
Culture Isolate of Rickettsia felis from a Tick


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Ticks
2.2. Isolation of Rickettsia from Ticks
2.3. Cultivation and Purification of Isolated R. felis
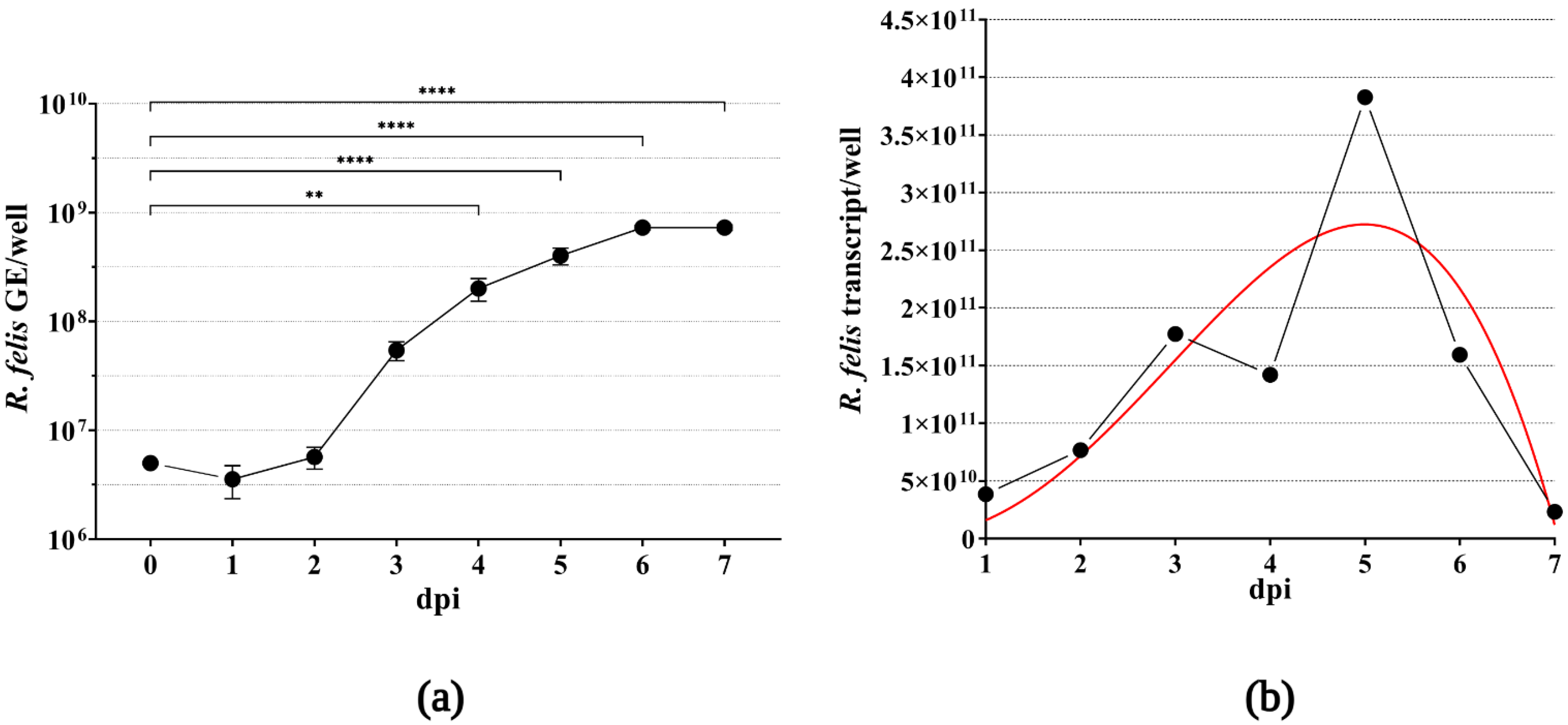
2.4. Monitoring of Rickettsial Intracellular Growth
2.5. RNA Extraction and Reverse-Transcriptase Quantitative PCR (RT-qPCR)
2.6. DNA Extraction
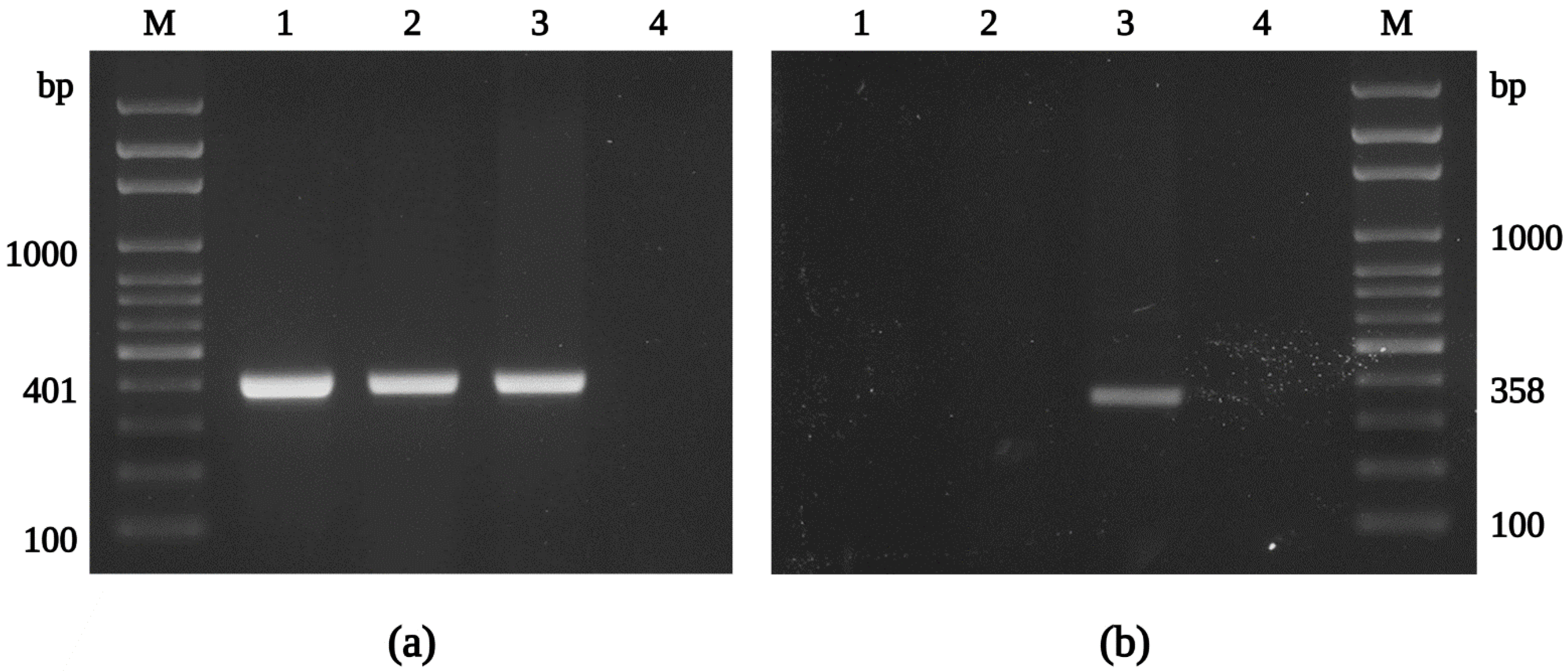
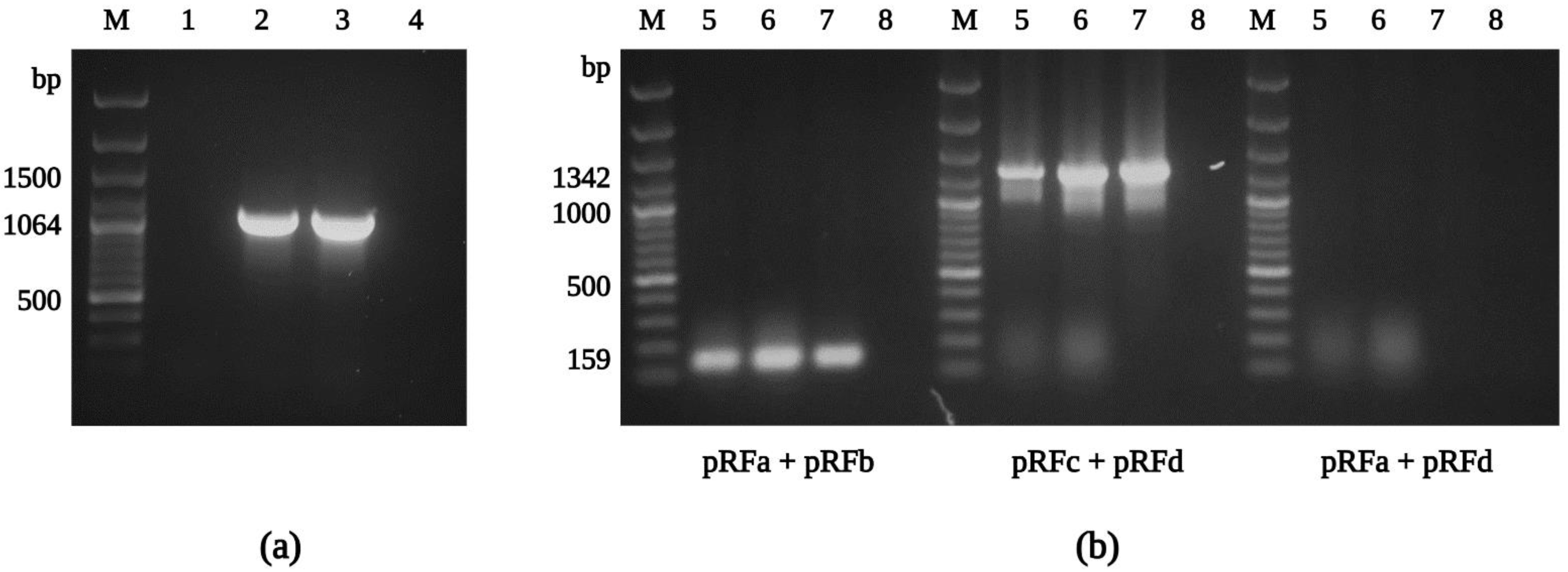
2.7. Molecular Identification by PCR and Sequencing
2.8. Immunofluorescence Assay (IFA)
2.9. Electron Microscopy
2.10. Statistical Analysis
3. Results and Discussion
3.1. Initial Isolation of R. felis from an I. ricinus Nymph
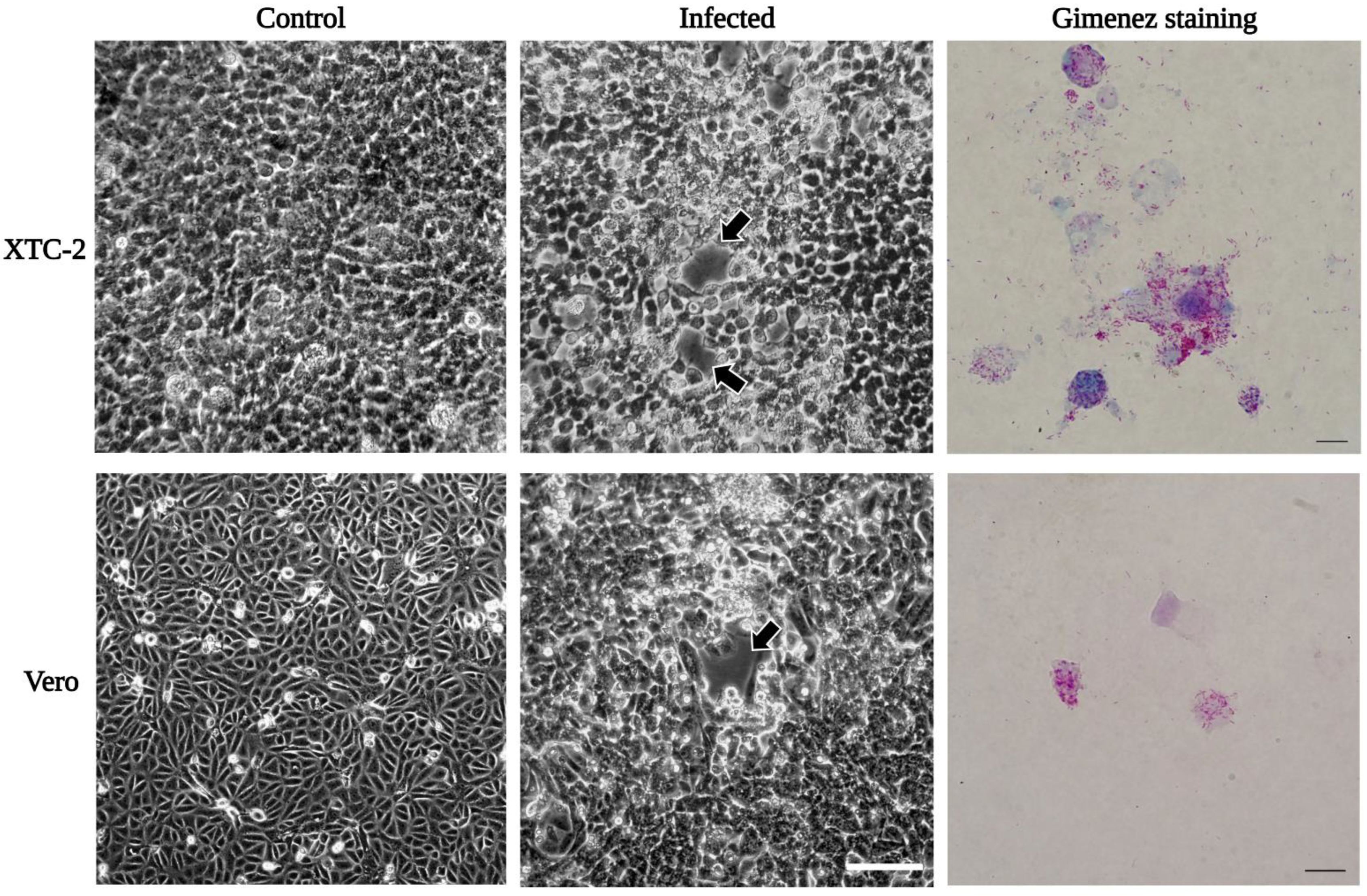
3.2. Propagation of Rickettsiae in XTC-2 and Vero Cell Lines
3.3. Molecular Characterization of the R. felis Isolate Danube
3.4. Microscopy
3.5. A Possible Role of Ticks in the Ecology of R. felis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schriefer, M.E.; Sacci, J.B.; Dumler, J.S.; Bullen, M.G.; Azad, A.F. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J. Clin. Microbiol. 1994, 32, 949–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdad, M.Y.; Stenos, J.; Graves, S. Rickettsia felis, an emerging flea-transmitted human pathogen. Emerg. Health Threat. J. 2011, 4, 7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanton, L.S.; Walker, D.H. Flea-Borne Rickettsioses and Rickettsiae. Am. J. Trop. Med. Hyg. 2017, 96, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Azad, A.F.; Radulovic, S.; Higgins, J.A.; Noden, B.H.; Troyer, J.M. Flea-borne rickettsioses: Ecologic considerations. Emerg. Infect. Dis. 1997, 3, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Zavala-Velázquez, J.E.; Ruiz-Sosa, J.A.; Sánchez-Elias, R.A.; Becerra-Carmona, G.; Walker, D.H. Rickettsia felis rickettsiosis in Yucatán. Lancet 2000, 356, 1079–1080. [Google Scholar] [CrossRef]
- Lindblom, A.; Severinson, K.; Nilsson, K. Rickettsia felis infection in Sweden: Report of two cases with subacute meningitis and review of the literature. Scand. J. Infect. Dis. 2010, 42, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Mawuntu, A.H.P.; Johar, E.; Anggraeni, R.; Feliana, F.; Bernadus, J.B.B.; Safari, D.; Yudhaputri, F.A.; Dhenni, R.; Dewi, Y.P.; Kato, C.; et al. Rickettsia felis identified in two fatal cases of acute meningoencephalitis. PLoS Negl. Trop. Dis. 2020, 14, e0007893. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Osorio, C.E.; Zavala-Velázquez, J.E.; Arias León, J.J.; Zavala-Castro, J.E. Rickettsia felis as emergent global threat for humans. Emerg. Infect. Dis. 2008, 14, 1019–1023. [Google Scholar] [CrossRef]
- Angelakis, E.; Mediannikov, O.; Parola, P.; Raoult, D. Rickettsia felis: The Complex Journey of an Emergent Human Pathogen. Trends Parasitol. 2016, 32, 554–564. [Google Scholar] [CrossRef]
- Parola, P. Rickettsia felis: From a rare disease in the USA to a common cause of fever in sub-Saharan Africa. Clin. Microbiol. Infect. 2011, 17, 996–1000. [Google Scholar] [CrossRef] [Green Version]
