
Comparative Nanopore Sequencing-Based Evaluation of the Midgut Microbiota of the Summer Chafer (Amphimallon solstitiale L.) Associated with Possible Resistance to Entomopathogenic Nematodes


 , , , ,
, , , , 
Abstract
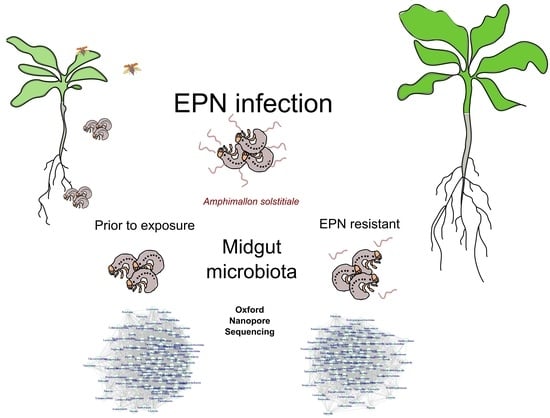
:
1. Introduction
2. Materials and Methods
2.1. Collection and Preparation of Larvae
2.2. DNA Extraction and Nanopore High-Throughput Sequencing
2.3. Bioinformatic Analysis
2.4. Exploratory Data Analyses
3. Results
3.1. Resistance of A. solstitiale Larvae to EPN Infection
3.2. Nanopore Sequencing Results
3.3. Bacterial Community Composition in the Midgut of the T0 Group of A. solstitiale Larvae

3.4. Bacterial Community Composition in the Midgut of the EPN-Resistant Group of A. solstitiale Larvae
3.5. Comparison of the Midgut Microbiota between the T0 and the EPN-Resistant Groups of Insects
4. Discussion
4.1. Natural Midgut Microbiota of A. solstitiale Larvae
4.2. Alterations in the Gut Microbial Composition Associated with EPN-Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOSIM | analysis of similarities |
| BLAST | basic local alignment Search Tool |
| EPN | entomopathogenic nematodes |
| IJs | infective juvenile |
| NCBI | National Center for Biotechnology Information |
| OTU | operative taxonomic unit |
| PCoA | principal component analysis |
| PCR | polymerase chain reaction |
| RA | average relative abundance |
References
- Jackson, T.A.; Klein, M.G. Scarabs as pests: Continuing problem. Coleopt. Bull. 2006, 60, 102–119. [Google Scholar] [CrossRef]
- Harrison Jdu, G.; Wingfield, M.J. A taxonomic review of white grubs and leaf chafers (Coleoptera: Scarabaeidae: Melolonthinae) recorded from forestry and agricultural crops in Sub-Saharan Africa. Bull. Entomol. Res. 2016, 6, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Yan, X.; Zhao, C.; Han, R. Efficacy of entomopathogenic Steinernema and Heterorhabditis nematodes against white grubs (Coleoptera: Scarabaeidae) in peanut fields. J. Econ. Entomol. 2013, 106, 1112–1117. [Google Scholar] [CrossRef]
- Patil, J.; Vijayumar, R.; Nagesh, M.; Holajjer, P. Biocontrol potential of entomopathogenic Heterorhabditis and Steinernema nematodes against Phyllognathus dionysius Fabricius (Coleoptera: Scarabaeidae). Biol. Control 2019, 140, 104103. [Google Scholar] [CrossRef]
- Raja, R.K.; Arun, A.; Touray, M.; Gulsen, S.H.; Cimen, H.; Gulcu, B.; Hazir, C.; Aiswarya, D.; Ulug, D.; Cakmak, I.; et al. Antagonists and defense mechanisms of entomopathogenic nematodes and their mutualistic bacteria. Biol. Control 2020, 152, 104452. [Google Scholar] [CrossRef]
- Allahverdipour, H.; Karimi, J. Nematodes versus white grubs: Long but challenging association. Ann. Entomol. Soc. Am. 2021, 114, 448–458. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Shokal, U.; Yadav, S.; Kenney, E.; Maldonado, T. Insect immunity to entomopathogenic nematodes and their mutualistic bacteria. Curr. Top. Microbiol. Immunol. 2017, 402, 123–156. [Google Scholar] [CrossRef] [PubMed]
- Rolhion, N.; Chassaing, B. When pathogenic bacteria meet the intestinal microbiota. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 71, 20150504. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Perlman, S.J. Toxin-mediated protection against natural enemies by insect defensive symbionts. Adv. Insect Physiol. 2020, 58, 277–303. [Google Scholar] [CrossRef]
