
The Interaction between Serotonin Transporter Allelic Variation and Maternal Care Modulates Instagram Sociability in a Sample of Singaporean Users

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Procedure
2.3. Parental Bonding
2.4. Instagram Parameters
2.5. Genotyping
2.6. Analysis
3. Results
3.1. Instagram Variables: Preliminary Results
3.2. Instagram Interaction Effects
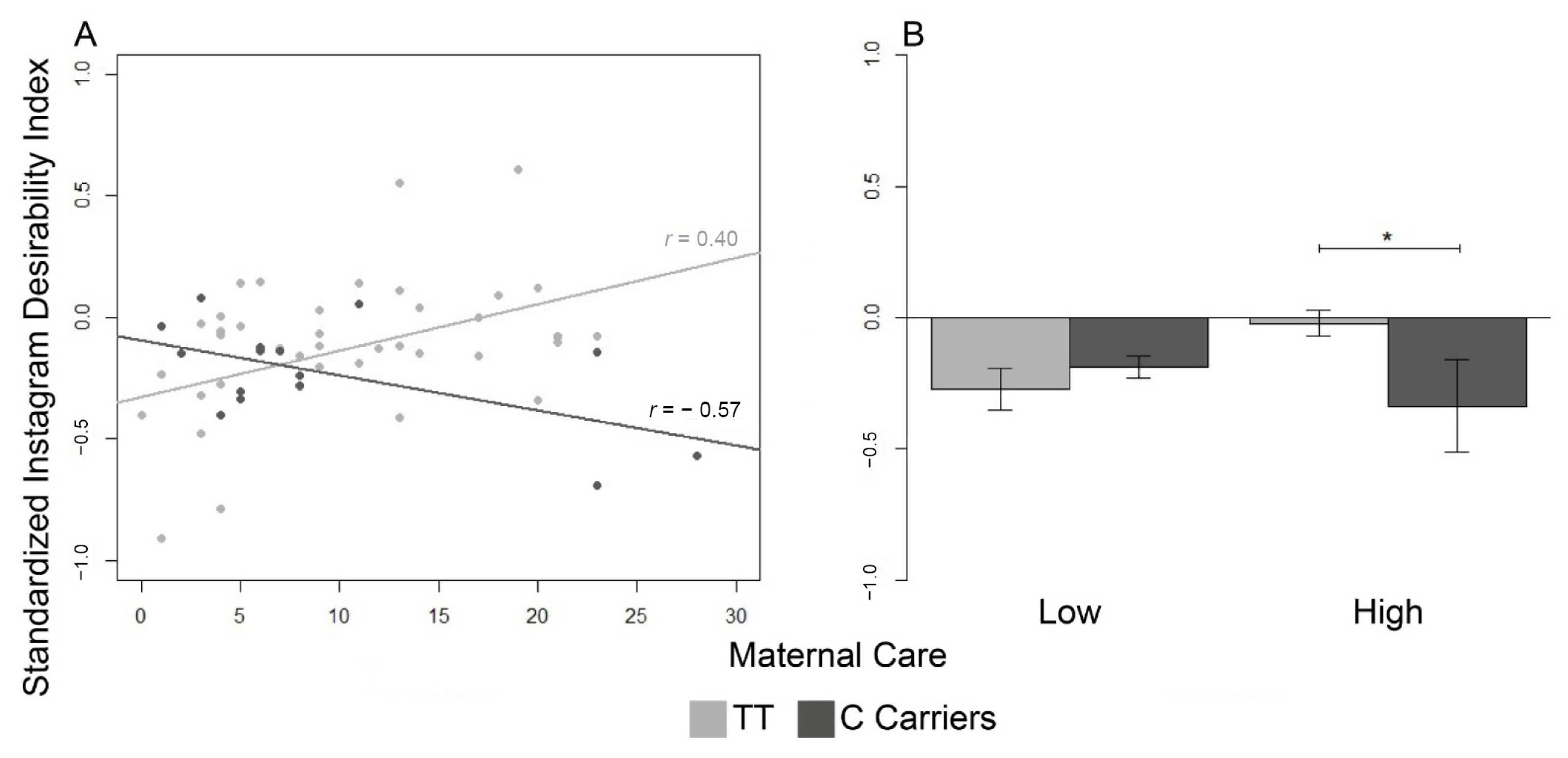
3.2.1. Social Desirability Index
3.2.2. Number of Instagram Posts, Followings, and Followers
4. Discussion
4.1. Possible Implications in Brain Mechanisms
4.2. The Importance of Social Desirability
5. Related Works

{kind=link}
| N | Reference | Genetic Feature | Environmental Factor | Results |
|---|---|---|---|---|
| 1 | Bonassi et al., 2020a [44] | OXTr rs2254298 (A/G) | Maternal Care; Maternal Overprotection; Paternal Care; Paternal Overprotection | A-carriers for OXTr rs2254298 with low paternal care showed a fewer number of posts on Instagram; (ii) effects of the interaction between OXTr rs2254298 and maternal overprotection on SDI scores |
| 2 | Bonassi et al., 2020b [66] | rs25531 (T/C) | Confidence; Need for approval; Relationships as secondary; Worry about relationships; Discomfort with closeness | Greater confidence levels in rs25531 T/T carriers are associated with greater number of followings. |
| 3 | Bonassi et al., 2021a [67] | rs25531 (T/C) | Avoidance; Anxiety | Absence of gene-by-environment interaction; association between Avoidance and “followings” |
| 4 | Carollo et al., 2021b [87] | OXTr rs53576 (A/G) | Avoidance; Anxiety | A/A carriers for OXTr rs53576 showed greater numbers of “followings” than G-carriers independent of the anxiety or avoidance levels |
| 5 | Present study | rs25531 (T/C) | Maternal Care; Maternal Overprotection; Paternal Care; Paternal Overprotection | Effects of the interaction between T/T and positive maternal care associated with greater values of SDI |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SNSs | Social Network Sites |
| IG | |
| 5-HTTLPR | Serotonin Transporter Gene-Linked Polymorphic Region |
| SNP | Single-Nucleotide Polymorphisms |
| PBI | Parental Bonding Instrument |
| SDI | Social Desirability Index |
References
- Bornstein, M.H. Handbook of Parenting: Children and Parenting; Psychology Press: Hove, UK, 2005; Volume I. [Google Scholar]
- Stocker, C.M.; Masarik, A.S.; Widaman, K.F.; Reeb, B.T.; Boardman, J.D.; Smolen, A.; Neppl, T.K.; Conger, K.J. Parenting and adolescents’ psychological adjustment: Longitudinal moderation by adolescents’ genetic sensitivity. Dev. Psychopathol. 2017, 29, 1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, M.H.; Putnick, D.L.; Hahn, C.S.; Tamis-LeMonda, C.S.; Esposito, G. Stabilities of infant behaviors and maternal responses to them. Infancy 2020, 25, 226–245. [Google Scholar] [CrossRef] [PubMed]
- Dalsant, A.; Truzzi, A.; Setoh, P.; Esposito, G. Maternal bonding in childhood moderates autonomic responses to distress stimuli in adult males. Behav. Brain Res. 2015, 292, 428–431. [Google Scholar] [CrossRef]
- Calvert, S.L. Children and digital media. In Handbook of Child Psychology and Developmental Science; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; pp. 1–41. [Google Scholar]
- Kabali, H.K.; Irigoyen, M.M.; Nunez-Davis, R.; Budacki, J.G.; Mohanty, S.H.; Leister, K.P.; Bonner, R.L. Exposure and use of mobile media devices by young children. Pediatrics 2015, 136, 1044–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meshi, D.; Tamir, D.I.; Heekeren, H.R. The emerging neuroscience of social media. Trends Cogn. Sci. 2015, 19, 771–782. [Google Scholar] [CrossRef]
