C.M.; Moss, C.J.; Onac, B.P.; Garey, J.R. Surface runoff alters cave microbial community structure and function. PLoS ONE 2020, 15, e0232742. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters 1. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- GBIF.org. Available online: https://www.gbif.org (accessed on 10 July 2021).
- De Niro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Vlasceanu, L.; Sarbu, S.M.; Engel, A.S.; Kinkle, B.K. Acidic cave-wall biofilms located in the Frasassi Gorge, Italy. Geomicrobiol. J. 2000, 17, 125–139. [Google Scholar] [CrossRef]
- Engel, A.S.; Porter, M.L.; Stern, L.A.; Quinlan, S.; Bennett, P.C. Bacterial diversity and ecosystem function of filamentous microbial mats from aphotic (cave) sulfidic springs dominated by chemolithoautotrophic “Epsilonproteobacteria”. FEMS Microbiol. Ecol. 2004, 51, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, M.G.; Beggio, M.; Dreon, A.L.; Pamio, A.; Gomiero, T.; Brilli, T.; Dorigo, T.; Concheri, G.; Squartini, A.; Summers Engel, A. A New foodweb based on microbes in calcitic caves: The Cansiliella (Beetles) case in Northern Italy. Int. J. Speleol. 2011, 40, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Michener, R.; Lajtha, K. Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Wiley-Blackwell: New York, NY, USA, 2007. [Google Scholar]
- R Core Team and Contributors Worldwide. Stats: The R Stats Package Version 4.2.0. Available online: https://stat.ethz.ch/R-manual/R-devel/library/stats/html/00Index.html (accessed on 20 November 2021).
- Rodriguez, P.; Martinez-Madrid, M.; Arrate, J.A.; Navarro, E. Selective feeding by the aquatic oligochaete Tubifex tubifex (Tubificidae, Clitellata). Hydrobiologia 2001, 463, 133–140. [Google Scholar] [CrossRef]
- Sarbu, S.M.; Galdenzi, S.; Menichetti, M.; Gentile, G. Geology and biology of Grotte di Frasassi (Frasassi Caves) in Central Italy, an ecological multi-disciplinary study of a hypogenic underground karst system. In Ecosystems of the World: Subterranean Ecosystems; Wilkens, H., Culver, D.C., Humphreys, S., Eds.; Elsevier Science: Oxford, UK, 2000; Volume 30, pp. 361–381. [Google Scholar]
- Volpers, M.; Neumann, D. Tolerance of two tubificid species (Tubifex tubifex and Limnodrilus hoffmeisteri) to hypoxic and sulfidic conditions in novel, long-term experiments. Arch. Für Orientforsch. 2005, 164, 13–38. [Google Scholar] [CrossRef]
- Martins, R.T.; Stephan, N.N.C.; Alves, R.G. Tubificidae (Annelida: Oligochaeta) as an indicator of water quality in an urban stream in southeast Brazil. Acta Limnol. Bras. 2008, 20, 221–226. [Google Scholar]
- Idroos, F.S.; Manage, P.M. Aquatic life health quality assessment of the Bolgoda Canal and Waga Stream with respect to selected physico-chemical parameters and bioindicators. J. Trop. For. Environ. 2012, 2, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Issel, R. Saggio sulla fauna termale italiana. Nota I. Turin R. Accad. D. Sci. 1901, 36, 53–72. [Google Scholar]
- Issel, R. Saggio sulla fauna termale italiana. Nota II. Turin R. Accad. D. Sci. 1901, 36, 3–15, [reprint]. [Google Scholar]
- Nardi, G. Note preliminari sull’entomofauna acquatica dei Monti Lepini (Lazio) (Ephemeroptera, Odonata, Plecoptera, Heteroptera, Coleoptera, Diptera, Tricoptera). In Ricerche Sulla Fauna Dei Monti Lepini. Quaderni Del Museo Storia Naturale Di Patrica; Corsetti, L., Nardi, G., Eds.; Comune di Patrica: Patrica, Italy, 1994; Volume 4, 184p. [Google Scholar]
- Latella, L.; Di Russo, C.; De Pasquale, L.; Dell’Anna, L.; Nardi, G.; Rampini, M. Preliminary investigations on a new sulfurous cave in central Italy. Mem. Biospeolog. 1999, 26, 131–135. [Google Scholar]
- Brignoli, P.M. Ragni d’Italia XXXII. Specie cavernicole di Sicilia (Araneae). Animalia 1979, 5, 273–286. [Google Scholar]
- Simon, E. Araneae et Opiliones (Seconde Série). Biospeologica. XV. Arch. Zool. Exp. Gen. 1910, 5, 49–66. [Google Scholar]
- Fage, L. Araneae, 5e série, précédée d’un essai sur l’évolution souterraine et son déterminisme. Biospeologica. XV. Arch. Zool. Exp. Gen. 1931, 71, 91–291. [Google Scholar]
- Denis, J. Notes d’aranéologie marocaine. X. Les érigonides du Maroc. Bull. Soc. Sc. Nat. Maroc. 1968, 47, 137–164. [Google Scholar]
- Thompson, D.B.; Olson, R.A. A preliminary survey of the protozoa and bacteria from Sulphur River, in Parker’s Cave, Kentucky. Natl. Speleol. Soc. Bull. 1988, 50, 42–46. [Google Scholar]