- Arora, A.K.; Douglas, A.E. Hype or opportunity? Using microbial symbionts in novel strategies for insect pest control. J. Insect Physiol. 2017, 103, 10–17. [Google Scholar] [CrossRef]
- Qadri, M.; Short, S.; Gast, K.; Hernandez, J.; Wong, A.C.-N. Microbiome innovation in agriculture: Development of microbial based tools for insect pest management. Front. Sustain. Food Syst. 2020, 4, 547751. [Google Scholar] [CrossRef]
- Stock, S.P. Diversity, biology and evolutionary relationships. In Nematode Pathogenesis of Insects and Other Pests: Ecology and Applied Technologies for Sustainable Plant and Crop Protection; Campos-Herrera, R., Ed.; Springer International Publishing: Neuchâtel, Switzerland, 2015; pp. 3–27. [Google Scholar]
- Skowronek, M.; Sajnaga, E.; Kazimierczak, W.; Lis, M.; Wiater, A. Screening and molecular identification of bacteria from the midgut of Amphimallon solstitiale larvae exhibiting antagonistic activity against bacterial symbionts of entomopathogenic nematodes. Int. J. Mol. Sci. 2021, 22, 12005. [Google Scholar] [CrossRef]
- Ziganshina, E.; Mohammed, W.; Shagimardanova, E.; Vankov, P.; Gogoleva, N.; Ziganshin, A. Fungal, bacterial, and archaeal diversity in the digestive tract of several beetle larvae (Coleoptera). Biomed Res. Int. 2018, 2018, 6765438. [Google Scholar] [CrossRef]
- Alonso-Pernas, P.; Arias-Cordero, E.; Novoselov, A.; Ebert, C.; Rybak, J.; Kaltenpoth, M.; Westermann, M.; Neugebauer, U.; Boland, W. Bacterial community and PHB-accumulating bacteria associated with the wall and specialized niches of the hindgut of the forest cockchafer (Melolontha hippocastani). Front. Microbiol. 2017, 28, 291. [Google Scholar] [CrossRef] [Green Version]
- Sajnaga, E.; Skowronek, M.; Kalwasińska, A.; Kazimierczak, W.; Ferenc, K.; Lis, M.; Wiater, A. Nanopore-sequencing characterization of the gut microbiota of Melolontha melolontha larvae: Contribution to protection against entomopathogenic nematodes? Pathogens 2021, 10, 396. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E. Effect of white grub developmental stage on susceptibility to entomopathogenic nematodes. J. Econ. Entomol. 2004, 97, 1842–1849. [Google Scholar] [CrossRef]
- Suzuki, M.T.; Giovannoni, S.J. Bias caused by template annealing in the amplification of mixtures of 16S rRNA genes by PCR. Appl. Environ. Microbiol. 1996, 62, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA 1985, 82, 6955–6959. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electronica 2001, 4, 1–9. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, 1003531. [Google Scholar] [CrossRef] [Green Version]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Constructing and analyzing microbiome networks in R. Methods Mol. Biol. 2018, 1849, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.; Alm, E.J. Inferring correlation networks from genomic survey data. PLoS Comput. Biol. 2012, 8, e1002687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandes, U. A faster algorithm for betweenness centrality. J. Math. Sociol. 2001, 25, 163–177. [Google Scholar] [CrossRef]
