
Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea



Abstract
:1. Introduction
2. Materials and Methods
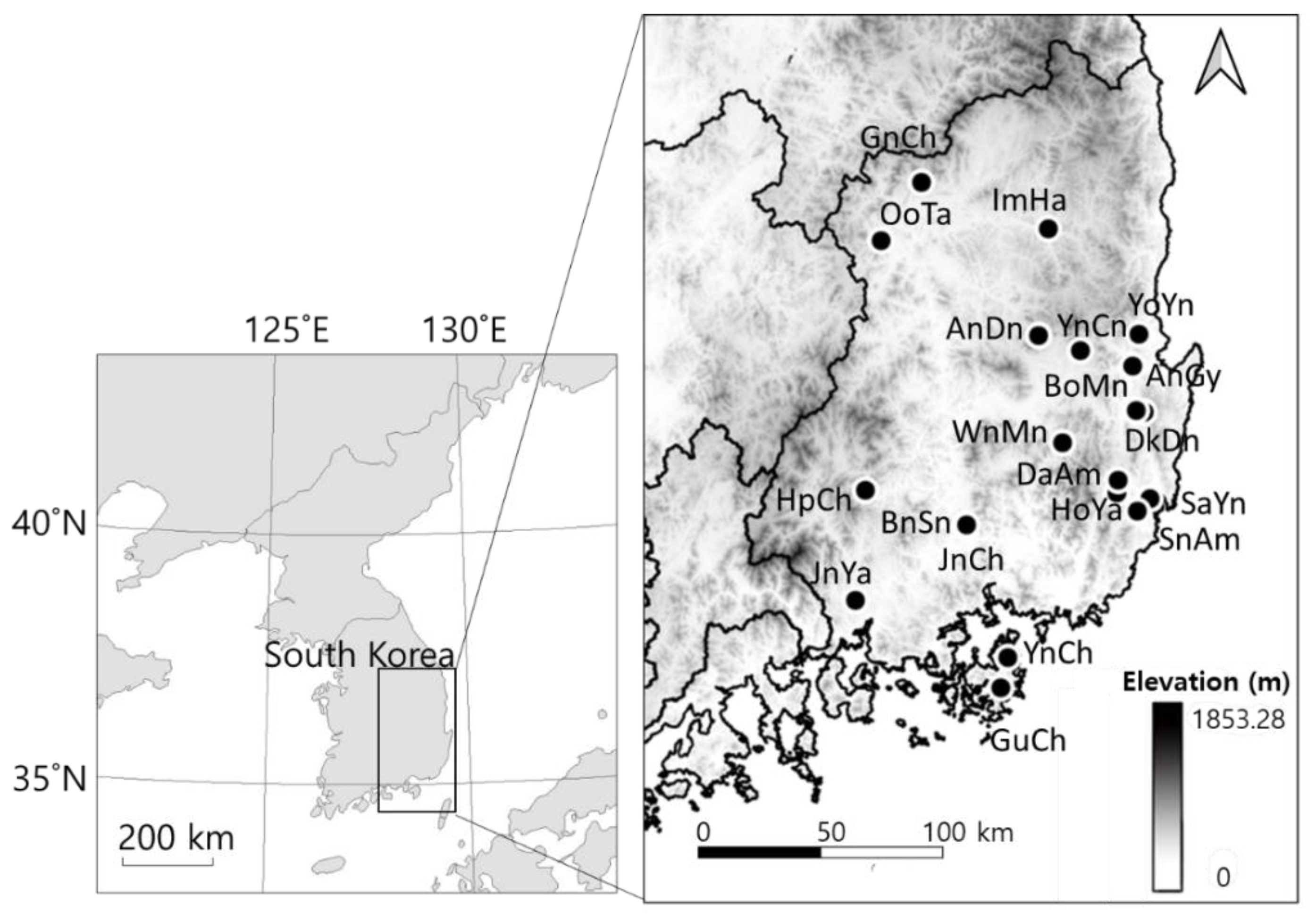
2.1. Ecological Data
2.2. Functional Traits of Macroinvertebrates
2.3. Data Analyses
2.3.1. Taxonomic Diversity and Functional Diversity
2.3.2. Community Characteristics
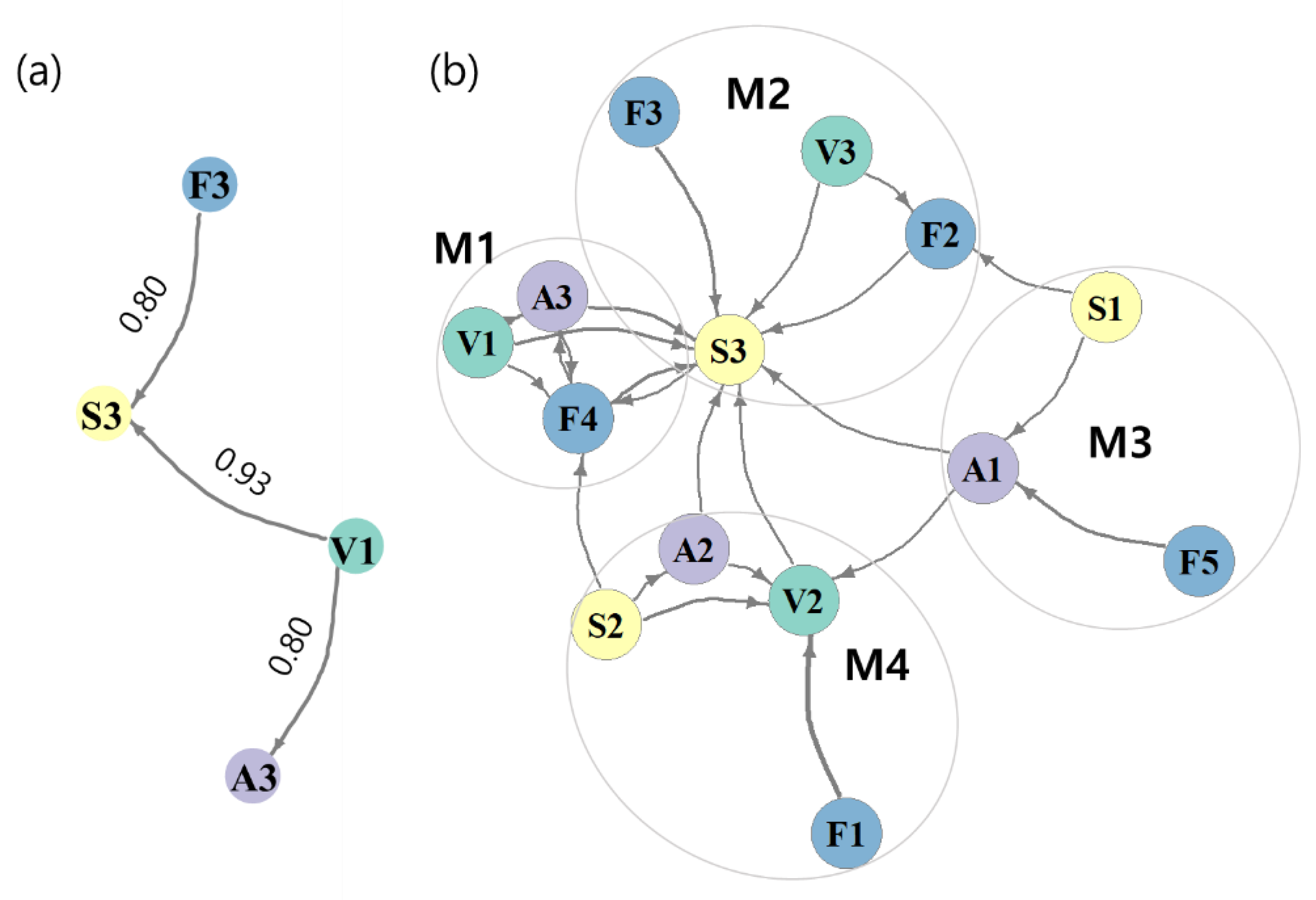
2.3.3. Functional Characteristics of Communities
3. Results
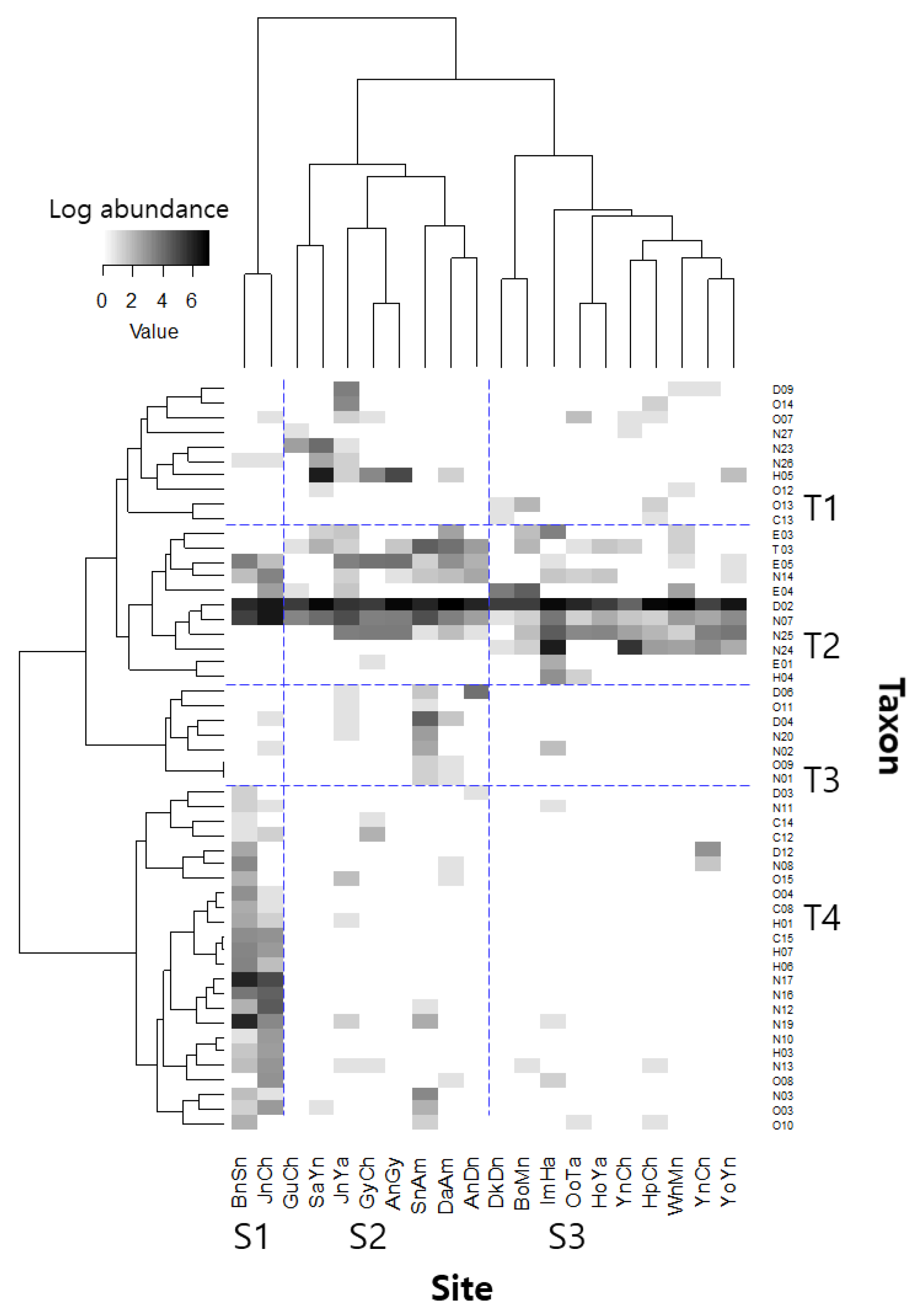
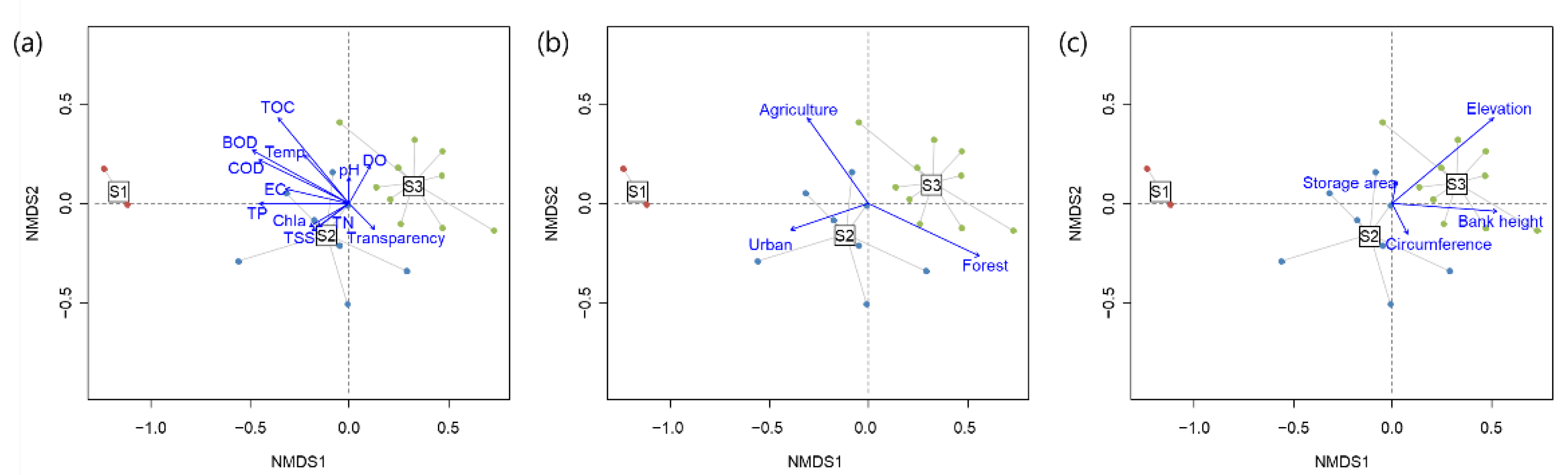
3.1. Patterns of Taxonomic Diversity
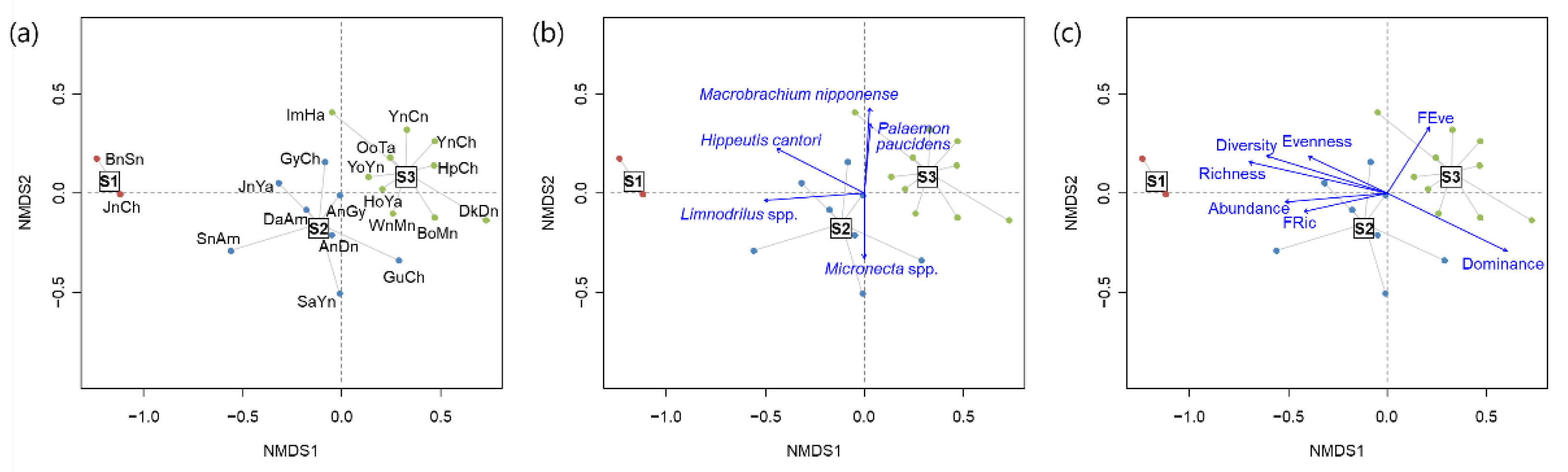
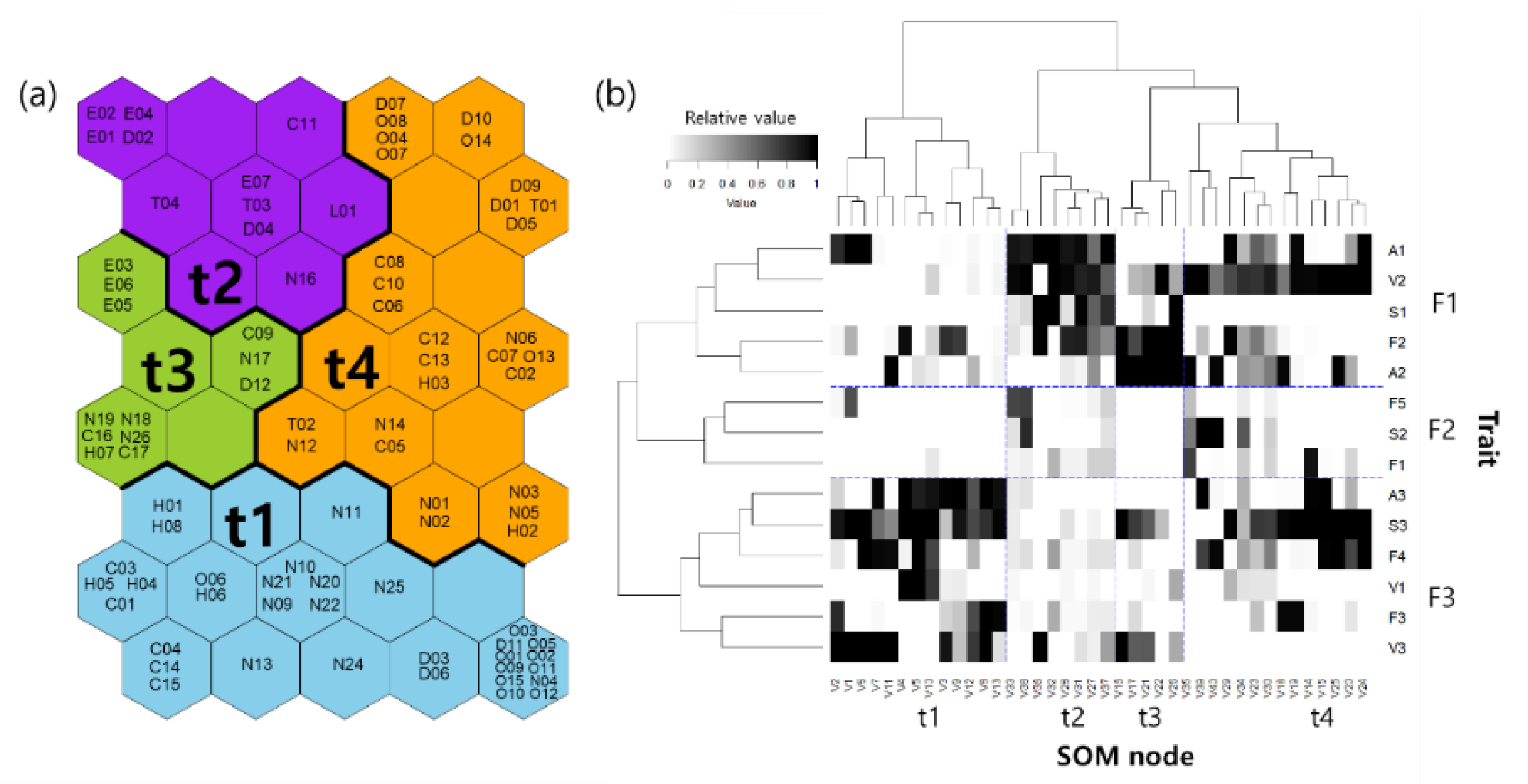
3.2. Functional Traits and Diversity
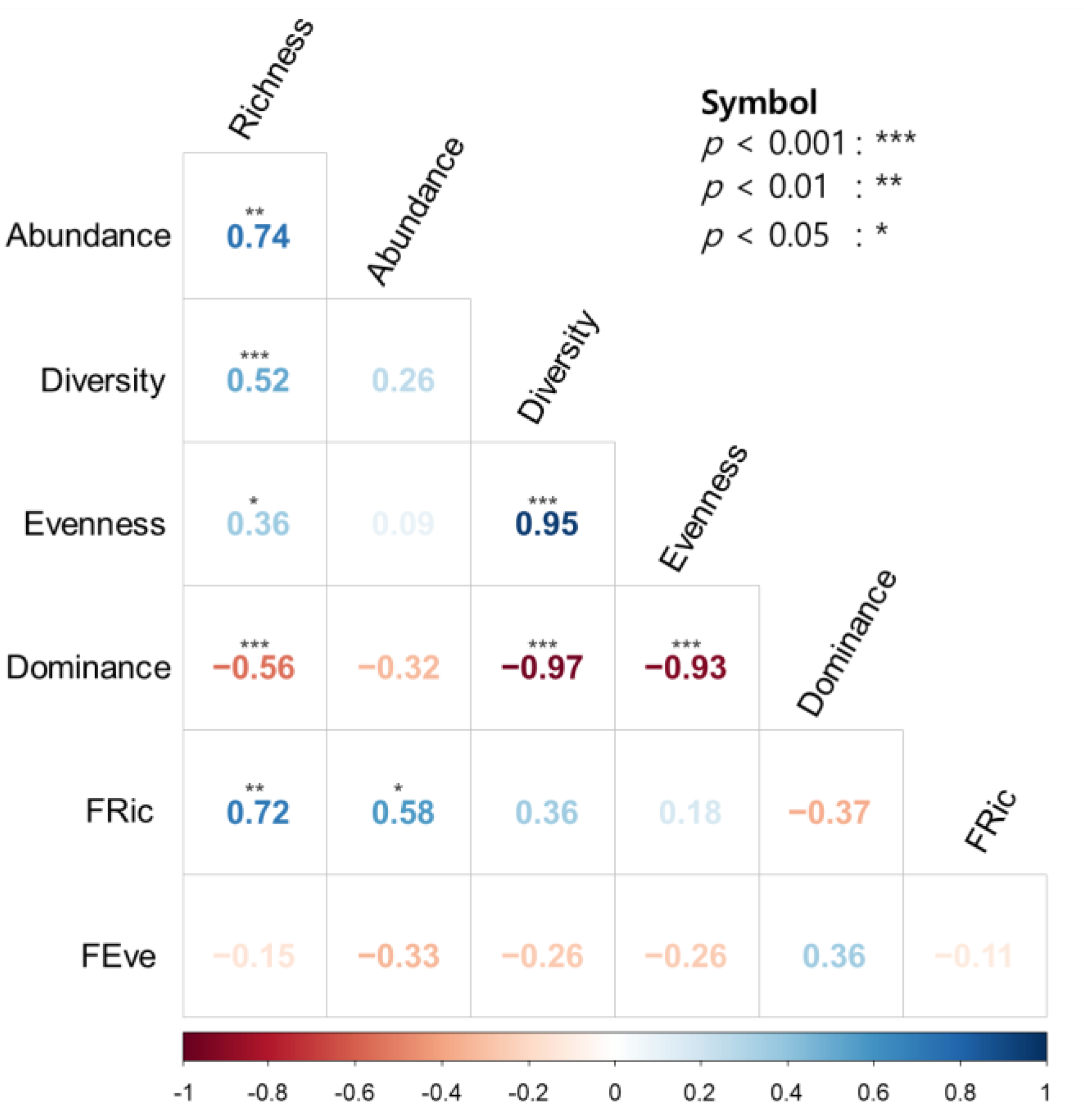
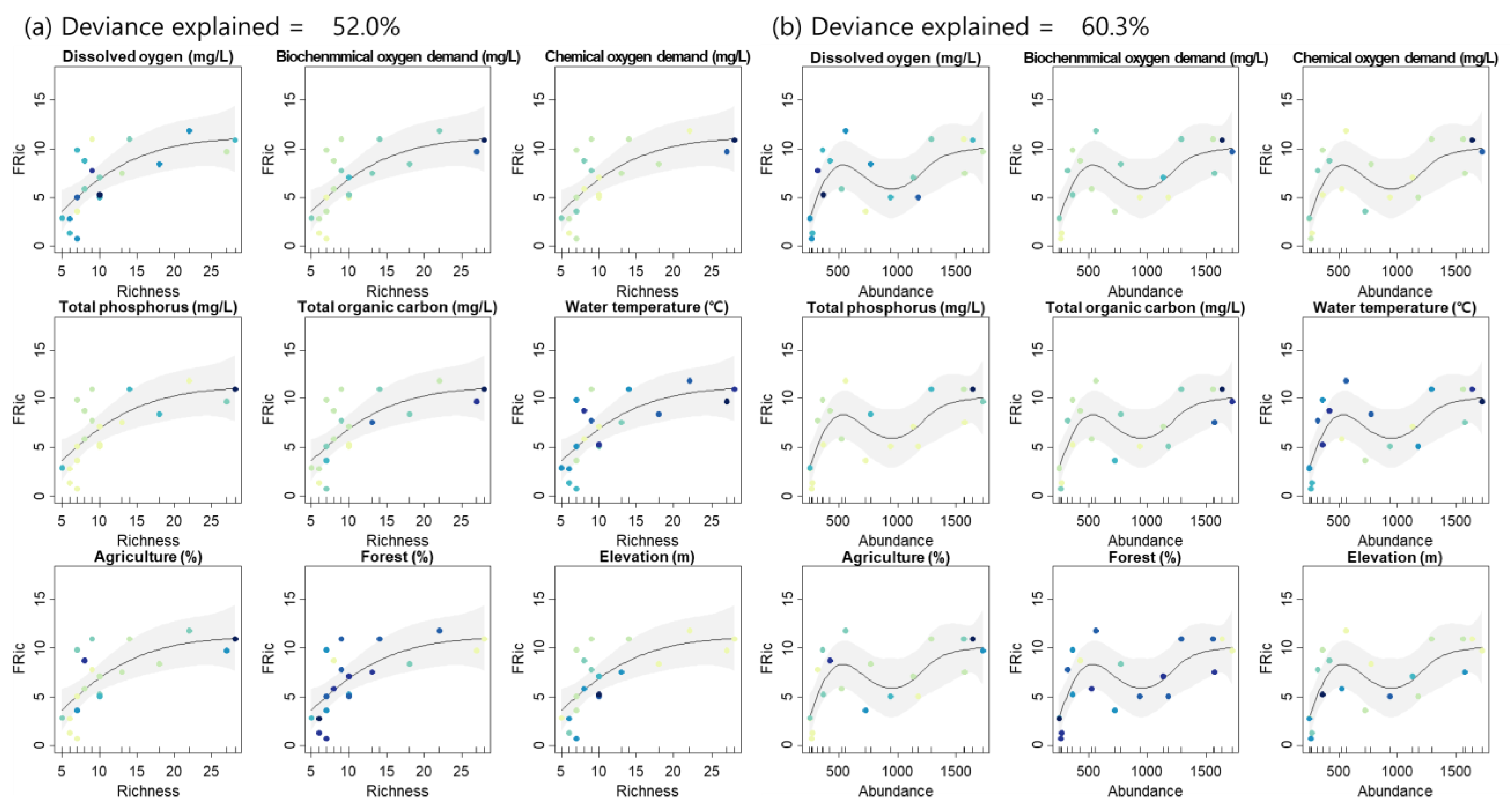
3.3. Relationship between Diversity Indices
4. Discussion
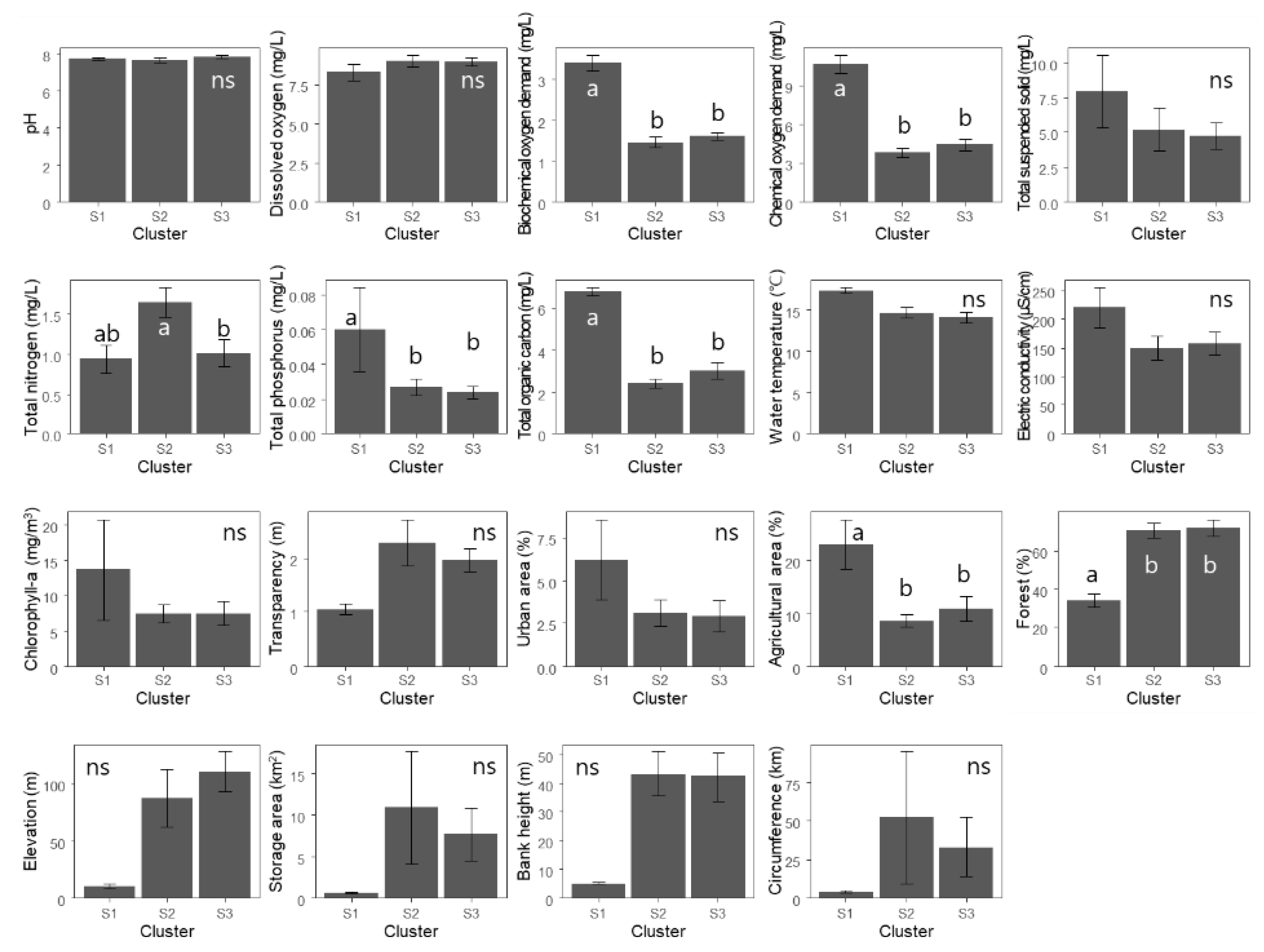
4.1. Taxonomic Community Structure
4.2. Relationships between Functional Traits of Macroinvertebrates
4.3. Taxonomic Diversity and Functional Diversity
4.4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Bae, M.-J.; Li, F.; Kwon, Y.-S.; Chung, N.; Choi, H.; Hwang, S.-J.; Park, Y.-S. Concordance of diatom, macroinvertebrate and fish assemblages in streams at nested spatial scales: Implications for ecological integrity. Ecol. Indic. 2014, 47, 89–101. [Google Scholar] [CrossRef]
- Menezes, S.; Baird, D.J.; Soares, A.M.V.M. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Tilman, D. Functional diversity. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Elsevier: New York, NY, USA, 2001; pp. 109–120. [Google Scholar]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Moon, M.Y.; Ji, C.W.; Lee, D.