- Pohlman, J.W.; Iliffe, T.M.; Cifuentes, L.A. A stable isotope study of organic cycling and the ecology of an anchialine cave ecosystem. Mar. Ecol. Prog. Ser. 1997, 155, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Van Dover, C.L. Trophic relationships among invertebrates at the Kairei hydrothermal vent field (Central Indian Ridge). Mar. Biol. 2002, 141, 761–772. [Google Scholar] [CrossRef]
- MacAvoy, S.E.; Morgan, E.; Carney, R.S.; Macko, S.A. Chemoautotrophic production incorporated by heterotrophs in Gulf of Mexico hydrocarbon seeps: An examination of mobile benthic predators and seep residents. J. Shellfish Res. 2008, 27, 153–161. [Google Scholar] [CrossRef]
- Galdenzi, S.; Cocchioni, M.; Morichetti, L.; Amici, V.; Scuri, S. Sulfidic ground-water chemistry in the Frasassi caves, Italy. J. Cave Karst Stud. 2008, 70, 94–107. [Google Scholar]
- Roach, K.A.; Tobler, M.; Winemiller, K.O. Hydrogen sulfide, bacteria, and fish: A unique, subterranean food chain. Ecology 2011, 92, 2056–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.Z.; De la Rosa, J.M.; Jiménez-Morillo, N.T.; Pereira, M.F.C.; González-Pérez, J.A.; Calaforra, J.M.; Saiz-Jimenez, C. Analytical pyrolysis and stable isotope analyses reveal past environmental changes in coralloid speleothems from Easter Island (Chile). J. Chromatogr. A 2016, 1461, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.Z.; Garcia-Sanchez, A.M.; Martin-Sanchez, P.M.; Costa Pereira, M.F.; Spangenberg, J.E.; Jurado, V.; Dionísio, A.; Afonso, M.J.; Iglésias Chaminé, H.I.; Hermosin, B.; et al. Origin of abundant moonmilk deposits in a sub-surface granitic environment. Sedimentology 2018, 65, 1482–1503. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.Z.; De la Rosa, J.M.; Jiménez-Morillo, N.T.; Pereira, M.F.C.; Gonzalez-Perez, J.A.; Knicker, H.; Saiz-Jimenez, C. Impact of wildfires on subsurface volcanic environments: New insights into speleothem chemistry. Sci. Total Environ. 2020, 698, 134321. [Google Scholar] [CrossRef]
- Gonzalez-Pimentel, J.L.; Miller, A.Z.; Jurado, V.; Laiz, L.; Pereira, M.F.; Saiz-Jimenez, C. Yellow coloured mats from lava tubes of La Palma (Canary Islands, Spain) are dominated by metabolically active Actinobacteria. Sci. Rep. 2018, 8, 1944. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Season | Rainfall (mm) of the Previous Month | Number of Rainy Days in the Previous Month | Daily Mean Outside Temperature |
|---|---|---|---|---|
| 20 June 2015 | wet | 44.2 | 8 | 20.28 |
| 11 July 2015 | dry | 0 | 0 | 26.16 |
| 29 August 2015 | dry | 18 | 5 | 25.5 |
| 6 February 2016 | wet | 117 | 21 | 7.55 |
| Cave Sector | Predators | Detritivores | Others | Total | ||||
|---|---|---|---|---|---|---|---|---|
| W | D | W | D | W | D | W | D | |
| Entrance | 18 | 9 | 3 | 3 | 6 | 4 | 27 | 16 |
| Non-sulfidic | 16 | 7 | 16 | 7 | 7 | 5 | 39 | 19 |
| Sulfidic | 20 | 9 | 20 | 14 | 8 | 6 | 48 | 29 |
| Total | 24 | 20 | 20 | 18 | 9 | 8 | 54 | 43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolosi, G.; Galdenzi, S.; Messina, M.A.; Miller, A.Z.; Petralia, S.; Sarbu, S.M.; Isaia, M. Sulfidic Habitats in the Gypsum Karst System of Monte Conca (Italy) Host a Chemoautotrophically Supported Invertebrate Community. Int. J. Environ. Res. Public Health 2022, 19, 2671. https://doi.org/10.3390/ijerph19052671
Nicolosi G, Galdenzi S, Messina MA, Miller AZ, Petralia S, Sarbu SM, Isaia M. Sulfidic Habitats in the Gypsum Karst System of Monte Conca (Italy) Host a Chemoautotrophically Supported Invertebrate Community. International Journal of Environmental Research and Public Health. 2022; 19(5):2671. https://doi.org/10.3390/ijerph19052671
Chicago/Turabian StyleNicolosi, Giuseppe, Sandro Galdenzi, Maria Anna Messina, Ana Z. Miller, Salvatore Petralia, Serban M. Sarbu, and Marco Isaia. 2022. "Sulfidic Habitats in the Gypsum Karst System of Monte Conca (Italy) Host a Chemoautotrophically Supported Invertebrate Community" International Journal of Environmental Research and Public Health 19, no. 5: 2671. https://doi.org/10.3390/ijerph19052671
APA StyleNicolosi, G., Galdenzi, S., Messina, M. A., Miller, A. Z., Petralia, S., Sarbu, S. M., & Isaia, M. (2022). Sulfidic Habitats in the Gypsum Karst System of Monte Conca (Italy) Host a Chemoautotrophically Supported Invertebrate Community. International Journal of Environmental Research and Public Health, 19(5), 2671. https://doi.org/10.3390/ijerph19052671