- Pons, P.; Latapy, M. Computing communities in large networks using random walks. Lect. Notes Comput. Sci. 2005, 3733, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Thiyonila, B.; Reneeta, N.P.; Kannan, M.; Shantkriti, S.; Krishnan., M. Dung beetle gut microbes: Diversity, metabolic and immunity related roles in host system. Int. J. Sci. Innovs. 2018, 4, 77–83. [Google Scholar] [CrossRef]
- Lemke, T.; Stingl, U.; Egert, M.; Friedrich, M.W.; Brune, A. Physicochemical conditions and microbial activities in the highly alkaline gut of the humus-feeding larva of Pachnoda ephippiata (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 2003, 69, 6650–6658. [Google Scholar] [CrossRef] [Green Version]
- Egert, M.; Stingl, U.; Bruun, L.D.; Brune, A.; Friedrich, M.W.; Pommerenke, B. Structure and topology of microbial communities in the major gut compartments of Melolontha melolontha larvae (Coleoptera:Scarabaeidae). Appl. Environ. Microbiol. 2005, 71, 4556–4566. [Google Scholar] [CrossRef] [Green Version]
- Arias-Cordero, E.; Ping, L.; Reichwald, K.; Delb, H.; Platzer, M.; Boland, W. Comparative evaluation of the gut microbiota associated with the below- and above-ground life stages (larvae and beetles) of the forest cockchafer, Melolontha hippocastani. PLoS ONE 2012, 7, e51557. [Google Scholar] [CrossRef] [Green Version]
- Andert, J.; Marten, A.; Brandl, R.; Brune, A. Inter- and intraspecific comparison of the bacterial assemblages in the hindgut of humivorous scarab beetle larvae (Pachnoda spp.). FEMS Microbiol. Ecol. 2010, 74, 439–449. [Google Scholar] [CrossRef]
- Wang, Y.; Rozen, D.E. Gut microbiota colonization and transmission in the burying beetle Nicrophorus vespilloides throughout development. Appl. Environ. Microbiol. 2017, 83, e03250–e03316. [Google Scholar] [CrossRef] [Green Version]
- Vasanthakumar, A.; Handelsman, J.; Schloss, P.D.; Bauer, L.S.; Raffa, K.F. Gut microbiota of an invasive subcortical beetle, Agrilus planipennis Fairmaire, across various life stages. Environ. Entomol. 2008, 37, 1344–1353. [Google Scholar] [CrossRef]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect Gut Bacterial Diversity Determined by Environmental Habitat, Diet, Developmental Stage, and Phylogeny of Host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zhang, H. The impact of environmental heterogeneity and life stage on the hindgut microbiota of Holotrichia parallela larvae (Coleoptera: Scarabaeidae). PLoS ONE 2013, 8, e57169. [Google Scholar] [CrossRef]
- Franzini, P.Z.; Ramond, J.B.; Scholtz, C.H.; Sole, C.L.; Ronca, S.; Cowan, D.A. The gut microbiomes of two Pachysoma MacLeay desert dung beetle species (Coleoptera: Scarabaeidae: Scarabaeinae) feeding on different diets. PLoS ONE 2016, 11, e0161118. [Google Scholar] [CrossRef]
- Egert, M.; Wagner, B.; Lemke, T.; Brune, A.; Friedrich, M.W. Microbial community structure in midgut and hindgut of the humus-feeding larva of Pachnoda ephippiata (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 2003, 69, 6659–6668. [Google Scholar] [CrossRef] [Green Version]
- Bosmans, L.; Pozo, M.I.; Verreth, C.; Crauwels, S.; Wilberts, L.; Sobhy, S.; Wäckers, F.; Jacquemyn, H.; Lievens, B. Habitat-specific variation in gut microbial communities and pathogen prevalence in bumblebee queens (Bombus terrestris). PLoS ONE 2018, 13, e0204612. [Google Scholar] [CrossRef] [Green Version]