-S.; Lee, D.-Y.; Hwang, S.-J.; Noh, S.-Y.; Kwak, I.-S.; Park, Y.-S. Characterizing responses of biological trait and functional diversity of benthic macroinvertebrates to environmental variables to develop aquatic ecosystem health assessment index. Korean J. Ecol. Environ. 2020, 53, 31–45. [Google Scholar] [CrossRef]
- Hooper, D.U.; Solan, M.; Symstad, A.; Díaz, S.; Gessener, M.O.; Buchmann, N.; Degrange, V.; Grime, P.; Hulot, F.; Mermillod-Blondin, F.; et al. Species diversity, functional diversity and ecosystem functioning. In Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Cummins, K.W. Trophic relations of aquatic insects. Annu. Rev. Entomol. 1973, 18, 183–206. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Díaz, S.; Purvis, A.; Cornelissen, J.H.C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomanova, S.; Moya, N.; Oberdorff, T. Using macroinvertebrate biological traits for assessing biotic integrity of neotropical streams. River Res. Appl. 2008, 24, 1230–1239. [Google Scholar] [CrossRef]
- Villéger, S.; Miranda, J.R.; Hernández, D.F.; Mouillot, D. Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation. Ecol. Appl. 2010, 20, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Jarzyna, M.A.; Jetz, W. Taxonomic and functional diversity change is scale dependent. Nat. Commun. 2018, 9, 2565. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.-J.; Kwon, Y.; Hwang, S.-J.; Chon, T.-S.; Yang, H.-J.; Kwak, I.-S.; Park, J.-H.; Ham, S.-A.; Park, Y.-S. Relationships between three major stream assemblages and their environmental factors in multiple spatial scales. Ann. De Limnol.-Int. J. Limnol. 2011, 47, S91–S105. [Google Scholar] [CrossRef] [Green Version]