- Cataldo, I.; Lepri, B.; Neoh, M.J.Y.; Esposito, G. Social Media Usage and Development of Psychiatric Disorders in Childhood and Adolescence: A Review. Front. Psychiatry 2020, 11, 1332. [Google Scholar] [CrossRef]
- Meier, A.; Schäfer, S. The positive side of social comparison on social network sites: How envy can drive inspiration on Instagram. Cyberpsychol. Behav. Soc. Netw. 2018, 21, 411–417. [Google Scholar] [CrossRef]
- Frison, E.; Eggermont, S. Browsing, posting, and liking on Instagram: The reciprocal relationships between different types of Instagram use and adolescents’ depressed mood. Cyberpsychol. Behav. Soc. Netw. 2017, 20, 603–609. [Google Scholar] [CrossRef]
- Ponnusamy, S.; Iranmanesh, M.; Foroughi, B.; Hyun, S.S. Drivers and outcomes of Instagram Addiction: Psychological well-being as moderator. Comput. Hum. Behav. 2020, 107, 106294. [Google Scholar] [CrossRef]
- Esposito, G.; Setoh, P.; Shinohara, K.; Bornstein, M.H. The development of attachment: Integrating genes, brain, behavior, and environment. Behav. Brain Res. 2017, 325, 87–89. [Google Scholar] [CrossRef] [Green Version]
- Belsky, J.; Pluess, M. Beyond diathesis stress: Differential susceptibility to environmental influences. Psychol. Bull. 2009, 135, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canli, T.; Lesch, K.P. Long story short: The serotonin transporter in emotion regulation and social cognition. Nat. Neurosci. 2007, 10, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Beevers, C.G.; Wells, T.T.; Ellis, A.J.; McGeary, J.E. Association of the serotonin transporter gene promoter region (5-HTTLPR) polymorphism with biased attention for emotional stimuli. J. Abnorm. Psychol. 2009, 118, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad-Zadeh, L.; Moses, L.; Gwaltney-Brant, S. Serotonin: A review. J. Vet. Pharmacol. Ther. 2008, 31, 187–199. [Google Scholar] [CrossRef]
- Kraft, J.B.; Slager, S.L.; McGrath, P.J.; Hamilton, S.P. Sequence analysis of the serotonin transporter and associations with antidepressant response. Biol. Psychiatry 2005, 58, 374–381. [Google Scholar] [CrossRef]
- Wendland, J.; Martin, B.; Kruse, M.; Lesch, K.; Murphy, D. Simultaneous genotyping of four functional loci of human SLC6A4, with a reappraisal of 5-HTTLPR and rs25531. Mol. Psychiatry 2006, 11, 224–226. [Google Scholar] [CrossRef]
- Morgan, J.E.; Hammen, C.; Lee, S.S. Parental serotonin transporter polymorphism (5-HTTLPR) moderates associations of stress and child behavior with parenting behavior. J. Clin. Child Adolesc. Psychol. 2018, 47, S76–S87. [Google Scholar] [CrossRef]
- Ford, B.Q.; Mauss, I.B.; Troy, A.S.; Smolen, A.; Hankin, B. Emotion regulation moderates the risk associated with the 5-HTT gene and stress in children. Emotion 2014, 14, 930. [Google Scholar] [CrossRef] [Green Version]
- Gazor, A.; Fang, H.; Gagne, J.R. An exploratory genetic analysis: Associations between parent depression symptoms, child temperament, and the serotonin transporter gene polymorphism (5 HTTLPR). J. Appl. Biobehav. Res. 2017, 22, e12099. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Gabrieli, G.; Lepri, B.; Esposito, G. Serotonin Transporter Gene Polymorphisms and Maternal Overprotection Regulate Adult Social Expectations on Close Relationships. Brain Sci. 2021, 11, 1123. [Google Scholar] [CrossRef]