- Park, R.; Dzialo, M.C.; Spaepen, S.; Nsabimana, D.; Gielens, K.; Devriese, H.; Crauwels, S.; Tito, R.Y.; Raes, J.; Lievens, B.; et al. Microbial communities of the house fly Musca domestica vary with geographical location and habitat. Microbiome 2019, 7, 147. [Google Scholar] [CrossRef]
- Fung, T.C.; Vuong, H.E.; Luna, C.D.G.; Pronovost, G.N.; Aleksandrova, A.; Riley, N.G.; Vavilina, A.; McGinn, J.; Rendon, T.; Forrest, L.R.; et al. Intestinal serotonin and fluoxetine exposure modulate bacterial colonization in the gut. Nat. Microbiol. 2019, 4, 2064–2073. [Google Scholar] [CrossRef]
- Hamady, M.; Knight, R. Microbial community profiling for human microbiome projects: Tools, techniques, and challenges. Genome Res. 2009, 19, 1141–1152. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.N.; Chaston, J.; Douglas, A. The inconstant gut microbiota of Drosophila species revealed by 16S rRNA gene analysis. ISME J. 2013, 7, 1922–1932. [Google Scholar] [CrossRef] [Green Version]
- Cambon, M.C.; Lafont, P.; Frayssinet, M.; Lanois, A.; Ogier, J.C.; Pagès, S.; Parthuisot, N.; Ferdy, J.B.; Gaudriault, S. Bacterial community profile after the lethal infection of Steinernema-Xenorhabdus pairs into soil-reared Tenebrio molitor larvae. FEMS Microbiol. Ecol. 2020, 96, fiaa009. [Google Scholar] [CrossRef]
- Skowronek, M.; Sajnaga, E.; Pleszczyńska, M.; Kazimierczak, W.; Lis, M.; Wiater, A. Bacteria from the midgut of common cockchafer (Melolontha melolontha L.) larvae exhibiting antagonistic activity against bacterial symbionts of entomopathogenic nematodes: Isolation and molecular identification. Int. J. Mol. Sci. 2020, 21, 580. [Google Scholar] [CrossRef] [Green Version]
- Topalović, O.; Vestergård, M. Can microorganisms assist the survival and parasitism of plant-parasitic nematodes? Trends Parasitol. 2021, 37, 947–958. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Yadav, S.; Kenney, E.; Cooper, D.; Ozakman, Y.; Patrnogic, J. Role of endosymbionts in insect-parasitic nematode interactions. Trends Parasitol. 2018, 34, 430–444. [Google Scholar] [CrossRef]
- Heise, P.; Liu, Y.; Degenkolb, T.; Vogel, H.; Schäberle, T.F.; Vilcinskas, A. Antibiotic-producing beneficial bacteria in the gut of the burying beetle Nicrophorus vespilloides. Front. Microbiol. 2019, 10, 1178. [Google Scholar] [CrossRef] [Green Version]
- Flórez, L.V.; Scherlach, K.; Miller, I.J.; Rodrigues, A.; Kwan, J.C.; Hertweck, C.; Kaltenpoth, M. An antifungal polyketide associated with horizontally acquired genes supports symbiont-mediated defense in Lagria villosa beetles. Nat. Commun. 2018, 9, 2478. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Grizanova, E.V.; Whitten, M.M.; Mukherjee, K.; Greig, C.; Alikina, T.; Kabilov, M.; Vilcinskas, A.; Glupov, V.V.; Butt, T.M. Immuno-physiological adaptations confer wax moth Galleria mellonella resistance to Bacillus thuringiensis. Virulence 2016, 7, 860–870. [Google Scholar] [CrossRef] [Green Version]
- Tetreau, G.; Grizard, S.; Patil, C.D.; Tran, F.H.; Tran Van, V.; Stalinski, R.; Laporte, F.; Mavingui, P.; Després, L.; Valiente Moro, C. Bacterial microbiota of Aedes aegypti mosquito larvae is altered by intoxication with Bacillus thuringiensis israelensis. Parasit. Vectors 2018, 11, 121. [Google Scholar] [CrossRef] [Green Version]