- Muralidharan, M.; Selvakumar, C.; Sudar, S.; Raja, M. Macroinvertebrates as potential indicators of environmental quality. Int. J. Biol. Technol. 2010, 1, 23–28. [Google Scholar]
- Bae, M.-J.; Park, Y.-S. Responses of the functional diversity of benthic macroinvertebrates to floods and droughts in small streams with different flow permanence. Inland Waters 2016, 6, 461–475. [Google Scholar] [CrossRef]
- Bae, M.-J.; Park, Y.-S. Evaluation of precipitation impacts on benthic macroinvertebrate communities at three different stream types. Ecol. Indic. 2019, 102, 446–456. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Species diversity and functional diversity relationship varies with disturbance intensity. Ecosphere 2011, 2, art52. [Google Scholar] [CrossRef]
- Fu, X.; Yang, W.; Zheng, L.; Liu, D.; Li, X. Spatial patterns of macrobenthos taxonomic and functional diversity throughout the ecotones from river to lake: A case study in Northern China. Front. Ecol. Evol. 2022, 10, 922539. [Google Scholar] [CrossRef]
- Coccia, C.; Almeida, B.A.; Green, A.J.; Gutiérrez, A.B.; Carbonell, J.A. Functional diversity of macroinvertebrates as a tool to evaluate wetland restoration. J. Appl. Ecol. 2021, 58, 2999–3011. [Google Scholar] [CrossRef]