- Madrid, J.E.; Mandalaywala, T.M.; Coyne, S.P.; Ahloy-Dallaire, J.; Garner, J.P.; Barr, C.S.; Maestripieri, D.; Parker, K.J. Adaptive developmental plasticity in rhesus macaques: The serotonin transporter gene interacts with maternal care to affect juvenile social behaviour. Proc. R. Soc. B 2018, 285, 20180541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truzzi, A.; Bornstein, M.H.; Senese, V.P.; Shinohara, K.; Setoh, P.; Esposito, G. Serotonin transporter gene polymorphisms and early parent-infant interactions are related to adult male heart rate response to female crying. Front. Physiol. 2017, 8, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraguti, G.; Pascale, E.; Lucarelli, M. Alcohol addiction: A molecular biology perspective. Curr. Med. Chem. 2015, 22, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Schiele, M.A.; Ziegler, C.; Holitschke, K.; Schartner, C.; Schmidt, B.; Weber, H.; Reif, A.; Romanos, M.; Pauli, P.; Zwanzger, P.; et al. Influence of 5-HTT variation, childhood trauma and self-efficacy on anxiety traits: A gene-environment-coping interaction study. J. Neural Transm. 2016, 123, 895–904. [Google Scholar] [CrossRef]
- Niitsu, K.; Rice, M.J.; Houfek, J.F.; Stoltenberg, S.F.; Kupzyk, K.A.; Barron, C.R. A systematic review of genetic influence on psychological resilience. Biol. Res. Nurs. 2019, 21, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonassi, A.; Cataldo, I.; Tandiono, M.; Foo, J.; Lepri, B.; Esposito, G.P. 112 Effect of early paternal caregiving and genotype rs25531 polymorphisms on the adult relationship with the partner. Eur. Neuropsychopharmacol. 2021, 44, S9–S10. [Google Scholar] [CrossRef]
- Sturge-Apple, M.L.; Cicchetti, D.; Davies, P.T.; Suor, J.H. Differential susceptibility in spillover between interparental conflict and maternal parenting practices: Evidence for OXTR and 5-HTT genes. J. Fam. Psychol. 2012, 26, 431. [Google Scholar] [CrossRef] [Green Version]
- Wüst, S.; Kumsta, R.; Treutlein, J.; Frank, J.; Entringer, S.; Schulze, T.G.; Rietschel, M. Sex-specific association between the 5-HTT gene-linked polymorphic region and basal cortisol secretion. Psychoneuroendocrinology 2009, 34, 972–982. [Google Scholar] [CrossRef]
- Fandiño-Losada, A.; Wei, Y.; Åberg, E.; Sjöholm, L.K.; Lavebratt, C.; Forsell, Y. Influence of serotonin transporter promoter variation on the effects of separation from parent/partner on depression. J. Affect. Disord. 2013, 144, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Steiger, H.; Richardson, J.; Joober, R.; Gauvin, L.; Israel, M.; Bruce, K.R.; Kin, N.N.Y.; Howard, H.; Young, S.N. The 5HTTLPR polymorphism, prior maltreatment and dramatic–erratic personality manifestations in women with bulimic syndromes. J. Psychiatry Neurosci. 2007, 32, 354. [Google Scholar]
- Cataldo, I.; Azhari, A.; Lepri, B.; Esposito, G. Oxytocin receptors (OXTR) and early parental care: An interaction that modulates psychiatric disorders. Res. Dev. Disabil. 2018, 82, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Carollo, A.; Bonassi, A.; Lim, M.; Gabrieli, G.; Setoh, P.; Dimitriou, D.; Aryadoust, V.; Esposito, G. Developmental disabilities across the world: A scientometric review from 1936 to 2020. Res. Dev. Disabil. 2021, 117, 104031. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; Sugden, K.; Moffitt, T.E.; Taylor, A.; Craig, I.W.; Harrington, H.; McClay, J.; Mill, J.; Martin, J.; Braithwaite, A.; et al. Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene. Science 2003, 301, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Belsky, J.; Bakermans-Kranenburg, M.J.; Van IJzendoorn, M.H. For better and for worse: Differential susceptibility to environmental influences. Curr. Dir. Psychol. Sci. 2007, 16, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Oxytocin receptor (OXTR) and serotonin transporter (5-HTT) genes associated with observed parenting. Soc. Cogn. Affect. Neurosci. 2008, 3, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.W.; Harris, T.O. Depression and the serotonin transporter 5-HTTLPR polymorphism: A review and a hypothesis concerning gene–environment interaction. J. Affect. Disord. 2008, 111, 1–12. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Tandiono, M.; Foo, J.; Lepri, B.; Esposito, G.P. 0516 Genotype rs25531 polymorphisms and quality in peer relationships adjust the neural response of the anterior prefrontal cortex to cry. Eur. Neuropsychopharmacol. 2021, 53, S380–S381. [Google Scholar] [CrossRef]
