
The Influence of Hip and Knee Joint Angles on Quadriceps Muscle-Tendon Unit Properties during Maximal Voluntary Isometric Contraction

 , , ,
, , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Trial Design
2.2. Participants
2.3. Randomization, Allocation Concealment, and Blinding
2.4. Experimental Procedures
2.5. Outcomes
2.5.1. Torque Evaluation
2.5.2. EMG
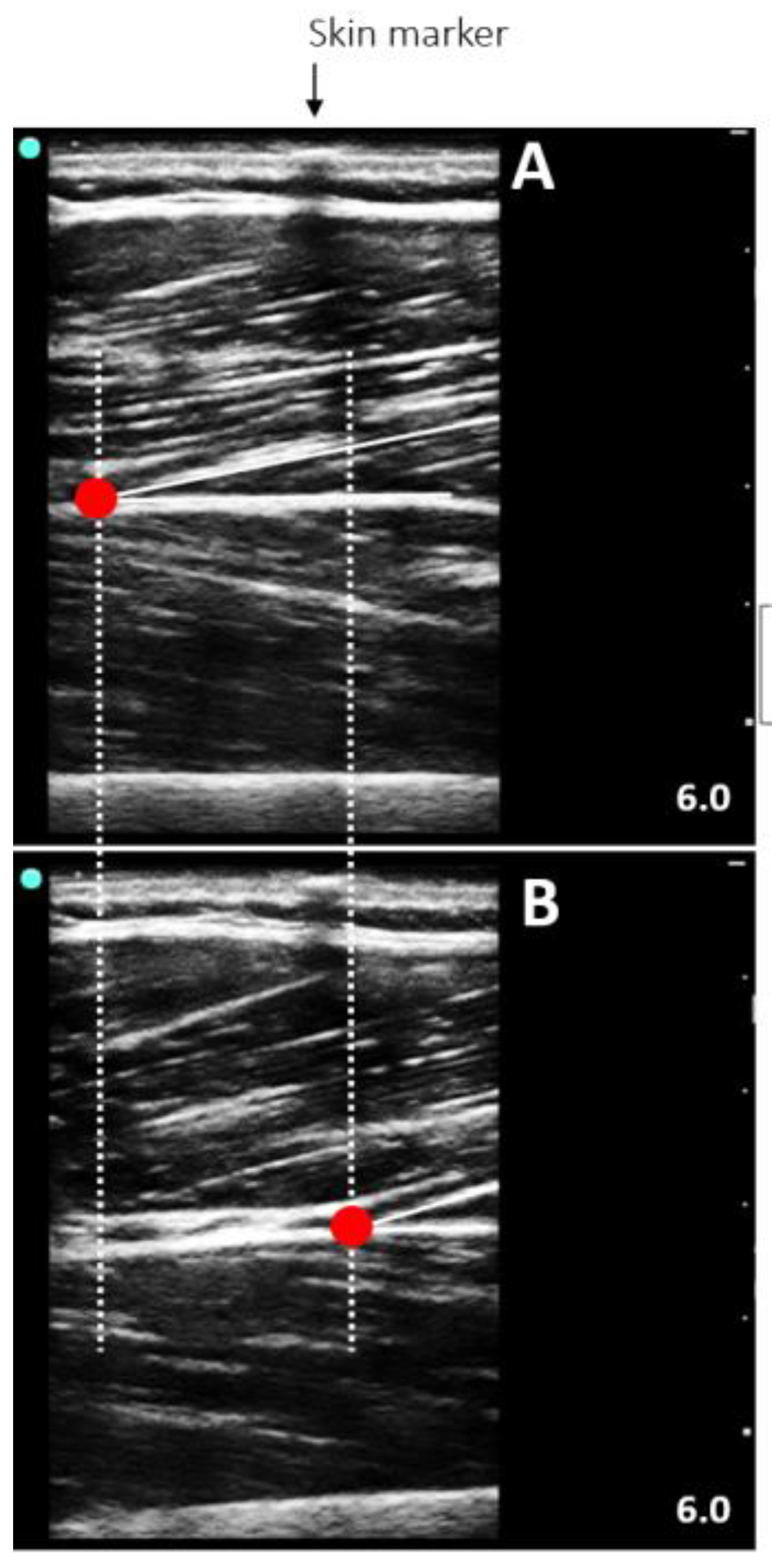
2.5.3. Muscle Architecture Assessment
2.5.4. TAC Stiffness
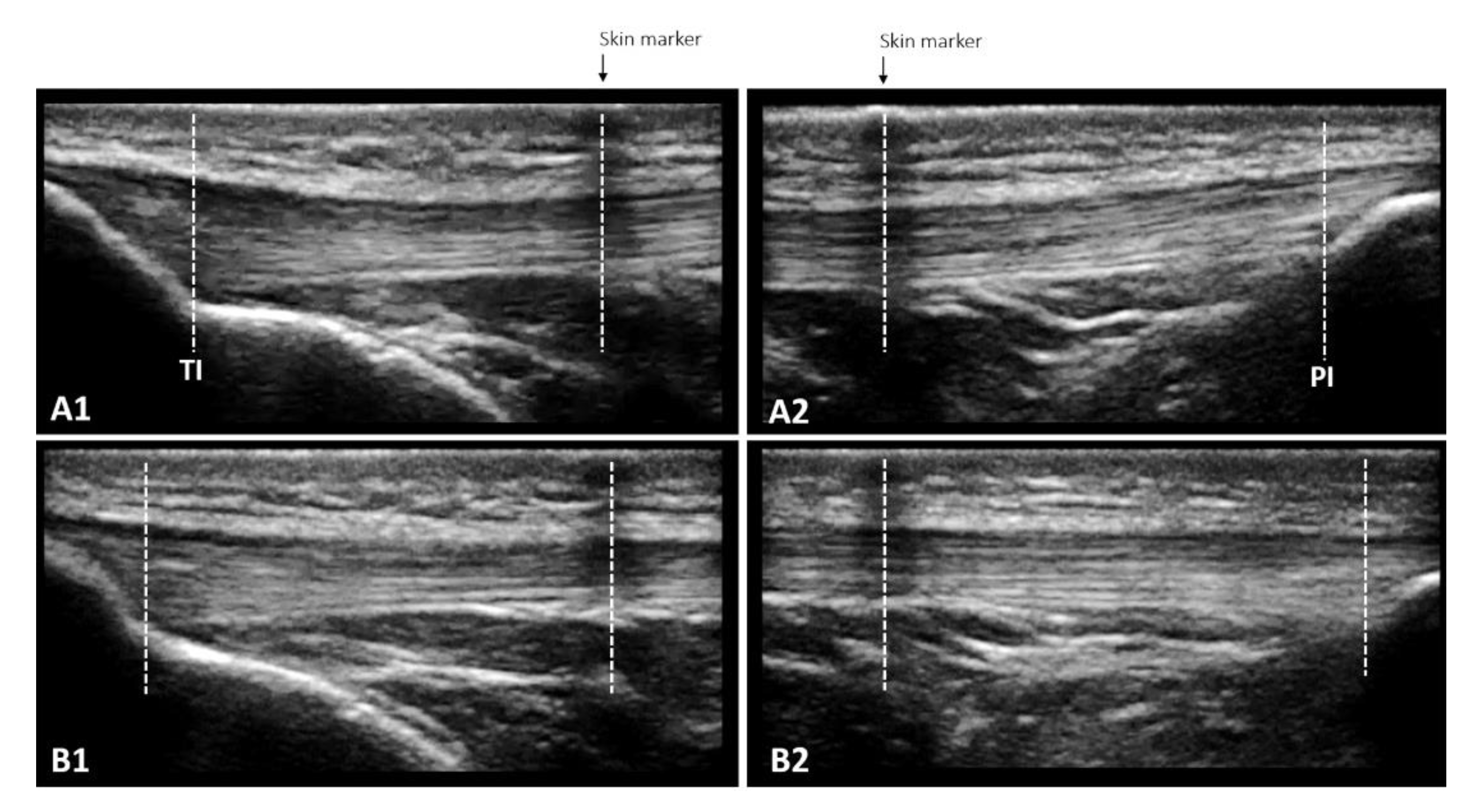
2.5.5. Patellar Tendon Properties
Morphological Properties
Mechanical Properties
Material Properties
2.6. Statistical Analysis and Sample Size
3. Results
3.1. Reliability of Measurements
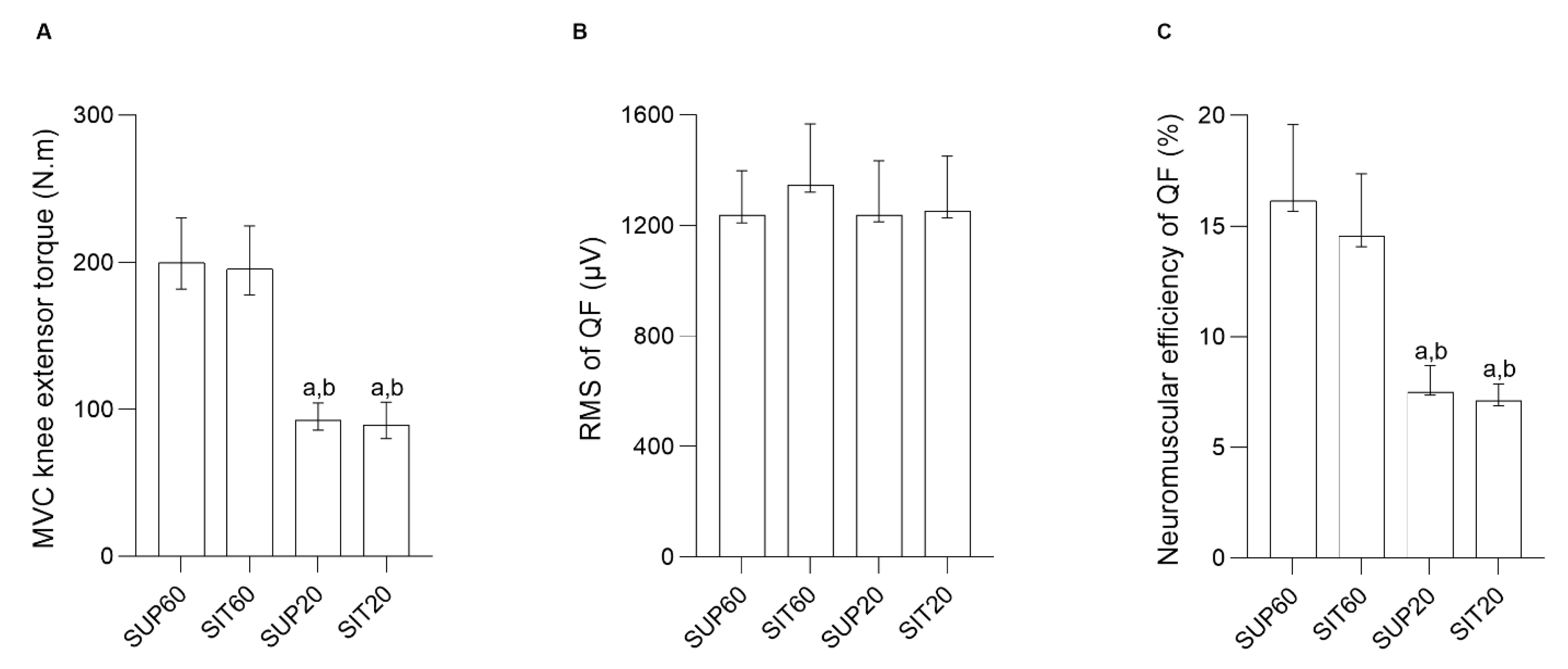
3.2. MVIC, Raw RMS, and Quadriceps Femoris Neuromuscular Efficiency
3.3. TAC Stiffness
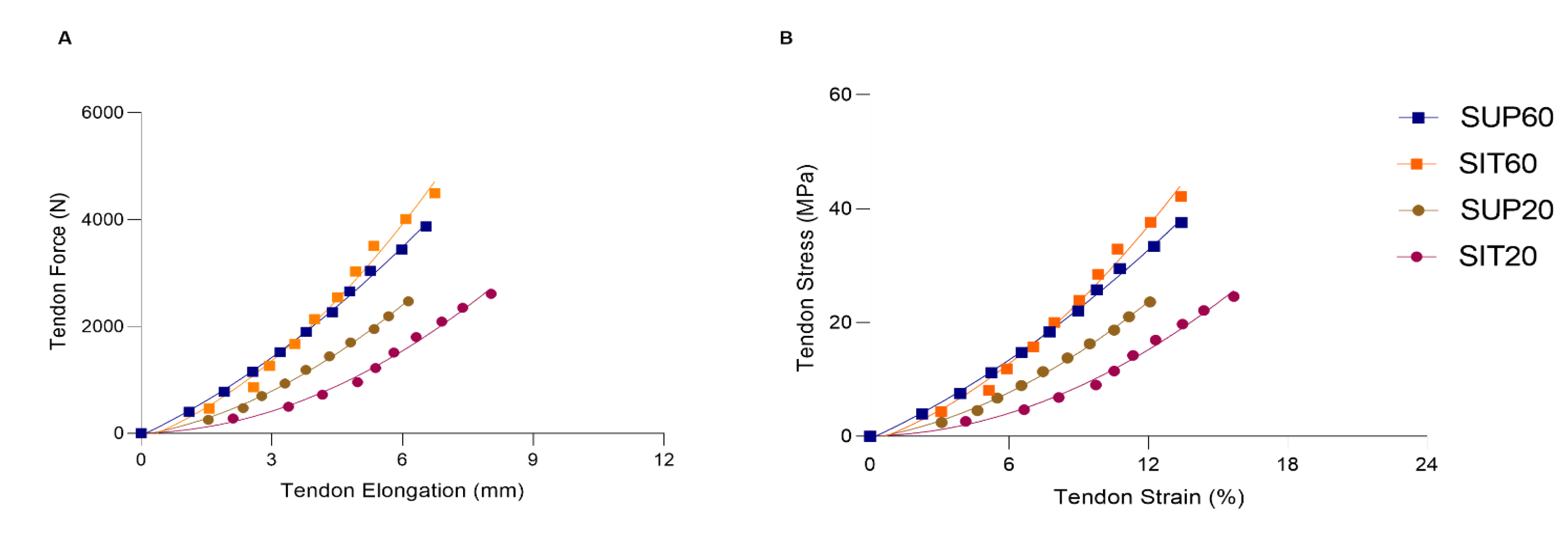
3.4. Tendon Properties
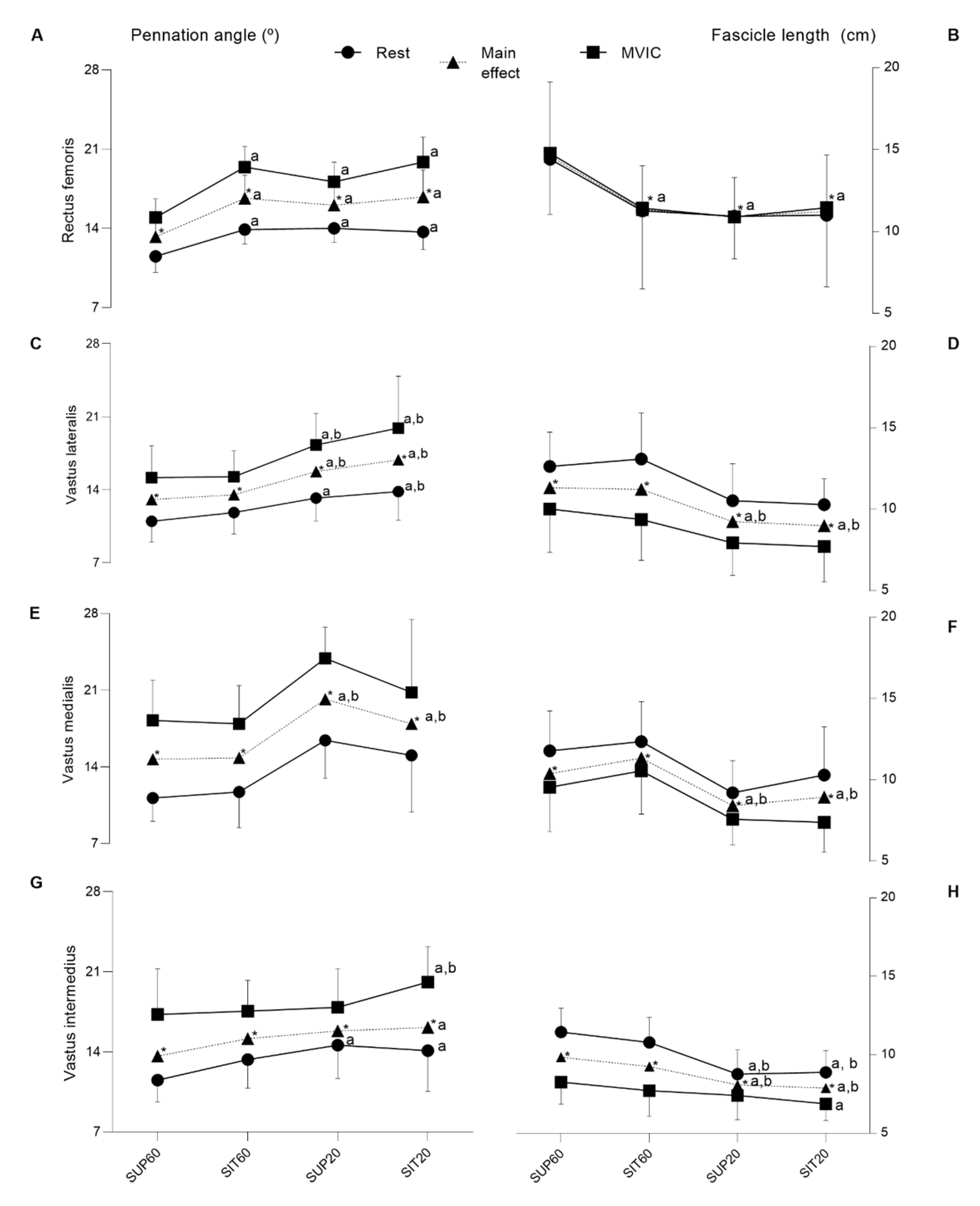
3.4.1. Morphological Properties
3.4.2. Mechanical Properties
3.4.3. Material Properties
4. Discussion
4.1. MVIC, RMS, and Quadriceps Neuromuscular Efficiency
4.2. Muscle Architecture
4.3. TAC Stiffness
4.4. Patellar Tendon Properties
4.4.1. Morphological Properties
4.4.2. Mechanical and Material Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MVIC | Maximal voluntary isometric contraction |
| RF | Rectus femoris |
| VM | Vastus medialis |
| VL | Vastus lateralis |
| VI | Vastus intermedius |
| Lf | fascicle length |
| Θp | pennation angle |
| TAC | Tendon aponeurosis complex |
| EMG | surface electromyography |
| CONSORT | Consolidated Standards of Reporting Trials |
| SUP60 | supine with 60° of knee flexion |
| SIT60 | seated with 60° of knee flexion |
| SUP20 | supine with 20° of knee flexion |
| SIT20 | seated with 20° of knee flexion |
| RMS | Root mean square |
| CI | Confidence interval 95% |
| ANOVA | Analysis of variance |
| ηρ2 | eta squared |