- Minister of Environment (MOE); National Institute of Environmental Research (NIER). Survey on the Environment and Ecosystem of Lakes; MOE/NIER: Incheon, Republic of Korea, 2017. [Google Scholar]
- Bae, M.-J.; Park, Y.-S. Diversity and distribution of endemic stream insects on a nationwide scale, South Korea: Conservation perspectives. Water 2017, 9, 833. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-Y.; Lee, D.-S.; Hwang, S.-J.; Lee, K.-L.; Park, Y.-S. Distribution patterns and vulnerability of stoneflies (Plecoptera: Insecta) in South Korean streams with conservation perspectives. Glob. Ecol. Conserv. 2022, 34, e02030. [Google Scholar] [CrossRef]
- Kim, S.-J.; Song, H.-J.; Park, T.-J.; Hwang, M.-Y.; Cho, H.-S.; Song, K.-D.; Lee, H.-J.; Kim, Y.-S. Survey on lake environments in the Yeongsan and Seomjin river basins-Based on 10 lakes such as Hadong and Sangsa. J. Korean Soc. Water Environ. 2015, 31, 665–679. [Google Scholar] [CrossRef] [Green Version]
- QGIS. org. QGIS Geographic Information Systemp; 3.22.09; QGIS Association: Geneva, Switzerland, 2022. [Google Scholar]
- Phillips, N. Stream biomonitoring using species traits. Water Atmos. 2004, 12, 586. [Google Scholar]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- Sarremejane, R.; Cid, N.; Stubbington, R.; Datry, T.; Alp, M.; Cañedo-Argüelles, M.; Cordero-Rivera, A.; Csabai, Z.; Gutiérrez-Cánovas, C.; Heino, J.; et al. DISPERSE, a trait database to assess the dispersal potential of European aquatic macroinvertebrates. Sci. Data 2020, 7, 386. [Google Scholar] [CrossRef]
- Kwon, S.J.; Jun, Y.-C.; Park, J.-H. Benthic Macroinvertebrates; Nature & Ecology: Seoul, Republic of Korea, 2013. [Google Scholar]