- Márquez, F.J.; Muniain, M.A.; Pérez, J.M.; Pachón, J. Presence of Rickettsia felis in the cat flea from southwestern Europe. Emerg. Infect. Dis. 2002, 8, 89–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reif, K.E.; Macaluso, K.R. Ecology of Rickettsia felis: A review. J. Med. Entomol. 2009, 46, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.D.; Macaluso, K.R. Rickettsia felis, an Emerging Flea-Borne Rickettsiosis. Curr. Trop. Med. Rep. 2016, 3, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, L.D.; Freitas, R.N.; Mafra, C.L.; Neves, C.V.; Figueira, F.C.; Labruna, M.B.; Gennari, S.M.; Walker, D.H.; Galvão, M.A. Characterization of Rickettsia spp. circulating in a silent peri-urban focus for Brazilian spotted fever in Caratinga, Minas Gerais, Brazil. Cad. Saude Publica 2006, 22, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, K.A.; Oliveira, L.S.; Dias, C.C.; Silva, A.; Almeida, M.R.; Almada, G.; Bouyer, D.H.; Galvão, M.A.; Mafra, C. Molecular identification of Rickettsia felis in ticks and fleas from an endemic area for Brazilian Spotted Fever. Mem. Inst. Oswaldo Cruz 2008, 103, 191–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, A.; Olmeda, A.S.; Escudero, R.; Jado, I.; Valcárcel, F.; Casado-Nistal, M.A.; Rodríguez-Vargas, M.; Gil, H.; Anda, P. Tick-borne zoonotic bacteria in ticks collected from central Spain. Am. J. Trop. Med. Hyg. 2009, 81, 67–74. [Google Scholar] [CrossRef]
- Abarca, K.; López, J.; Acosta-Jamett, G.; Martínez-Valdebenito, C. Rickettsia felis in Rhipicephalus sanguineus from two distant Chilean cities. Vector Borne Zoonotic Dis. 2013, 13, 607–609. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, G.; Kelly, P.; Zhang, Z.; Wei, L.; Yu, D.; Kayizha, S.; Wang, C. First report of Rickettsia felis in China. BMC Infect. Dis. 2014, 14, 682. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.L.; Colella, V.; Greco, G.; Fang, F.; Nurcahyo, W.; Hadi, U.K.; Venturina, V.; Tong, K.B.Y.; Tsai, Y.L.; Taweethavonsawat, P.; et al. Molecular detection of pathogens in ticks and fleas collected from companion dogs and cats in East and Southeast Asia. Parasites Vectors 2020, 13, 420. [Google Scholar] [CrossRef]
- Campos, S.D.E.; Cunha, N.C.D.; Machado, C.S.C.; Telleria, E.L.; Cordeiro, M.D.; Fonseca, A.H.D.; Toma, H.K.; Santos, J.P.C.D.; Almosny, N.R.P. Rickettsial pathogens circulating in urban districts of Rio de Janeiro, without report of human Brazilian Spotted Fever. Rev. Bras. Parasitol. Vet. 2020, 29, e014220. [Google Scholar] [CrossRef]
- de Oliveira, J.C.P.; Reckziegel, G.H.; Ramos, C.A.D.N.; Giannelli, A.; Alves, L.C.; de Carvalho, G.A.; Ramos, R.A.N. Detection of Rickettsia felis in ectoparasites collected from domestic animals. Exp. Appl. Acarol. 2020, 81, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Gargili, A.; Palomar, A.M.; Midilli, K.; Portillo, A.; Kar, S.; Oteo, J.A. Rickettsia species in ticks removed from humans in Istanbul, Turkey. Vector Borne Zoonotic Dis. 2012, 12, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Raele, D.A.; Galante, D.; Pugliese, N.; Salandra, G.; Cafiero, M.A. Spotted fever group rickettsiae associated with ixodid ticks in wild environment in Southern Italy. Microbiologyopen 2018, 7, e00527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikura, M.; Ando, S.; Shinagawa, Y.; Matsuura, K.; Hasegawa, S.; Nakayama, T.; Fujita, H.; Watanabe, M. Phylogenetic analysis of spotted fever group rickettsiae based on gltA, 17-kDa, and rOmpA genes amplified by nested PCR from ticks in Japan. Microbiol. Immunol. 2003, 47, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Duh, D.; Punda-Polić, V.; Trilar, T.; Petrovec, M.; Bradarić, N.; Avsic-Zupanc, T. Molecular identification of Rickettsia felis-like bacteria in Haemaphysalis sulcata ticks collected from domestic animals in southern Croatia. Ann. N. Y. Acad. Sci. 2006, 1078, 347–351. [Google Scholar] [CrossRef]
- Roth, T.; Lane, R.S.; Foley, J. A Molecular Survey for Francisella tularensis and Rickettsia spp. in Haemaphysalis leporispalustris (Acari: Ixodidae) in Northern California. J. Med. Entomol. 2017, 54, 492–495. [Google Scholar] [CrossRef]