| ICC | Intra-class correlation |
References
- Vincent, K.R.; Vincent, H.K. Resistance exercise for knee osteoarthritis. PM R 2012, 4, S45–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, A. Current issues in arthrogenous inhibition. Ann Rheum Dis. 1993, 52, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, D.A.; McNair, P.J. Quadriceps arthrogenic muscle inhibition: Neural mechanisms and treatment perspectives. Semin. Arthritis Rheum. 2010, 40, 250–266. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, G.K.; Piva, S.R.; Irrgang, J.J. A modified neuromuscular electrical stimulation protocol for quadriceps strength training following anterior cruciate ligament reconstruction. J. Orthop. Sports Phys. Ther. 2003, 33, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Grob, K.; Manestar, M.; Gascho, D.; Ackland, T.; Gilbey, H.; Fretz, C.; Kuster, M.S. Magnetic resonance imaging of the tensor vastus intermedius: A topographic study based on anatomical dissections. Clin. Anat. 2017, 30, 1096–1102. [Google Scholar] [CrossRef]
- Herzog, W.; Abrahamse, S.K.; ter Keurs, H.E.D.J. Theoretical determination of force-length relations of intact human skeletal muscles using the cross-bridge model. Pflugers Arch. 1990, 416, 113–119. [Google Scholar] [CrossRef]
- Pincivero, D.M.; Salfetnikov, Y.; Campy, R.M.; Coelho, A.J. Angle- and gender-specific quadriceps femoris muscle recruitment and knee extensor torque. J. Biomech. 2004, 37, 1689–1697. [Google Scholar] [CrossRef]
- Lanza, M.B.; Balshaw, T.G.; Folland, J.P. Do changes in neuromuscular activation contribute to the knee extensor angle-torque relationship? Exp. Physiol. 2017, 102, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, J.G.T.; Marqueti, R.D.C.; Geremia, J.M.; Neto, I.V.D.S.; Baroni, B.M.; Silbernagel, K.G.; Bottaro, M.; Babault, N.; Durigan, J.L.Q. The Effect of Quadriceps Muscle Length on Maximum Neuromuscular Electrical Stimulation Evoked Contraction, Muscle Architecture, and Tendon-Aponeurosis Stiffness. Front. Physiol. 2021, 12, 633589. [Google Scholar] [CrossRef]