- Paddock, K.J.; Pereira, A.E.; Finke, D.L.; Ericsson, A.C.; Hibbard, B.E.; Shelby, K.S. Host resistance to Bacillus thuringiensis is linked to altered bacterial community within a specialist insect herbivore. Mol. Ecol. 2021, 30, 1–16. [Google Scholar] [CrossRef]
- Singh, S.; Reese, J.M.; Casanova-Torres, A.M.; Goodrich-Blair, H.; Forst, S. Microbial population dynamics in the hemolymph of Manduca sexta infected with Xenorhabdus nematophila and the entomopathogenic nematode Steinernema carpocapsae. Appl. Environ. Microbiol. 2014, 80, 4277–4285. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Zheng, D.; Zhong, H.; Qin, B.; Gurr, G.M.; Vasseur, L.; Lin, H.; Bai, J.; He, W.; You, M. DNA sequencing reveals the midgut microbiota of diamondback moth, Plutella xylostella (L.) and a possible relationship with insecticide resistance. PLoS ONE 2013, 8, e68852. [Google Scholar] [CrossRef] [PubMed]
- Boissière, A.; Tchioffo, M.T.; Bachar, D.; Abate, L.; Marie, A.; Nsango, S.E.; Shahbazkia, H.R.; Awono-Ambene, P.H.; Levashina, E.; Christen, R.; et al. Midgut Microbiota of the Malaria Mosquito Vector Anopheles gambiae and Interactions with Plasmodium falciparum Infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.; Macchietto, M.; Chang, D.; Barros, M.M.; Baldwin, J.; Mortazavi, A.; Dillman, A.R. Activated entomopathogenic nematode infective juveniles release lethal venom proteins. PLoS Pathog. 2017, 13, e1006302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierking, K.; Yang, W.; Schulenburg, H. Antimicrobial effectors in the nematode Caenorhabditis elegans: An outgroup to the Arthropoda. Philo. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midha, A.; Janek, K.; Niewienda, A.; Henklein, P.; Guenther, S.; Serra, D.O.; Schlosser, J.; Hengge, R.; Hartmann, S. The Intestinal roundworm Ascaris suum releases antimicrobial factors which interfere with bacterial growth and biofilm formation. Front. Cell Infect. Microbiol. 2018, 8, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rausch, S.; Midha, A.; Kuhring, M.; Affinass, N.; Radonic, A.; Kühl, A.A.; Bleich, A.; Renard, B.Y.; Hartmann, S. Parasitic nematodes exert antimicrobial activity and benefit from microbiota-driven support for host immune regulation. Front. Immunol. 2018, 9, 2282. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.P.; Waterfield, N.R.; Daborn, P.J.; Dean, P.; Chilver, T.; Au, C.P.; Sharma, S.; Potter, U.; Reynolds, S.E.; French-Constant, R.H. Bacterial infection of a model insect: Photorhabdus luminescens and Manduca sexta. Cell Microbiol. 2002, 4, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, C.N.; DaMatta, R.A.; Samuels, R.I.; Silva, C.P. Effects of entomopathogenic bacterium Photorhabdus temperata infection on the intestinal microbiota of the sugarcane stalk borer Diatraea saccharalis (Lepidoptera: Crambidae). J. Invertebr. Pathol. 2008, 99, 87–91. [Google Scholar] [CrossRef]
- El-Borai, F.E.; Duncan, L.W.; Preston, J.F. Bionomics of a phoretic association between Paenibacillus sp. and the entomopathogenic nematode Steinernema diaprepesi. J. Nematol. 2005, 37, 18–25, Erratum in: J. Nematol. 2008, 40, 55. [Google Scholar]