- Parker, G.; Tupling, H.; Brown, L.B. A parental bonding instrument. Br. J. Med. Psychol. 1979, 52, 1–10. [Google Scholar] [CrossRef]
- Parker, G. The parental bonding instrument: Psychometric properties reviewed. Psychiatr. Dev. 1989, 7, 317–335. [Google Scholar]
- Parker, G. The Parental Bonding Instrument: A decade of research. Soc. Psychiatry Psychiatr. Epidemiol. Int. J. Res. Soc. Genet. Epidemiol. Ment. Health Serv. 1990, 25, 281–282. [Google Scholar] [CrossRef]
- Bonassi, A.; Ghilardi, T.; Truzzi, A.; Cataldo, I.; Azhari, A.; Setoh, P.; Shinohara, K.; Esposito, G. Dataset on genetic and physiological adults’ responses to social distress. Data Brief 2017, 13, 742–748. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Gabrieli, G.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin receptor gene polymorphisms and early parental bonding interact in shaping instagram social behavior. Int. J. Environ. Res. Public Health 2020, 17, 7232. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Wang, N.; Kong, L.; Dong, X.; Tian, L. A large number of online friends and a high frequency of social interaction compensate for each Other’s shortage in regard to perceived social support. Curr. Psychol. 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gil de Zúñiga, H.; Diehl, T.; Huber, B.; Liu, J. Personality traits and social media use in 20 countries: How personality relates to frequency of social media use, social media news use, and social media use for social interaction. Cyberpsychol. Behav. Soc. Netw. 2017, 20, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longobardi, C.; Settanni, M.; Fabris, M.; Marengo, D. Follow or be followed: Exploring the links between Instagram popularity, social media addiction, cyber victimization, and subjective happiness in Italian adolescents. Child. Youth Serv. Rev. 2020, 113, 104955. [Google Scholar] [CrossRef]
- Quercia, D.; Ellis, J.; Capra, L. Using mobile phones to nurture social networks. IEEE Pervasive Comput. 2010, 9, 12–20. [Google Scholar] [CrossRef]
- Quercia, D.; Bodaghi, M.; Crowcroft, J. Loosing “friends” on Facebook. In Proceedings of the 4th Annual ACM Web Science Conference, New York, NY, USA, 22–24 June 2012; pp. 251–254. [Google Scholar]
- Pittman, M.; Abell, A. More Trust in Fewer Followers: Diverging Effects of Popularity Metrics and Green Orientation Social Media Influencers. J. Interact. Mark. 2021, 56, 70–82. [Google Scholar] [CrossRef]
- Trifiro, B.M.; Gerson, J. Social media usage patterns: Research note regarding the lack of universal validated measures for active and passive use. Soc. Media Soc. 2019, 5, 2056305119848743. [Google Scholar] [CrossRef]
- Reece, A.G.; Danforth, C.M. Instagram photos reveal predictive markers of depression. EPJ Data Sci. 2017, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Han, K.; Shih, P.C.; Lee, D. Generation like: Comparative characteristics in Instagram. In Proceedings of the 33rd Annual ACM Conference on Human Factors in Computing Systems, Seoul, Korea, 18–23 April 2015; pp. 4039–4042. [Google Scholar]