- Garnier, Y.M.; Lepers, R.; Canepa, P.; Martin, A.; Paizis, C. Effect of the Knee and Hip Angles on Knee Extensor Torque: Neural, Architectural, and Mechanical Considerations. Front. Physiol. 2022, 12, 789867. [Google Scholar] [CrossRef]
- Maffiuletti, N.A.; Lepers, R. Quadriceps femoris torque and EMG activity in seated versus supine position. Med. Sci. Sports Exerc. 2003, 35, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Ema, R.; Wakahara, T.; Kawakami, Y. Effect of hip joint angle on concentric knee extension torque. J. Electromyogr. Kinesiol. 2017, 37, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, R.F. Knee biomechanics of the dynamic squat exercise. Med. Sci. Sports Exerc. 2001, 33, 127–141. [Google Scholar] [CrossRef]
- Gupton, M.; Imonugo, O.; Terreberry, R.R. Anatomy, Bony Pelvis and Lower Limb, Knee. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Folland, J.P.; Williams, A.G. The adaptations to strength training: Morphological and neurological contributions to increased strength. Sports Med. 2007, 37, 145–168. [Google Scholar] [CrossRef]
- Babault, N.; Pousson, M.; Michaut, A.; Van Hoecke, J. Effect of quadriceps femoris muscle length on neural activation during isometric and concentric contractions. J. Appl. Physiol. 2003, 94, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Fukunaga, T.; Kawakami, Y.; Kuno, S.; Funato, K.; Fukashiro, S. Muscle architecture and function in humans. J. Biomech. 1997, 30, 457–463. [Google Scholar] [CrossRef]
- Wiesinger, H.-P.; Seynnes, O.R.; Kösters, A.; Müller, E.; Rieder, F. Mechanical and Material Tendon Properties in Patients With Proximal Patellar Tendinopathy. Front. Physiol. 2020, 11, 704. [Google Scholar] [CrossRef] [PubMed]