- Login, F.H.; Balmand, S.; Vallier, A.; Vincent-Monégat, C.; Vigneron, A.; Weiss-Gayet, M.; Rochat, D.; Heddi, A. Antimicrobial peptides keep insect endosymbionts under control. Science 2011, 334, 362–365. [Google Scholar] [CrossRef]
- Dahan, D.; Preston, G.; Sealey, J.; King, K.C. Impacts of a novel defensive symbiosis on the nematode host microbiome. BMC Microbiol. 2020, 20, 159. [Google Scholar] [CrossRef]
- Li, E.; Qin, J.; Feng, H.; Li, J.; Li, X.; Nyamwasa, I.; Cao, Y.; Ruan, W.; Li, K.; Yin, J. Immune-related genes of the larval Holotrichia parallela in response to entomopathogenic nematodes Heterorhabditis beicherriana LF. BMC Genom. 2021, 22, 192. [Google Scholar] [CrossRef]
- Yadav, S.; Daugherty, S.; Shetty, A.C.; Eleftherianos, I. RNAseq analysis of the Drosophila response to the entomopathogenic nematode Steinernema. G3 2017, 7, 1955–1967. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.X.; Zhang, Y.Q.; Freed, S.; Yu, J.; Gao, Y.F.; Wang, S.; Ouyang, L.N.; Ju, W.Y.; Jin, F.L. An anionic defensin from Plutella xylostella with potential activity against Bacillus thuringiensis. Bull. Entomol. Res. 2016, 106, 790–800. [Google Scholar] [CrossRef]
- Tonk, M.; Knorr, E.; Cabezas-Cruz, A.; Valdés, J.J.; Kollewe, C.; Vilcinskas, A. Tribolium castaneum defensins are primarily active against Gram-positive bacteria. J. Invertebr. Pathol. 2015, 132, 208–215. [Google Scholar] [CrossRef]
- Moore, A.J.; Beazley, W.D.; Bibby, M.C.; Devine, D.A. Antimicrobial activity of cecropins. J. Antimicrob Chemother. 1996, 37, 1077–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Nematode Exposure | Sampling Site * | Developmental Stage of Larva | Total Number of Reads | Classification Rate (%) | Diversity Metrics | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genus Level | Species Level | Sobs | Shannon | Simpson | Evenness | |||||
| G1 | No | FN | L2 | 252.226 | 91.8 | 79.1 | 890 | 4.63 | 0.98 | 0.68 |
| G2 | No | FN | L2 | 175.056 | 96.1 | 83.8 | 646 | 3.49 | 0.93 | 0.54 |
| G3 | No | FN | L3 | 191.586 | 93.7 | 86.1 | 967 | 4.38 | 0.97 | 0.64 |
| G3R | No | FN | L3 | 130.810 | 89.1 | 85.7 | 914 | 4.09 | 0.95 | 0.60 |
| G4 | No | FN | L3 | 161.824 | 95.9 | 91.8 | 1157 | 4.56 | 0.97 | 0.65 |
| G5 | No | FN | L2 | 666.345 | 91.0 | 86.8 | 977 | 4.25 | 0.97 | 0.62 |
| G6 | No | FN | L2 | 202.974 | 86.7 | 81.8 | 969 | 3.96 | 0.91 | 0.58 |
| G7 | No | FN | L2 | 193.900 | 89.4 | 83.4 | 862 | 4.22 | 0.95 | 0.62 |
| G8 | No | UL | L3 | 139.289 | 95.1 | 81.5 | 1464 | 5.32 | 0.99 | 0.73 |
| G9 | No | UL | L3 | 83.109 | 93.8 | 87.7 | 1265 | 4.42 | 0.92 | 0.62 |
| G10 | No | UL | L3 | 112.619 | 93.0 | 75.3 | 1378 | 4.82 | 0.98 | 0.67 |
| G11 | No | UL | L3 | 194.582 | 90.7 | 79.1 | 1182 | 4.33 | 0.95 | 0.61 |
| G12 | No | UL | L3 | 97.439 | 93.4 | 83.7 | 1051 | 4.25 | 0.95 | 0.61 |
| GA1 | Yes | FN | L2 | 137.310 | 92.7 | 86.9 | 647 | 3.59 | 0.93 | 0.55 |
| GA2 | Yes | FN | L2 | 107.186 | 92.6 | 81.3 | 987 | 4.60 | 0.98 | 0.67 |
| GA3 | Yes | FN | L2 | 173119 | 97.6 | 93.2 | 622 | 2.85 | 0.79 | 0.44 |
| GB1 | Yes | FN | L2 | 146.971 | 93.1 | 78.2 | 1360 | 5.01 | 0.98 | 0.69 |
| GB2 | Yes | FN | L2 | 181.119 | 93.3 | 86.7 | 931 | 3.93 | 0.95 | 0.58 |