- Thorisdottir, I.E.; Sigurvinsdottir, R.; Asgeirsdottir, B.B.; Allegrante, J.P.; Sigfusdottir, I.D. Active and passive social media use and symptoms of anxiety and depressed mood among Icelandic adolescents. Cyberpsychol. Behav. Soc. Netw. 2019, 22, 535–542. [Google Scholar] [CrossRef]
- Dumas, T.M.; Maxwell-Smith, M.; Davis, J.P.; Giulietti, P.A. Lying or longing for likes? Narcissism, peer belonging, loneliness and normative versus deceptive like-seeking on Instagram in emerging adulthood. Comput. Hum. Behav. 2017, 71, 1–10. [Google Scholar] [CrossRef]
- Ben-Gal, I. Outlier detection. In Data Mining and Knowledge Discovery Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 131–146. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Kiser, D.; SteemerS, B.; Branchi, I.; Homberg, J.R. The reciprocal interaction between serotonin and social behaviour. Neurosci. Biobehav. Rev. 2012, 36, 786–798. [Google Scholar] [CrossRef]
- Belsky, J.; Jonassaint, C.; Pluess, M.; Stanton, M.; Brummett, B.; Williams, R. Vulnerability genes or plasticity genes? Mol. Psychiatry 2009, 14, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Blaya, C.; Salum, G.A.; Moorjani, P.; Seganfredo, A.C.; Heldt, E.; Leistner-Segal, S.; Smoller, J.W.; Manfro, G.G. Panic disorder and serotonergic genes (SLC6A4, HTR1A and HTR2A): Association and interaction with childhood trauma and parenting. Neurosci. Lett. 2010, 485, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Flasbeck, V.; Moser, D.; Pakusch, J.; Kumsta, R.; Brüne, M. The association between childhood maltreatment and empathic perspective taking is moderated by the 5-HTT linked polymorphic region: Another example of “differential susceptibility”. PLoS ONE 2019, 14, e0226737. [Google Scholar] [CrossRef]
- Wang, M.; Tian, X.; Zhang, W. Interactions between the combined genotypes of 5-HTTLPR and BDNF Val66Met polymorphisms and parenting on adolescent depressive symptoms: A three-year longitudinal study. J. Affect. Disord. 2020, 265, 104–111. [Google Scholar] [CrossRef]
- Miller, G.; Zhu, G.; Wright, M.J.; Hansell, N.K.; Martin, N.G. The heritability and genetic correlates of mobile phone use: A twin study of consumer behavior. Twin Res. Hum. Genet. 2012, 15, 97–106. [Google Scholar] [CrossRef]
- Deryakulu, D.; Ursavaş, Ö.F. Genetic and environmental influences on problematic Internet use: A twin study. Comput. Hum. Behav. 2014, 39, 331–338. [Google Scholar] [CrossRef]
- York, C. A regression approach to testing genetic influence on communication behavior: Social media use as an example. Comput. Hum. Behav. 2017, 73, 100–109. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Giulio, G.; Foo, J.N.; Lepri, B.; Gianluca, E.P. 677 Serotonin transporter gene polymorphisms and confidence in adult relationships affect the number of followed people on Instagram. In Proceedings of the 33rd ECNP Congress—Posters, Virtual, 12–15 September 2020; Volume 40, pp. S385–S386. [Google Scholar]
- Bonassi, A.; Carollo, A.; Cataldo, I.; Gabrieli, G.; Tandiono, M.; Foo, J.N.; Lepri, B.; Esposito, G. Modulation of Instagram Number of Followings by Avoidance in Close Relationships in Young Adults under a Gene x Environment Perspective. Int. J. Environ. Res. Public Health 2021, 18, 7547. [Google Scholar] [CrossRef] [PubMed]
- Pérez, S.P. Serotonin and emotional decision-making. In Serotonin; IntechOpen: London, UK, 2018. [Google Scholar]