- U.S. EPA. Freshwater Biological Traits Database (Final Report); U.S. EPA: Washington, DC, USA, 2012. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessmnet Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, Second Edition; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. [Google Scholar]
- Botsford, L.W.; Holland, M.D.; Samhouri, J.F.; White, J.W.; Hastings, A. Importance of age structure in models of the response of upper trophic levels to fishing and climate change. ICES J. Mar. Sci. 2011, 68, 1270–1283. [Google Scholar] [CrossRef]
- Öckinger, E.; Schweiger, O.; Crist, T.O.; Debinski, D.M.; Krauss, J.; Kuussaari, M.; Petersen, J.D.; Pöyry, J.; Settele, J.; Summerville, K.S.; et al. Life-history traits predict species responses to habitat area and isolation: A cross-continental synthesis. Ecol. Lett. 2010, 13, 969–979. [Google Scholar] [CrossRef]
- García-Barros, E. Body size, egg size, and their interspecific relationships with ecological and life history traits in butterflies (Lepidoptera: Papilionoidea, Hesperioidea). Biol. J. Linn. Soc. 2008, 70, 251–284. [Google Scholar] [CrossRef]
- Swanson, S.K.; Bahr, J.M.; Schwar, M.T.; Potter, K.W. Two-way cluster analysis of geochemical data to constrain spring source waters. Chem. Geol. 2001, 179, 73–91. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Grace, J.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; Volume 28. [Google Scholar]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Vesanto, J. Neural Network Tool for Data Mining: SOM toolbox; TOOLMET 2000-3rd International Symposium on Tool Environments and Development Methods for Intelligent Systems: Oulu, Finland, 2000. [Google Scholar]
- Céréghino, R.; Park, Y.S. Review of the Self-Organizing Map (SOM) approach in water resources: Commentary. Environ. Model. Softw. 2009, 24, 945–947. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Lee, D.-S.; Bae, M.-J.; Hwang, S.-J.; Noh, S.-Y.; Moon, J.-S.; Park, Y.-S. Distribution patterns of odonate assemblages in relation to environmental variables in streams of South Korea. Insects 2018, 9, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahsler, M.; Grün, B.; Hornik, K. arules-A computational environment for mining association rules and frequent item sets. J. Stat. Softw. 2005, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Brin, S.; Motwani, R.; Ullman, J.D.; Tsur, S. Dynamic itemset counting and implication rules for market basket data. In Proceedings of the 1997 ACM SIGMOD International Conference on Management of Data, Tucson, AZ, USA, 13–15 May 1997; Association for Computing Machinery: Tucson, AZ, USA, 1997; pp. 255–264. [Google Scholar]
- Hastie, T.J.; Tibshirani, R.J. Generalized Additive Models; Routledge: Abingdon, UK, 1990. [Google Scholar]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberte, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package. 2014. Available online: https://cran.r-project.org/web/packages/FD/FD.pdf (accessed on 2 May 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 17 April 2022).