- Khoo, J.J.; Chen, F.; Kho, K.L.; Shanizza, A.I.A.; Lim, F.S.; Tan, K.K.; Chang, L.Y.; AbuBakar, S. Bacterial community in Haemaphysalis ticks of domesticated animals from the Orang Asli communities in Malaysia. Ticks Tick Borne Dis. 2016, 7, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Chalada, M.J.; Stenos, J.; Vincent, G.; Barker, D.; Bradbury, R.S. A Molecular Survey of Tick-Borne Pathogens from Ticks Collected in Central Queensland, Australia. Vector Borne Zoonotic Dis. 2018, 18, 151–163. [Google Scholar] [CrossRef]
- Jiang, J.; Stromdahl, E.Y.; Richards, A.L. Detection of Rickettsia parkeri and Candidatus Rickettsia andeanae in Amblyomma maculatum Gulf Coast ticks collected from humans in the United States. Vector Borne Zoonotic Dis. 2012, 12, 175–182. [Google Scholar] [CrossRef]
- Soares, H.S.; Barbieri, A.R.; Martins, T.F.; Minervino, A.H.; de Lima, J.T.; Marcili, A.; Gennari, S.M.; Labruna, M.B. Ticks and rickettsial infection in the wildlife of two regions of the Brazilian Amazon. Exp. Appl. Acarol. 2015, 65, 125–140. [Google Scholar] [CrossRef]
- Bitencourth, K.; Amorim, M.; DE Oliveira, S.V.; Caetano, R.L.; Voloch, C.M.; Gazêta, G.S. Amblyomma sculptum: Genetic diversity and rickettsias in the Brazilian Cerrado biome. Med. Vet. Entomol. 2017, 31, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Bitencourth, K.; Amorim, M.; de Oliveira, S.V.; Voloch, C.M.; Gazêta, G.S. Genetic diversity, population structure and rickettsias in Amblyomma ovale in areas of epidemiological interest for spotted fever in Brazil. Med. Vet. Entomol. 2019, 33, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Tsui, P.Y.; Tsai, K.H.; Weng, M.H.; Hung, Y.W.; Liu, Y.T.; Hu, K.Y.; Lien, J.C.; Lin, P.R.; Shaio, M.F.; Wang, H.C.; et al. Molecular detection and characterization of spotted fever group rickettsiae in Taiwan. Am. J. Trop. Med. Hyg. 2007, 77, 883–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascucci, I.; Di Domenico, M.; Curini, V.; Cocco, A.; Averaimo, D.; D’Alterio, N.; Cammà, C. Diversity of Rickettsia in Ticks Collected in Abruzzi and Molise Regions (Central Italy). Microorganisms 2019, 7, 696. [Google Scholar] [CrossRef] [Green Version]
- Dobler, G.; Wölfel, R. Typhus and other rickettsioses: Emerging infections in Germany. Dtsch. Arztebl. Int. 2009, 106, 348–354. [Google Scholar] [CrossRef]
- Lejal, E.; Marsot, M.; Chalvet-Monfray, K.; Cosson, J.F.; Moutailler, S.; Vayssier-Taussat, M.; Pollet, T. A three-years assessment of Ixodes ricinus-borne pathogens in a French peri-urban forest. Parasites Vectors 2019, 12, 551. [Google Scholar] [CrossRef] [Green Version]
- Lejal, E.; Moutailler, S.; Šimo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasites Vectors 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Remesar, S.; Díaz, P.; Portillo, A.; Santibáñez, S.; Prieto, A.; Díaz-Cao, J.M.; López, C.M.; Panadero, R.; Fernández, G.; Díez-Baños, P.; et al. Prevalence and molecular characterization of Rickettsia spp. in questing ticks from north-western Spain. Exp. Appl. Acarol. 2019, 79, 267–278. [Google Scholar] [CrossRef]
- Borşan, S.D.; Ionică, A.M.; Galon, C.; Toma-Naic, A.; Peştean, C.; Sándor, A.D.; Moutailler, S.; Mihalca, A.D. High Diversity, Prevalence, and Co-infection Rates of Tick-Borne Pathogens in Ticks and Wildlife Hosts in an Urban Area in Romania. Front. Microbiol. 2021, 12, 645002. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Simin, V.; Mijatović, D.; Obregón, D.; Moutailler, S.; Cabezas-Cruz, A. Humans infested with Ixodes ricinus are exposed to a diverse array of tick-borne pathogens in Serbia. Ticks Tick Borne Dis. 2021, 12, 101609. [Google Scholar] [CrossRef]