- Puig, M.; Gulledge, A.T. Serotonin and prefrontal cortex function: Neurons, networks, and circuits. Mol. Neurobiol. 2011, 44, 449–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, E.A.; Wise, S.P. Interactions between orbital prefrontal cortex and amygdala: Advanced cognition, learned responses and instinctive behaviors. Curr. Opin. Neurobiol. 2010, 20, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, E.E.; Guyer, A.E. The development of the ventral prefrontal cortex and social flexibility. Dev. Cogn. Neurosci. 2011, 1, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.N. Social cognition and the prefrontal cortex. Behav. Cogn. Neurosci. Rev. 2003, 2, 97–114. [Google Scholar] [CrossRef]
- Gillath, O.; Bunge, S.A.; Shaver, P.R.; Wendelken, C.; Mikulincer, M. Attachment-style differences in the ability to suppress negative thoughts: Exploring the neural correlates. Neuroimage 2005, 28, 835–847. [Google Scholar] [CrossRef]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social reward requires coordinated activity of nucleus accumbens oxytocin and serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Carollo, A.; Balagtas, J.P.M.; Neoh, M.J.Y.; Esposito, G. A scientometric approach to review the role of the medial preoptic area (MPOA) in parental behavior. Brain Sci. 2021, 11, 393. [Google Scholar] [CrossRef]
- Olazábal, D.; Abercrombie, E.; Rosenblatt, J.; Morrell, J. The content of dopamine, serotonin, and their metabolites in the neural circuit that mediates maternal behavior in juvenile and adult rats. Brain Res. Bull. 2004, 63, 259–268. [Google Scholar] [CrossRef]
- Cataldo, I.; Neoh, M.J.Y.; Chew, W.F.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin receptor gene and parental bonding modulate prefrontal responses to cries: A NIRS Study. Sci. Rep. 2020, 10, 8588. [Google Scholar] [CrossRef]
- Sherman, L.E.; Greenfield, P.M.; Hernandez, L.M.; Dapretto, M. Peer influence via instagram: Effects on brain and behavior in adolescence and young adulthood. Child Dev. 2018, 89, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Dumas, T.M.; Maxwell-Smith, M.A.; Tremblay, P.F.; Litt, D.M.; Ellis, W. Gaining likes, but at what cost? Longitudinal relations between young adults’ deceptive like-seeking on instagram, peer belonging and self-esteem. Comput. Hum. Behav. 2020, 112, 106467. [Google Scholar] [CrossRef]
- Sherman, L.E.; Payton, A.A.; Hernandez, L.M.; Greenfield, P.M.; Dapretto, M. The power of the like in adolescence: Effects of peer influence on neural and behavioral responses to social media. Psychol. Sci. 2016, 27, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Fisher, H. Lust, attraction, attachment: Biology and evolution of the three primary emotion systems for mating, reproduction, and parenting. J. Sex Educ. Ther. 2000, 25, 96–104. [Google Scholar] [CrossRef]
- Stoltenberg, S.F.; Christ, C.C.; Carlo, G. Afraid to help: Social anxiety partially mediates the association between 5-HTTLPR triallelic genotype and prosocial behavior. Soc. Neurosci. 2013, 8, 400–406. [Google Scholar] [CrossRef]
- Jackson, C.A.; Luchner, A.F. Self-presentation mediates the relationship between self-criticism and emotional response to Instagram feedback. Personal. Individ. Differ. 2018, 133, 1–6. [Google Scholar] [CrossRef]
- Ferrara, E.; Interdonato, R.; Tagarelli, A. Online popularity and topical interests through the lens of instagram. In Proceedings of the 25th ACM Conference on Hypertext and Social Media, Santiago, Chile, 1–4 September 2014; pp. 24–34. [Google Scholar]
- Ivorra, J.L.; Sanjuan, J.; Jover, M.; Carot, J.M.; de Frutos, R.; Molto, M.D. Gene-environment interaction of child temperament. J. Dev. Behav. Pediatr. 2010, 31, 545–554. [Google Scholar] [CrossRef]
- Mileva-Seitz, V.; Kennedy, J.; Atkinson, L.; Steiner, M.; Levitan, R.; Matthews, S.G.; Meaney, M.J.; Sokolowski, M.B.; Fleming, A.S. Serotonin transporter allelic variation in mothers predicts maternal sensitivity, behavior and attitudes toward 6-month-old infants. Genes Brain Behav. 2011, 10, 325–333. [Google Scholar] [CrossRef]