- Wehrens, R.; Kruisselbrink, J. Kohonen: Supervised and Unsupervised Self-Organising Maps. R Package. 2018. Available online: https://cran.r-project.org/web/packages/kohonen/kohonen.pdf (accessed on 29 March 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Pohlert, T. PMCMRplus: Calculate Pairwise Multiple Comparisons of Mean Rank Sums Extended. R Package. 2021. Available online: https://cran.r-project.org/web/packages/PMCMRplus/PMCMRplus.pdf (accessed on 10 July 2022).
- Hahsler, M.; Buchta, C.; Gruen, B.; Hornik, K. Arules: Mining Association Rules and Frequent Itemsets, 1.7–4. 2022. Available online: https://cran.r-project.org/web/packages/arules/arules.pdf (accessed on 16 August 2022).
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: New York, NY, USA, 2017. [Google Scholar]
- Wood, S. Mgcv: Mixed GAM Computation Vehicle with Automatic Smoothness Estimation. R Package. 2022. Available online: https://cran.r-project.org/web/packages/mgcv/mgcv.pdf (accessed on 21 October 2022).
- Pangle, K.L.; Malinich, T.D.; Bunnell, D.B.; DeVries, D.R.; Ludsin, S.A. Context-dependent planktivory: Interacting effects of turbidity and predation risk on adaptive foraging. Ecosphere 2012, 3, art114. [Google Scholar] [CrossRef]
- Sweka, J.A.; Hartman, K.J. Influence of turbidity on brook trout reactive distance and foraging success. Trans. Am. Fish. Soc. 2001, 130, 138–146. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, L.; Li, K.; Zhang, L.; Cai, Y.; Wang, X.; Heino, J. Nutrient enrichment homogenizes taxonomic and functional diversity of benthic macroinvertebrate assemblages in shallow lakes. Limnol. Oceanogr. 2019, 64, 1047–1058. [Google Scholar] [CrossRef]
- Yang, Y.; Yi, Y.; Zhou, Y.; Wang, X.; Zhang, S.; Yang, Z. Spatio-temporal variations of benthic macroinvertebrates and the driving environmental variables in a shallow lake. Ecol. Indic. 2020, 110, 105948. [Google Scholar] [CrossRef]
- Rawson, C.A.; Lim, R.P.; Tremblay, L.A.; Warne, M.S.J.; Ying, G.-G.; Laginestra, E.; Chapman, J.C. Benthic macroinvertebrate assemblages in remediated wetlands around Sydney, Australia. Ecotoxicology 2010, 19, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kwon, Y.-S.; Hwang, S.-J.; Park, S. Characterizing effects of landscape and morphometric factors on water quality of reservoirs using a self-organizing map. Environ. Model. Softw. 2014, 55, 214–221. [Google Scholar] [CrossRef]
- Kwon, Y.-S.; Bae, M.J.; Kim, J.-S.; Kim, Y.-J.; Kim, B.-H.; Park, Y.S. Characterizing changes of water quality and relationships with environmental factors in the selected Korean reservoirs. Korean J. Ecol. Environ. 2014, 47, 146–159. [Google Scholar] [CrossRef]
- Blocksom, K.A.; Kurtenbach, J.P.; Klemm, D.J.; Fulk, F.A.; Cormier, S.M. Development and evaluation of the lake macroinvertebrate integrity index (LMII) for New Jersey lakes and reservoirs. Environ. Monit. Assess. 2002, 77, 311–333. [Google Scholar] [CrossRef]
- Spieles, D.J.; Mitsch, W.J. A model of macroinvertebrate trophic structure and oxygen demand in freshwater wetlands. Ecol. Model. 2003, 161, 183–194. [Google Scholar] [CrossRef]
- Lougheed, V.L.; Crosbie, B.; Chow-Fraser, P. Primary determinants of macrophyte community structure in 62 marshes across the Great Lakes basin: Latitude, land use, and water quality effects. Can. J. Fish. Aquat. Sci. 2001, 58, 1603–1612. [Google Scholar] [CrossRef]
- Usseglio-Polatera, P.; Bournaud, M.; Richoux, P.; Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Firmiano, K.R.; Castro, D.M.P.; Linares, M.S.; Callisto, M. Functional responses of aquatic invertebrates to anthropogenic stressors in riparian zones of Neotropical savanna streams. Sci. Total Environ. 2021, 753, 141865. [Google Scholar] [CrossRef]
- Pianka, E.R. On r- and K-Selection. Am. Nat. 1970, 104, 592–597. [Google Scholar] [CrossRef]
- Zeuss, D.; Brunzel, S.; Brandl, R. Environmental drivers of voltinism and body size in insect assemblages across Europe. Glob. Ecol. Biogeogr. 2017, 26, 154–165. [Google Scholar] [CrossRef]
- Yang, F.; Kawabata, E.; Tufail, M.; Brown, J.J.; Takeda, M. r/K-like trade-off and voltinism discreteness: The implication to allochronic speciation in the fall webworm, Hyphantria cunea complex (Arctiidae). Ecol. Evol. 2017, 7, 10592–10603. [Google Scholar] [CrossRef] [PubMed]
- Keppeler, F.W.; Montaña, C.G.; Winemiller, K.O. The relationship between trophic level and body size in fishes depends on functional traits. Ecol. Monogr. 2020, 90, e01415. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Gergs, A.; Ratte, H.T. Predicting functional response and size selectivity of juvenile Notonecta maculata foraging on Daphnia magna. Ecol. Model. 2009, 220, 3331–3341. [Google Scholar] [CrossRef]
- Pawar, S.; Dell, A.I.; Van, M.S. Dimensionality of consumer search space drives trophic interaction strengths. Nature 2012, 486, 485–489. [Google Scholar] [CrossRef]
- Anderson, N.H.; Cummins, K.W. Influences of diet on the life histories of aquatic insects. J. Fish. Res. Board Can. 1979, 36, 335–342. [Google Scholar] [CrossRef]
- Wissinger, S.A. Life history and size structure of larval dragonfly populations. J. N. Am. Benthol. Soc. 1988, 7, 13–28. [Google Scholar] [CrossRef]
- Teder, T.; Tammaru, T.; Esperk, T. Dependence of phenotypic variance in body size on environmental quality. Am. Nat. 2008, 172, 223–232. [Google Scholar] [CrossRef] [Green Version]
- García-Navas, V.; Sattler, T.; Schmid, H.; Ozgul, A. High elevation bird communities in the Swiss Alps exhibit reduced fecundity and lifespan independently of phylogenetic effects. Biodivers. Conserv. 2021, 30, 991–1010. [Google Scholar] [CrossRef]
- Collins, S.L.; Avolio, M.L.; Gries, C.; Hallett, L.M.; Koerner, S.E.; La Pierre, K.J.; Rypel, A.L.; Sokol, E.R.; Fey, S.B.; Flynn, D.F.B.; et al. Temporal heterogeneity increases with spatial heterogeneity in ecological communities. Ecology 2018, 99, 858–865. [Google Scholar] [CrossRef]