- Díaz-Sánchez, A.A.; Chilton, N.B.; Roblejo-Arias, L.; Fonseca-Rodríguez, O.; Marrero-Perera, R.; Diyes, C.P.; Yunik, M.E.M.; Lobo-Rivero, E.; Corona-González, B. Molecular detection and identification of spotted fever group rickettsiae in ticks collected from horses in Cuba. Med. Vet. Entomol. 2021, 35, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Stanley, H.; Rhodes, D.V.L. Presence of Rickettsia Species in Ticks Collected from Companion Animals in Northeastern Georgia, United States. Vet. Sci. 2021, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.K.; Loftis, A.D.; Sanders, F.; Spinks, M.D.; Wills, W.; Denison, A.M.; Dasch, G.A. Borrelia, Coxiella, and Rickettsia in Carios capensis (Acari: Argasidae) from a brown pelican (Pelecanus occidentalis) rookery in South Carolina, USA. Exp. Appl. Acarol. 2006, 39, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Špitalská, E.; Boldiš, V.; Mošanský, L.; Sparagano, O.; Stanko, M. Rickettsia species in fleas collected from small mammals in Slovakia. Parasitol. Res. 2015, 114, 4333–4339. [Google Scholar] [CrossRef] [PubMed]
- Heglasová, I.; Víchová, B.; Stanko, M. Detection of Rickettsia spp. in Fleas Collected from Small Mammals in Slovakia, Central Europe. Vector Borne Zoonotic Dis. 2020, 20, 652–656. [Google Scholar] [CrossRef]
- Adams, J.R.; Schmidtmann, E.T.; Azad, A.F. Infection of colonized cat fleas, Ctenocephalides felis (Bouché), with a rickettsia-like microorganism. Am. J. Trop. Med. Hyg. 1990, 43, 400–409. [Google Scholar] [CrossRef]
- Radulovic, S.; Higgins, J.A.; Jaworski, D.C.; Dasch, G.A.; Azad, A.F. Isolation, cultivation, and partial characterization of the ELB agent associated with cat fleas. Infect. Immun. 1995, 63, 4826–4829. [Google Scholar] [CrossRef] [Green Version]
- Radulovic, S.; Higgins, J.A.; Jaworski, D.C.; Azad, A.F. In vitro and in vivo antibiotic susceptibilities of ELB rickettsiae. Antimicrob. Agents Chemother. 1995, 39, 2564–2566. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.A.; Radulovic, S.; Schriefer, M.E.; Azad, A.F. Rickettsia felis: A new species of pathogenic rickettsia isolated from cat fleas. J. Clin. Microbiol. 1996, 34, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Bouyer, D.H.; Stenos, J.; Crocquet-Valdes, P.; Moron, C.G.; Popov, V.L.; Zavala-Velazquez, J.E.; Foil, L.D.; Stothard, D.R.; Azad, A.F.; Walker, D.H. Rickettsia felis: Molecular characterization of a new member of the spotted fever group. Int. J. Syst. Evol. Microbiol. 2001, 51, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Raoult, D.; La Scola, B.; Enea, M.; Fournier, P.E.; Roux, V.; Fenollar, F.; Galvao, M.A.; de Lamballerie, X. A flea-associated Rickettsia pathogenic for humans. Emerg. Infect. Dis. 2001, 7, 73–81. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Meconi, S.; Fenollar, F.; Rolain, J.M.; Roux, V.; Raoult, D. Emended description of Rickettsia felis (Bouyer et al. 2001), a temperature-dependent cultured bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 2035–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hii, S.F.; Abdad, M.Y.; Kopp, S.R.; Stenos, J.; Rees, R.L.; Traub, R.J. Seroprevalence and risk factors for Rickettsia felis exposure in dogs from Southeast Queensland and the Northern Territory, Australia. Parasites Vectors 2013, 6, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng-Nguyen, D.; Hii, S.F.; Hoang, M.T.; Nguyen, V.T.; Rees, R.; Stenos, J.; Traub, R.J. Domestic dogs are mammalian reservoirs for the emerging zoonosis flea-borne spotted fever, caused by Rickettsia felis. Sci. Rep. 2020, 10, 4151. [Google Scholar] [CrossRef] [PubMed]