- Suydam, S.M.; Buchanan, T.S.; Manal, K.; Silbernagel, K.G. Compensatory muscle activation caused by tendon lengthening post-Achilles tendon rupture. Knee Surg. Sports Traumatol. Arthrosc. 2015, 23, 868–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, K.; Ikebukuro, T.; Maki, A.; Yata, H.; Tsunoda, N. Time course of changes in the human Achilles tendon properties and metabolism during training and detraining in vivo. Eur. J. Appl. Physiol. 2012, 112, 2679–2691. [Google Scholar] [CrossRef]
- Guzzoni, V.; Ribeiro, M.B.T.; Lopes, G.N.; Marqueti, R.D.C.; de Andrade, R.V.; Selistre-De-Araujo, H.S.; Durigan, J.L.Q. Effect of Resistance Training on Extracellular Matrix Adaptations in Skeletal Muscle of Older Rats. Front. Physiol. 2018, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Kubo, K.; Ohgo, K.; Takeishi, R.; Yoshinaga, K.; Tsunoda, N.; Kanehisa, H.; Fukunaga, T. Effects of isometric training at different knee angles on the muscle-tendon complex in vivo. Scand. J. Med. Sci. Sports 2006, 16, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.E.; Graham-Smith, P.; Pearson, S.J. Effect of acute tensile loading on gender-specific tendon structural and mechanical properties. J. Orthop. Res. 2009, 27, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Fukutani, A.; Misaki, J.; Isaka, T. Influence of Joint Angle on Residual Force Enhancement in Human Plantar Flexors. Front. Physiol. 2017, 8, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, G.J.; Balshaw, T.G.; Maden-Wilkinson, T.M.; Tillin, N.A.; Folland, J.P. Tendinous Tissue Adaptation to Explosive- vs. Sustained-Contraction Strength Training. Front. Physiol. 2018, 9, 1170. [Google Scholar] [CrossRef] [PubMed]