| GB2R | Yes | FN | L2 | 199.582 | 96.0 | 77.0 | 225 | 0.83 | 0.28 | 0.15 |
| GB3 | Yes | FN | L2 | 217.014 | 99.7 | 69.3 | 926 | 3.26 | 0.87 | 0.48 |
| GC1 | Yes | FN | L2 | 216.091 | 93.2 | 78.1 | 1161 | 4.89 | 0.98 | 0.69 |
| GC2 | Yes | FN | L2 | 175.533 | 93.2 | 85.8 | 851 | 4.04 | 0.96 | 0.60 |
| GC3 | Yes | FN | L2 | 217.951 | 92.1 | 81.9 | 1228 | 3.35 | 0.79 | 0.47 |
| GK1 | Yes | FN | L2 | 158.666 | 95.9 | 74.8 | 521 | 2.37 | 0.76 | 0.38 |
| GK2 | Yes | FN | L2 | 257.766 | 96.3 | 74.6 | 848 | 3.37 | 0.88 | 0.50 |
| GM1 | Yes | FN | L2 | 129.209 | 96.8 | 73.3 | 797 | 3.33 | 0.91 | 0.50 |
| GM2 | Yes | FN | L2 | 300.879 | 95.1 | 81.3 | 898 | 3.71 | 0.94 | 0.55 |
| GM3 | Yes | FN | L2 | 244.162 | 94.7 | 85.0 | 970 | 3.87 | 0.93 | 0.56 |
| Diversity Metrics | T0 | EPN-Resistant | Statistic | Statistic-Value | p-Value |
|---|---|---|---|---|---|
| N = 13 | N = 15 | ||||
| Observed | 1055 | 865 | t | 1.94 | 0.064 |
| Shannon | 4.36 | 3.53 | t | 2.82 | 0.013 |
| Simpson | 8.21 | 6.29 | Mann–Whitney U | 56 | 0.059 |
| Evenness | 8.79 | 5.71 | Mann–Whitney U | 40 | 0.009 |
| T0 Group | EPN-Resistant Group | |
|---|---|---|
| (N = 13) | (N = 15) | |
| Keystone genera | Sphingomonas (1.00 *) | Lachnoclostridium (1.0) |
| Colidextribacter (0.99) | Bosea (0.99) | |
| Cloacibacillus (0.98) | Mesorhizobium (0.98) | |
| Bradyrhizobium (0.96) | Mycolicibacterium (0.96) | |
| Nocardioides (0.95) | Devosia | |
| Number of clusters | 2 | 3 |
| Modularity | 0.067 | 0.126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sajnaga, E.; Skowronek, M.; Kalwasińska, A.; Kazimierczak, W.; Lis, M.; Jach, M.E.; Wiater, A. Comparative Nanopore Sequencing-Based Evaluation of the Midgut Microbiota of the Summer Chafer (Amphimallon solstitiale L.) Associated with Possible Resistance to Entomopathogenic Nematodes. Int. J. Environ. Res. Public Health 2022, 19, 3480. https://doi.org/10.3390/ijerph19063480
Sajnaga E, Skowronek M, Kalwasińska A, Kazimierczak W, Lis M, Jach ME, Wiater A. Comparative Nanopore Sequencing-Based Evaluation of the Midgut Microbiota of the Summer Chafer (Amphimallon solstitiale L.) Associated with Possible Resistance to Entomopathogenic Nematodes. International Journal of Environmental Research and Public Health. 2022; 19(6):3480. https://doi.org/10.3390/ijerph19063480
Chicago/Turabian StyleSajnaga, Ewa, Marcin Skowronek, Agnieszka Kalwasińska, Waldemar Kazimierczak, Magdalena Lis, Monika Elżbieta Jach, and Adrian Wiater. 2022. "Comparative Nanopore Sequencing-Based Evaluation of the Midgut Microbiota of the Summer Chafer (Amphimallon solstitiale L.) Associated with Possible Resistance to Entomopathogenic Nematodes" International Journal of Environmental Research and Public Health 19, no. 6: 3480. https://doi.org/10.3390/ijerph19063480
APA StyleSajnaga, E., Skowronek, M., Kalwasińska, A., Kazimierczak, W., Lis, M., Jach, M. E., & Wiater, A. (2022). Comparative Nanopore Sequencing-Based Evaluation of the Midgut Microbiota of the Summer Chafer (Amphimallon solstitiale L.) Associated with Possible Resistance to Entomopathogenic Nematodes. International Journal of Environmental Research and Public Health, 19(6), 3480. https://doi.org/10.3390/ijerph19063480