- Pornwiroon, W.; Pourciau, S.S.; Foil, L.D.; Macaluso, K.R. Rickettsia felis from cat fleas: Isolation and culture in a tick-derived cell line. Appl. Environ. Microbiol. 2006, 72, 5589–5595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thepparit, C.; Sunyakumthorn, P.; Guillotte, M.L.; Popov, V.L.; Foil, L.D.; Macaluso, K.R. Isolation of a rickettsial pathogen from a non-hematophagous arthropod. PLoS ONE 2011, 6, e16396. [Google Scholar] [CrossRef] [PubMed]
- Horta, M.C.; Labruna, M.B.; Durigon, E.L.; Schumaker, T.T. Isolation of Rickettsia felis in the mosquito cell line C6/36. Appl. Environ. Microbiol. 2006, 72, 1705–1707. [Google Scholar] [CrossRef] [Green Version]
- Hun, L.; Troyo, A.; Taylor, L.; Barbieri, A.M.; Labruna, M.B. First report of the isolation and molecular characterization of Rickettsia amblyommii and Rickettsia felis in Central America. Vector Borne Zoonotic Dis. 2011, 11, 1395–1397. [Google Scholar] [CrossRef]
- Sakamoto, J.M.; Azad, A.F. Propagation of arthropod-borne Rickettsia spp. in two mosquito cell lines. Appl. Environ. Microbiol. 2007, 73, 6637–6643. [Google Scholar] [CrossRef] [Green Version]
- Luce-Fedrow, A.; Macaluso, K.R.; Richards, A.L. Growth of Rickettsia felis in Drosophila melanogaster S2 cells. Vector Borne Zoonotic Dis. 2014, 14, 101–110. [Google Scholar] [CrossRef]
- Saisongkorh, W.; El Karkouri, K.; Patrice, J.Y.; Bernard, A.; Rolain, J.M.; Raoult, D. Tryptose phosphate broth improves Rickettsia felis replication in mammalian cells. FEMS Immunol. Med. Microbiol. 2012, 64, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, F.; Pons, I.; Pla, J.; Nogueras, M.M. Shell-vial culture and real-time PCR applied to Rickettsia typhi and Rickettsia felis detection. World J. Microbiol. Biotechnol. 2015, 31, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, H.L.; Labruna, M.B.; Montenieri, J.A.; Kosoy, M.Y.; Gage, K.L.; Walker, D.H. Detection of Rickettsia felis in a New World flea species, Anomiopsyllus nudata (Siphonaptera: Ctenophthalmidae). J. Med. Entomol. 2005, 42, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Nosek, J.; Sixl, W. Central-European ticks (Ixodoidea). Key for determination. Mitt. Abt. Zool. Landesmus. Joanneum 1972, 1, 61–92. [Google Scholar]
- Sekeyová, Z.; Mediannikov, O.; Subramanian, G.; Kowalczewska, M.; Quevedo-Diaz, M.; Kocianová, E.; Raoult, D. Isolation of rickettsia helvetica from ticks in Slovakia. Acta Virol. 2012, 56, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Pudney, M.; Varma, M.G.; Leake, C.J. Establishment of a cell line (XTC-2) from the South African clawed toad, Xenopus laevis. Experientia 1973, 29, 466–467. [Google Scholar] [CrossRef]
- Giménez, D.F. Staining Rickettsiae in Yolk-Sac Cultures. Stain Technol. 1964, 39, 135–140. [Google Scholar] [CrossRef]
- Ammerman, N.C.; Beier-Sexton, M.; Azad, A.F. Laboratory maintenance of Rickettsia rickettsii. Curr. Protoc. Microbiol. 2008, 11, 3A.5.1–3A.5.21. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.; Coolbaugh, J.C.; Williams, J.C. Separation of viable Rickettsia typhi from yolk sac and L cell host components by renografin density gradient centrifugation. Appl. Microbiol. 1975, 30, 456–463. [Google Scholar] [CrossRef]
- Bohacsova, M.; Filipcik, P.; Opattova, A.; Valarikova, J.; Diaz, M.Q.; Skultety, L.; Novak, M.; Sekeyova, Z. Survival of rat cerebrocortical neurons after rickettsial infection. Microbes Infect. 2015, 17, 845–849. [Google Scholar] [CrossRef]