- Lytle, D.A. Disturbance regimes and life-history evolution. Am. Nat. 2001, 157, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Reynaga, M.C.; Dos Santos, D.A. Contrasting taxonomical and functional responses of stream invertebrates across space and time in a Neotropical basin. Fundam. Appl. Limnol. 2013, 183, 121–133. [Google Scholar] [CrossRef]
- Hacala, A.; Lafage, D.; Prinzing, A.; Sawtschuk, J.; Pétillon, J. Drivers of taxonomic, functional and phylogenetic diversities in dominant ground-dwelling arthropods of coastal heathlands. Oecologia 2021, 197, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Schmera, D.; Heino, J.; Podani, J. Characterising functional strategies and trait space of freshwater macroinvertebrates. Sci. Rep. 2022, 12, 12283. [Google Scholar] [CrossRef] [PubMed]
- Barnum, T.R.; Weller, D.E.; Williams, M. Urbanization reduces and homogenizes trait diversity in stream macroinvertebrate communities. Ecol. Appl. 2017, 27, 2428–2442. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, X.; Wang, J.; Meng, X.; Heino, J.; Xie, Z. Multiple facets of stream macroinvertebrate alpha diversity are driven by different ecological factors across an extensive altitudinal gradient. Ecol. Evol. 2019, 9, 1306–1322. [Google Scholar] [CrossRef] [PubMed]
- Heino, J. Functional biodiversity of macroinvertebrate assemblages along major ecological gradients of boreal headwater streams. Freshw. Biol. 2005, 50, 1578–1587. [Google Scholar] [CrossRef]
- Luck, G.W.; Carter, A.; Smallbone, L. Changes in bird functional diversity across multiple land uses: Interpretations of functional redundancy depend on functional group identity. PLoS ONE 2013, 8, e63671. [Google Scholar] [CrossRef] [Green Version]
- Klemm, D.J.; Lewis, P.A.; Fulk, F.; Lazorchak, J.M. Macroinvertebrate Field and Laboratory Methods for Evaluating the Biological Integrity of Surface Waters; US EPA: Washington, DC, USA, 1990. [Google Scholar]
- Hails, J.R. Grap Samplers; Springer US: Boston, MA, USA, 1982. [Google Scholar]
- de Mendoza, G.; Catalan, J. Lake macroinvertebrates and the altitudinal environmental gradient in the Pyrenees. Hydrobiologia 2010, 648, 51–72. [Google Scholar] [CrossRef]
- Moore, I.E.; Murphy, K.J. Evaluation of alternative macroinvertebrate sampling techniques for use in a new tropical freshwater bioassessment scheme. Acta Limnol. Bras. 2015, 27, 213–222. [Google Scholar] [CrossRef]
- Li, Z.; Heino, J.; Liu, Z.; Meng, X.; Chen, X.; Ge, Y.; Xie, Z. The drivers of multiple dimensions of stream macroinvertebrate beta diversity across a large montane landscape. Limnol. Oceanogr. 2021, 66, 226–236. [Google Scholar] [CrossRef]
- Vieira, N.M.K.; Poff, N.L.; Carlisle, D.; Moulton Ii, S.R.; Koski, M.L.; Kondratieff, B.C. A Database of Lotic Invertebrate Traits for North America; Data Series 187; 2006; 19p. Available online: https://pubs.er.usgs.gov/publication/ds187 (accessed on 7 December 2022).
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’eau Douce Systématique, Biologie, Écologie; CNRS Editions: Paris, France, 2000. [Google Scholar]
- Rodhain, F. Ecology of Aedes aegypti in Africa and Asia. Bull. De La Soc. De Pathol. Exot. 1996, 89, 103–106. [Google Scholar]
- Koch, M. Observations of the reproduction and population structure of the caenogastropod, Gabbia vertiginosa Frauenfeld, 1862 (Rissooidea: Bithyniidae). Molluscan Res. 2004, 24, 65–73. [Google Scholar] [CrossRef]
- Villalobos, M.C.; Monge-Nájera, J.; Barrientos, Z.; Franco, J. Life cycle and field abundance of the snail Succinea costaricana (Stylommatophora: Succineidae), a tropical agricultural pest. Rev. De Biol. Trop. 2015, 43, 181–188. [Google Scholar]
- Kuznik-Kowalska, E.; Pokryszko, B.M.; Prockow, M.; Oczkowska, M. On the population dynamics, reproductive biology and growth of Succinea putris (Linnaeus 1758)(Gastropoda: Pulmonata: Succineidae). Folia Malacol. 2013, 21, 215–224. [Google Scholar] [CrossRef]
- Jeliazkov, A. Scale-Effects in Agriculture-Environment-Biodiversity Relationships; Université Pierre et Marie Curie: Paris, France, 2013. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Trait State (Modality) | Code | Abbreviation |
|---|---|---|---|
| Voltinism | Semivoltine (<1 generation/y) | Volt1 | V1 |
| Univoltine (1 generation/y) | Volt2 | V2 | |
| Bi- or multivoltine (>1 generation/y) | Volt3 | V3 | |
| Adult life span | Very short (<1 wk) | Life1 | A1 |
| Short (<1 mo) | Life2 | A2 | |
| Long (>1 mo) | Life3 | A3 | |
| Adult size | Small (<9 mm) | Size1 | S1 |
| Medium (9–16 mm) | Size2 | S2 | |
| Large (>16 mm) | Size3 | S3 | |
| Tropic habit | Filterer-collector | FFG1 | F1 |
| Gatherer-collector | FFG2 | F2 | |
| Herbivore (scraper, piercer, and shedder) | FFG3 | F3 | |
| Predator (piercer and engulfer) | FFG4 | F4 | |
| Shredder (detritivore) | FFG5 | F5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-Y.; Lee, D.-S.; Park, Y.-S. Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea. Int. J. Environ. Res. Public Health 2023, 20, 673. https://doi.org/10.3390/ijerph20010673
Lee D-Y, Lee D-S, Park Y-S. Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea. International Journal of Environmental Research and Public Health. 2023; 20(1):673. https://doi.org/10.3390/ijerph20010673
Chicago/Turabian StyleLee, Da-Yeong, Dae-Seong Lee, and Young-Seuk Park. 2023. "Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea" International Journal of Environmental Research and Public Health 20, no. 1: 673. https://doi.org/10.3390/ijerph20010673
APA StyleLee, D. -Y., Lee, D. -S., & Park, Y. -S. (2023). Taxonomic and Functional Diversity of Benthic Macroinvertebrate Assemblages in Reservoirs of South Korea. International Journal of Environmental Research and Public Health, 20(1), 673. https://doi.org/10.3390/ijerph20010673