- Abellaneda, S.; Guissard, N.; Duchateau, J. The relative lengthening of the myotendinous structures in the medial gastrocnemius during passive stretching differs among individuals. J. Appl. Physiol. 2009, 106, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Raiteri, B.J. Aponeurosis behaviour during muscular contraction: A narrative review. Eur. J. Sport Sci. 2018, 18, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.F.; Altman, D.G.; Moher, D. CONSORT 2010 statement: Updated guidelines for reporting parallel group randomised trials. J. Pharmacol. Pharmacother. 2010, 1, 100–107. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.J. The Use of Surface Electromyography in Biomechanics. J. Appl. Biomech. J. Appl. Biomech. 1997, 13, 135–163. [Google Scholar] [CrossRef] [Green Version]
- Babault, N.; Desbrosses, K.; Fabre, M.S.; Michaut, A.; Pousson, M. Neuromuscular fatigue development during maximal concentric and isometric knee extensions. J. Appl. Physiol. 2006, 100, 780–785. [Google Scholar] [CrossRef]
- Remaud, A.; Cornu, C.; Guevel, A. A Methodologic Approach for the Comparison Between Dynamic Contractions: Influences on the Neuromuscular System. J. Athl. Train. 2005, 40, 281. [Google Scholar]
- Shahrjerdi, S.; Bahrpeyma, F.; Savelberg, H.H.C.M. Mohajeri-Tehrani, M.R.; Effect of a 6-week strength-training program on neuromuscular efficiency in type 2 diabetes mellitus patients. Diabetol. Int. 2020, 11, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Massey, G.; Evangelidis, P.; Folland, J. Influence of contractile force on the architecture and morphology of the quadriceps femoris. Exp. Physiol. 2015, 100, 1342–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroni, B.M.; Geremia, J.M.; Rodrigues, R.; De Azevedo Franke, R.; Karamanidis, K.; Vaz, M.A. Muscle architecture adaptations to knee extensor eccentric training: Rectus femoris vs. vastus lateralis. Muscle Nerve 2013, 48, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Geremia, J.M.; Baroni, B.M.; Bini, R.R.; Lanferdini, F.J.; de Lima, A.R.; Herzog, W.; Vaz, M.A. Triceps Surae Muscle Architecture Adaptations to Eccentric Training. Front. Physiol. 2019, 10, 1456. [Google Scholar] [CrossRef] [Green Version]