- Valarikova, J.; Sekeyova, Z.; Skultety, L.; Bohacsova, M.; Quevedo-Diaz, M. New way of purification of pathogenic rickettsiae reducing health risks. Acta Virol. 2016, 60, 206–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of the genus Rickettsia by 16S rDNA sequencing. Res. Microbiol. 1995, 146, 385–396. [Google Scholar] [CrossRef]
- Al Masalma, M.; Armougom, F.; Scheld, W.M.; Dufour, H.; Roche, P.H.; Drancourt, M.; Raoult, D. The expansion of the microbiological spectrum of brain abscesses with use of multiple 16S ribosomal DNA sequencing. Clin. Infect. Dis. 2009, 48, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Sekeyova, Z.; Roux, V.; Raoult, D. Phylogeny of Rickettsia spp. inferred by comparing sequences of ’gene D’, which encodes an intracytoplasmic protein. Int. J. Syst. Evol. Microbiol. 2001, 51, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavala-Castro, J.E.; Small, M.; Keng, C.; Bouyer, D.H.; Zavala-Velázquez, J.; Walker, D.H. Transcription of the Rickettsia felis ompA gene in naturally infected fleas. Am. J. Trop. Med. Hyg. 2005, 73, 662–666. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Drancourt, M.; Raoult, D. Characterization of mutations in the rpoB gene in naturally rifampin-resistant Rickettsia species. Antimicrob. Agents Chemother. 1999, 43, 2400–2403. [Google Scholar] [CrossRef] [Green Version]
- Boretti, F.S.; Perreten, A.; Meli, M.L.; Cattori, V.; Willi, B.; Wengi, N.; Hornok, S.; Honegger, H.; Hegglin, D.; Woelfel, R.; et al. Molecular Investigations of Rickettsia helvetica infection in dogs, foxes, humans, and Ixodes ticks. Appl. Environ. Microbiol. 2009, 75, 3230–3237. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P.E.; Belghazi, L.; Robert, C.; Elkarkouri, K.; Richards, A.L.; Greub, G.; Collyn, F.; Ogawa, M.; Portillo, A.; Oteo, J.A.; et al. Variations of plasmid content in Rickettsia felis. PLoS ONE 2008, 3, e2289. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Danchenko, M.; Csaderova, L.; Fournier, P.E.; Sekeyova, Z. Optimized fixation of actin filaments for improved indirect immunofluorescence staining of rickettsiae. BMC Res. Notes 2019, 12, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobue, K.; Fujio, Y.; Kanda, K. Tumor promoter induces reorganization of actin filaments and calspectin (fodrin or nonerythroid spectrin) in 3T3 cells. Proc. Natl. Acad. Sci. USA 1988, 85, 482–486. [Google Scholar] [CrossRef] [Green Version]
- Benada, O.; Pokorný, V. Modification of the Polaron sputter-coater unit for glow-discharge activation of carbon support films. J. Electron. Microsc. Tech. 1990, 16, 235–239. [Google Scholar] [CrossRef]
- Sekeyová, Z.; Fournier, P.E.; Rehácek, J.; Raoult, D. Characterization of a new spotted fever group rickettsia detected in Ixodes ricinus (Acari: Ixodidae) collected in Slovakia. J. Med. Entomol. 2000, 37, 707–713. [Google Scholar] [CrossRef]
- Boldis, V.; Strus, J.; Kocianová, E.; Tusek-Znidaric, M.; Stefanidesová, K.; Schwarzová, K.; Kúdelová, M.; Sekeyová, Z.; Spitalská, E. Life cycle of Rickettsia slovaca in L929 cell line studied by quantitative real-time PCR and transmission electron microscopy. FEMS Microbiol. Lett. 2009, 293, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Mediannikov, O.; Sekeyová, Z.; Birg, M.L.; Raoult, D. A novel obligate intracellular gamma-proteobacterium associated with ixodid ticks, Diplorickettsia massiliensis, Gen. Nov., sp. Nov. PLoS ONE 2010, 5, e11478. [Google Scholar] [CrossRef]
- Mediannikov, O.; Subramanian, G.; Sekeyova, Z.; Bell-Sakyi, L.; Raoult, D. Isolation of Arsenophonus nasoniae from Ixodes ricinus ticks in Slovakia. Ticks Tick Borne Dis. 2012, 3, 367–370. [Google Scholar] [CrossRef]