- Carollo, A.; Bonassi, A.; Cataldo, I.; Gabrieli, G.; Tandiono, M.; Foo, J.N.; Lepri, B.; Esposito, G. The relation between Oxytocin Receptor Gene polymorphisms, adult attachment and Instagram sociability: An exploratory analysis. Heliyon 2021, 7, e07894. [Google Scholar] [CrossRef]
- Ling Ooi, G. The role of the developmental state and interethnic relations in Singapore. Asian Ethn. 2005, 6, 109–120. [Google Scholar] [CrossRef]
- Jiang, S.; Ngien, A. The effects of Instagram use, social comparison, and self-esteem on social anxiety: A survey study in Singapore. Soc. Media Soc. 2020, 6, 2056305120912488. [Google Scholar] [CrossRef]
- Sheldon, P.; Bryant, K. Instagram: Motives for its use and relationship to narcissism and contextual age. Comput. Hum. Behav. 2016, 58, 89–97. [Google Scholar] [CrossRef]
- Sen, I.; Aggarwal, A.; Mian, S.; Singh, S.; Kumaraguru, P.; Datta, A. Worth its weight in likes: Towards detecting fake likes on instagram. In Proceedings of the 10th ACM Conference on Web Science, Amsterdam, The Netherlands, 27–30 May 2018; pp. 205–209. [Google Scholar]
- Zarei, K.; Farahbakhsh, R.; Crespi, N. Typification of impersonated accounts on instagram. In Proceedings of the 2019 IEEE 38th International Performance Computing and Communications Conference (IPCCC), London, UK, 29–31 October 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 1–6. [Google Scholar]
- Slater, M.; Guger, C.; Edlinger, G.; Leeb, R.; Pfurtscheller, G.; Antley, A.; Garau, M.; Brogni, A.; Friedman, D. Analysis of physiological responses to a social situation in an immersive virtual environment. Presence 2006, 15, 553–569. [Google Scholar] [CrossRef]
- Liu, D.; Liu, S.; Liu, X.; Zhang, C.; Li, A.; Jin, C.; Chen, Y.; Wang, H.; Zhang, X. Interactive brain activity: Review and progress on EEG-based hyperscanning in social interactions. Front. Psychol. 2018, 9, 1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, M.W.; Rodrigues, J.J.; Kumar, N.; Saleem, K.; Illin, I.V. Postpartum depression prediction through pregnancy data analysis for emotion-aware smart systems. Inf. Fusion 2019, 47, 23–31. [Google Scholar] [CrossRef]
- Carneiro, M.B.; Moreira, M.W.; Pereira, S.S.; Gallindo, E.L.; Rodrigues, J.J. Recommender System for Postpartum Depression Monitoring based on Sentiment Analysis. In Proceedings of the 2020 IEEE International Conference on E-Health Networking, Application & Services (HEALTHCOM), Shenzhen, China, 1–2 March 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 1–6. [Google Scholar]

| Variable | N | Percent | Mean | SD |
|---|---|---|---|---|
| Age | 57 | - | 20.82 | 1.59 |
| 18 y-o | 1 | 1.75 | - | - |
| 19 y-o | 10 | 17.54 | - | - |
| 20 y-o | 20 | 35.09 | - | - |
| 21 y-o | 8 | 14.04 | - | - |
| 22 y-o | 8 | 14.04 | - | - |
| 23 y-o | 6 | 10.53 | - | - |
| 24 y-o | 3 | 5.26 | - | - |
| 25 y-o | 1 | 1.75 | - | - |
| Gender | ||||
| Level | N | Percent | Mean | SD |
| Males | 16 | 28.07 | - | - |
| Females | 41 | 71.93 | - | - |
| Ethnicity | ||||
| Level | N | Percent | Mean | SD |
| Chinese | 56 | 98.25 | - | - |
| Indian | 1 | 1.75 | - | - |
| Chinese Female | 40 | 70.18 | - | - |
| Chinese Male | 16 | 28.07 | - | - |
| Indian Female | 1 | 1.75 | - | - |
| Indian Male | 0 | 0 | - | - |
| Variable | Min | 1st Q | Median | Mean | 3rd Q | Max |
|---|---|---|---|---|---|---|
| Instagram Variables | ||||||
| Posts Number | 0.33 | 0.39 | 0.49 | 0.56 | 0.67 | 1.19 |
| Followings Number | −1.54 | −0.77 | −0.12 | −0.12 | 0.52 | 1.80 |
| Followers Number | −0.80 | −0.50 | −0.16 | −0.11 | 0.22 | 1.45 |