- Finni, T.; Komi, P.V. Two methods for estimating tendinous tissue elongation during human movement. J. Appl. Biomech. 2002, 18, 180–188. [Google Scholar] [CrossRef]
- Bojsen-Moøller, J.; Hansen, P.; Aagaard, P.; Kjær, M.; Magnusson, S.P. Measuring mechanical properties of the vastus lateralis tendon-aponeurosis complex in vivo by ultrasound imaging. Scand. J. Med. Sci. Sport. 2003, 13, 259–265. [Google Scholar] [CrossRef]
- Krevolin, J.L.; Pandy, M.G.; Pearce, J.C. Moment arm of the patellar tendon in the human knee. J. Biomech. 2004, 37, 785–788. [Google Scholar] [CrossRef]
- Seynnes, O.R.; Bojsen-Møller, J.; Albracht, K.; Arndt, A.; Cronin, N.J.; Finni, T.; Magnusson, S.P. Ultrasound-based testing of tendon mechanical properties: A critical evaluation. J. Appl. Physiol. 2015, 118, 133–141. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, T.D.; Reeves, N.D.; Baltzopoulos, V.; Jones, D.A.; Maganaris, C.N. Mechanical properties of the patellar tendon in adults and children. J. Biomech. 2010, 43, 1190–1195. [Google Scholar] [CrossRef]
- Urlando, A.; Hawkins, D. Achilles tendon adaptation during strength training in young adults. Med. Sci. Sports Exerc. 2007, 39, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, J.G.T.; Marqueti, R.D.C.; Corrigan, P.; Sousa, A.M.M.; Bottaro, M.; Babault, N.; Geremia, J.M.; Silbernagel, K.G.; Durigan, J.L.Q. The effects of knee and hip joint angles on patellar tendon loading during quadriceps neuromuscular electrical stimulation. Transl. Sport. Med. 2021, 4, 587–596. [Google Scholar] [CrossRef]