- Spitalská, E.; Stefanidesová, K.; Kocianová, E.; Boldiš, V. Rickettsia slovaca and Rickettsia raoultii in Dermacentor marginatus and Dermacentor reticulatus ticks from Slovak Republic. Exp. Appl. Acarol. 2012, 57, 189–197. [Google Scholar] [CrossRef]
- Špitalská, E.; Boldiš, V.; Derdáková, M.; Selyemová, D.; Rusňáková Taragel’ová, V. Rickettsial infection in Ixodes ricinus ticks in urban and natural habitats of Slovakia. Ticks Tick Borne Dis. 2014, 5, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Svehlová, A.; Berthová, L.; Sallay, B.; Boldiš, V.; Sparagano, O.A.; Spitalská, E. Sympatric occurrence of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna ticks and Rickettsia and Babesia species in Slovakia. Ticks Tick Borne Dis. 2014, 5, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Špitalská, E.; Stanko, M.; Mošanský, L.; Kraljik, J.; Miklisová, D.; Mahríková, L.; Bona, M.; Kazimírová, M. Seasonal analysis of Rickettsia species in ticks in an agricultural site of Slovakia. Exp. Appl. Acarol. 2016, 68, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Berthová, L.; Slobodník, V.; Slobodník, R.; Olekšák, M.; Sekeyová, Z.; Svitálková, Z.; Kazimírová, M.; Špitalská, E. The natural infection of birds and ticks feeding on birds with Rickettsia spp. and Coxiella burnetii in Slovakia. Exp. Appl. Acarol. 2016, 68, 299–314. [Google Scholar] [CrossRef]
- Ogata, H.; Renesto, P.; Audic, S.; Robert, C.; Blanc, G.; Fournier, P.E.; Parinello, H.; Claverie, J.M.; Raoult, D. The genome sequence of Rickettsia felis identifies the first putative conjugative plasmid in an obligate intracellular parasite. PLoS Biol. 2005, 3, e248. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Utsuki, T.; Husseneder, C.; Chouljenko, V.N.; Azad, A.F.; Macaluso, K.R. Genomic diversification in strains of Rickettsia felis Isolated from different arthropods. Genome Biol. Evol. 2014, 7, 35–56. [Google Scholar] [CrossRef]
- Bitam, I.; Parola, P.; De La Cruz, K.D.; Matsumoto, K.; Baziz, B.; Rolain, J.M.; Belkaid, M.; Raoult, D. First molecular detection of Rickettsia felis in fleas from Algeria. Am. J. Trop. Med. Hyg. 2006, 74, 532–535. [Google Scholar] [CrossRef]
- Znazen, A.; Rolain, J.M.; Hammami, A.; Jemaa, M.B.; Raoult, D. Rickettsia felis infection, Tunisia. Emerg. Infect. Dis. 2006, 12, 138–140. [Google Scholar] [CrossRef]
- Fang, R.; Raoult, D. Antigenic classification of Rickettsia felis by using monoclonal and polyclonal antibodies. Clin. Diagn. Lab. Immunol. 2003, 10, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Kahl, O.; Lane, R.S.; Levin, M.L.; Tsao, J.I. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick-Borne Dis. 2016, 7, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Matuschka, F.R.; Fischer, P.; Musgrave, K.; Richter, D.; Spielman, A. Hosts on which nymphal Ixodes ricinus most abundantly feed. Am. J. Trop. Med. Hyg. 1991, 44, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Spierling, N.G.; Heuser, E.; Kling, C.; Schmidt, S.; Rosenfeld, U.M.; Reil, D.; Imholt, C.; Jacob, J.; Ulrich, R.G.; et al. High prevalence of Rickettsia helvetica in wild small mammal populations in Germany. Ticks Tick Borne Dis. 2018, 9, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Schex, S.; Dobler, G.; Riehm, J.; Müller, J.; Essbauer, S. Rickettsia spp. in wild small mammals in Lower Bavaria, South-Eastern Germany. Vector Borne Zoonotic Dis. 2011, 11, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.Y.; Chomel, B.B. Hedgehog zoonoses. Emerg. Infect. Dis. 2005, 11, 1–5. [Google Scholar] [CrossRef]