| Social Desirability Index | −1.16 | −0.28 | −0.13 | −0.16 | −0.03 | 0.61 |
| Parental Bonding Dimensions | ||||||
| Maternal care | 0.00 | 4.00 | 8.00 | 9.97 | 14.00 | 28.00 |
| Paternal care | 0.00 | 9.00 | 14.00 | 14.49 | 18.00 | 30.00 |
| Maternal Overprotection | 8.00 | 18.00 | 23.00 | 23.82 | 30.00 | 37.00 |
| Paternal Overprotection | 11.00 | 23.00 | 29.00 | 27.05 | 32.00 | 37.00 |
| PBI Dimension | Low/TT | Low/C | High/TT | High/C |
|---|---|---|---|---|
| Number of Posts | ||||
| Maternal Care | 0.57 (0.05) | 0.56 (0.07) | 0.55 (0.05) | 0.57 (0.12) |
| Paternal Care | 0.56 (0.05) | 0.50 (0.04) | 0.57 (0.05) | 0.64 (0.12) |
| Maternal Overprotection | 0.57 (0.05) | 0.63 (0.12) | 0.56 (0.04) | 0.50 (0.04) |
| Paternal Overprotection | 0.57 (0.05) | 0.65 (0.12) | 0.55 (0.05) | 0.51 (0.06) |
| Number of Followings | ||||
| Maternal Care | 0.11 (0.19) | −0.27 (0.21) | −0.27 (0.17) | 0.01 (0.75) |
| Paternal Care | −0.19 (0.17) | −0.60 (0.18) | 0.01 (0.20) | 0.28 (0.41) |
| Maternal Overprotection | −0.09 (0.19) | −0.034 (0.39) | −0.11 (0.18) | −0.33 (0.30) |
| Paternal Overprotection | −0.12 (0.16) | 0.32 (0.34) | −0.06 (0.22) | −0.53 (0.28) |
| Number of Followers | ||||
| Maternal Care | −0.03 (0.13) | −0.19 (0.13) | −0.15 (0.10) | −0.10 (0.38) |
| Paternal Care | −0.12 (0.11) | −0.29 (0.17) | −0.07 (0.11) | −0.02 (0.20) |
| Maternal Overprotection | −0.07 (0.12) | −0.15 (0.20) | −0.13 (0.10) | −0.17 (0.19) |
| Paternal Overprotection | −0.10 (0.11) | 0.16 (0.22) | −0.09 (0.12) | −0.38 (0.13) |
| Social Desirability Index | ||||
| Maternal Care | −0.27 (0.08) | −0.19 (0.04) | −0.02 (0.05) | −0.34 (0.18) |
| Paternal Care | −0.21 (0.07) | −0.23 (0.08) | −0.05 (0.06) | −0.23 (0.08) |
| Maternal Overprotection | −0.11 (0.08) | −0.22 (0.09) | −0.16 (0.06) | −0.23 (0.07) |
| Paternal Overprotection | −0.14 (0.07) | −0.17 (0.06) | −0.13 (0.06) | −0.26 (0.08) |
| Instagram Variable | Males | Females |
|---|---|---|
| Posts Number | 0.60 (0.05) | 0.55 (0.03) |
| Followings Number | 0.32 (0.23) | −0.30 (0.12) |
| Followers Number | 0.08 (0.12) | −0.19 (0.08) |
| Social Desirability Index | −0.11 (0.03) | −0.18 (0.05) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonassi, A.; Cataldo, I.; Gabrieli, G.; Tandiono, M.; Foo, J.N.; Lepri, B.; Esposito, G. The Interaction between Serotonin Transporter Allelic Variation and Maternal Care Modulates Instagram Sociability in a Sample of Singaporean Users. Int. J. Environ. Res. Public Health 2022, 19, 5348. https://doi.org/10.3390/ijerph19095348
Bonassi A, Cataldo I, Gabrieli G, Tandiono M, Foo JN, Lepri B, Esposito G. The Interaction between Serotonin Transporter Allelic Variation and Maternal Care Modulates Instagram Sociability in a Sample of Singaporean Users. International Journal of Environmental Research and Public Health. 2022; 19(9):5348. https://doi.org/10.3390/ijerph19095348
Chicago/Turabian StyleBonassi, Andrea, Ilaria Cataldo, Giulio Gabrieli, Moses Tandiono, Jia Nee Foo, Bruno Lepri, and Gianluca Esposito. 2022. "The Interaction between Serotonin Transporter Allelic Variation and Maternal Care Modulates Instagram Sociability in a Sample of Singaporean Users" International Journal of Environmental Research and Public Health 19, no. 9: 5348. https://doi.org/10.3390/ijerph19095348
APA StyleBonassi, A., Cataldo, I., Gabrieli, G., Tandiono, M., Foo, J. N., Lepri, B., & Esposito, G. (2022). The Interaction between Serotonin Transporter Allelic Variation and Maternal Care Modulates Instagram Sociability in a Sample of Singaporean Users. International Journal of Environmental Research and Public Health, 19(9), 5348. https://doi.org/10.3390/ijerph19095348