- Kongsgaard, M.; Reitelseder, S.; Pedersen, T.G.; Holm, L.; Aagaard, P.; Kjaer, M.; Magnusson, S.P. Region specific patellar tendon hypertrophy in humans following resistance training. Acta Physiol. 2007, 191, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Geremia, J.M.; Baroni, B.M.; Bobbert, M.F.; Bini, R.R.; Lanferdini, F.J.; Vaz, M.A. Effects of high loading by eccentric triceps surae training on Achilles tendon properties in humans. Eur. J. Appl. Physiol. 2018, 118, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed; Lawrence Erbaum Associates: New York, NY, USA, 1988. [Google Scholar]
- Bampouras, T.M.; Reeves, N.D.; Baltzopoulos, V.; Maganaris, C.N. The role of agonist and antagonist muscles in explaining isometric knee extension torque variation with hip joint angle. Eur. J. Appl. Physiol. 2017, 117, 2039–2045. [Google Scholar] [CrossRef] [Green Version]
- Newman, S.A.; Jones, G.; Newham, D.J. Quadriceps voluntary activation at different joint angles measured by two stimulation techniques. Eur. J. Appl. Physiol. 2003, 89, 496–499. [Google Scholar] [CrossRef]
- Kooistra, R.D.; de Ruiter, C.J.; de Haan, A. Conventionally assessed voluntary activation does not represent relative voluntary torque production. Eur. J. Appl. Physiol. 2007, 100, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Seiberl, W.; Hahn, D.; Paternoster, F.K. Reduced Activation in Isometric Muscle Action after Lengthening Contractions Is Not Accompanied by Reduced Performance Fatigability. Sci Rep 2016, 6. [Google Scholar] [CrossRef]
- Aragão, F.A.; Schäfer, G.S.; de Albuquerque, C.E.; Vituri, R.F.; de Azevedo, F.M.; Bertolini, G.R.F. Neuromuscular Efficiency of the Vastus Lateralis and Biceps Femoris Muscles in Individuals with Anterior Cruciate Ligament Injuries. Rev. Bras. Ortop. Engl. Ed. 2015, 50, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, P.; Andersen, J.L.; Dyhre-Poulsen, P.; Leffers, A.-M.; Wagner, A.; Magnusson, S.P.; Halkjaer-Kristensen, J.; Simonsen, E.B. A mechanism for increased contractile strength of human pennate muscle in response to strength training: Changes in muscle architecture. J. Physiol. 2001, 534, 613–623. [Google Scholar] [CrossRef]