- Jahfari, S.; Ruyts, S.C.; Frazer-Mendelewska, E.; Jaarsma, R.; Verheyen, K.; Sprong, H. Melting pot of tick-borne zoonoses: The European hedgehog contributes to the maintenance of various tick-borne diseases in natural cycles urban and suburban areas. Parasites Vectors 2017, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Marié, J.L.; Davoust, B.; Socolovschi, C.; Raoult, D.; Parola, P. Molecular detection of rickettsial agents in ticks and fleas collected from a European hedgehog (Erinaceus europaeus) in Marseilles, France. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 77–79. [Google Scholar] [CrossRef]
- Harris, E.K.; Verhoeve, V.I.; Banajee, K.H.; Macaluso, J.A.; Azad, A.F.; Macaluso, K.R. Comparative vertical transmission of Rickettsia by Dermacentor variabilis and Amblyomma maculatum. Ticks Tick Borne Dis. 2017, 8, 598–604. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
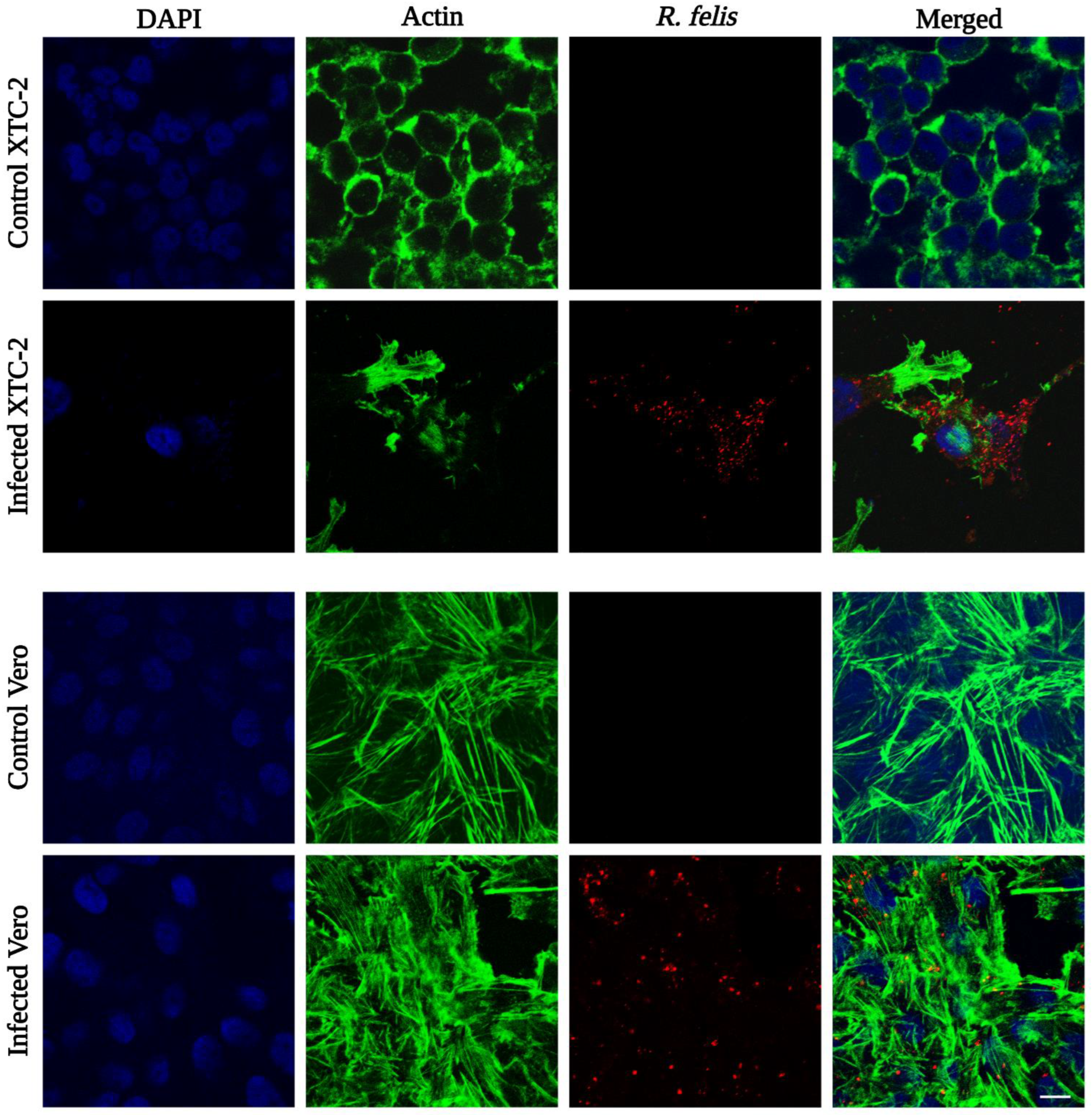{kind=link}
{kind=link}
| Strain | Year | Origin | Cell Culture | Reference |
|---|---|---|---|---|
| URRWXCal2 | 2001 | Cat flea of the colony maintained by Flea Data, Inc., in the United States | Xenopus laevis cell line XTC-2 (28 °C) | Raoult D. et al. Emerg Infect Dis 7(1): 73–81 [51] |
| Pedreira | 2006 | Cat flea collected from a naturally infested farm dog, in Brazil | Aedes albopictus cell line C6/36 (25 °C) | Horta M.C. et al. Appl Environ Microbiol 72(2): 1705–1707 [57] |
| LSU | 2006 | Cat flea of the laboratory colony maintained at the Louisiana State University, in the United States | Ixodes scapularis cell line ISE6 (32 °C) | Pornwiroon W. et al. Appl Environ Microbiol 72(8): 5589–5595 [55] |
| LSU-Lb | 2011 | Booklouse Liposcelis bostrychophila from shredded corn cobs, in the United States | Ixodes scapularis cell line ISE6 (32 °C) | Thepparit C. et al. PLoS ONE 6(1): e16396 [56] |
| CfCR(SJ) | 2011 | Cat flea collected from a domestic dog, in Costa Rica | Aedes albopictus cell line C6/36 (28 °C) | Hun L. et al. Vector Borne Zoonotic Dis 11(10): 1395–1397 [58] |
| N/A * | 2013 | Cat flea collected from a pound dog, and the flea laboratory colony maintained at the University of Queensland, in Australia | Xenopus laevis cell line XTC-2 (28 °C) | Hii S.F. et al. Parasit Vectors 6:159 [53] |
| N/A * | 2020 | Domestic dog infected with R. felis in a laboratory setting, in Australia | Xenopus laevis cell line XTC-2 (28 °C) | Ng-Nguyen D. et al. Sci Rep 10: 4151 [54] |
| Danube | This study | Questing Ixodes ricinus nymph, in Slovakia | Xenopus laevis cell line XTC-2 (28 °C) | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danchenko, M.; Benada, O.; Škultéty, Ľ.; Sekeyová, Z. Culture Isolate of Rickettsia felis from a Tick. Int. J. Environ. Res. Public Health 2022, 19, 4321. https://doi.org/10.3390/ijerph19074321
Danchenko M, Benada O, Škultéty Ľ, Sekeyová Z. Culture Isolate of Rickettsia felis from a Tick. International Journal of Environmental Research and Public Health. 2022; 19(7):4321. https://doi.org/10.3390/ijerph19074321
Chicago/Turabian StyleDanchenko, Monika, Oldřich Benada, Ľudovít Škultéty, and Zuzana Sekeyová. 2022. "Culture Isolate of Rickettsia felis from a Tick" International Journal of Environmental Research and Public Health 19, no. 7: 4321. https://doi.org/10.3390/ijerph19074321
APA StyleDanchenko, M., Benada, O., Škultéty, Ľ., & Sekeyová, Z. (2022). Culture Isolate of Rickettsia felis from a Tick. International Journal of Environmental Research and Public Health, 19(7), 4321. https://doi.org/10.3390/ijerph19074321