- Blazevich, A.J.; Gill, N.D.; Zhou, S. Intra- and intermuscular variation in human quadriceps femoris architecture assessed in vivo. J. Anat. 2006, 209, 289–310. [Google Scholar] [CrossRef]
- Oliveira, P.; Modesto, K.A.G.; Bottaro, M.; Babault, N.; Durigan, J.L.Q. Training Effects of Alternated and Pulsed Currents on the Quadriceps Muscles of Athletes. Int. J. Sports Med. 2018, 39, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.E.; Smith, C.B.; Harwood, B.; Rice, C.L. In vivo measurement of fascicle length and pennation of the human anconeus muscle at several elbow joint angles. J. Anat. 2014, 225, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.M.; Huxley, A.F.; Julian, F.J. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. J. Physiol. 1966, 184, 170–192. [Google Scholar] [CrossRef] [PubMed]
- Lazarczuk, S.L.; Maniar, N.; Opar, D.A.; Duhig, S.J.; Shield, A.; Barrett, R.S.; Bourne, M.N. Mechanical, Material and Morphological Adaptations of Healthy Lower Limb Tendons to Mechanical Loading: A Systematic Review and Meta-Analysis. Sports Med. 2022, 52, 2405. [Google Scholar] [CrossRef] [PubMed]
- DeFrate, L.E.; Nha, K.W.; Papannagari, R.; Moses, J.M.; Gill, T.J.; Li, G. The biomechanical function of the patellar tendon during in-vivo weight-bearing flexion. J. Biomech. 2007, 40, 1716–1722. [Google Scholar] [CrossRef] [Green Version]
- Edama, M.; Takeishi, M.; Kurata, S.; Kikumoto, T.; Takabayashi, T.; Hirabayashi, R.; Inai, T.; Ikezu, M.; Kaneko, F.; Kageyama, I. Morphological features of the inferior fascicle of the anterior inferior tibiofibular ligament. Sci. Rep. 2019, 9, 10472. [Google Scholar] [CrossRef] [Green Version]
- Depalle, B.; Qin, Z.; Shefelbine, S.J.; Buehler, M.J. Influence of cross-link structure, density and mechanical properties in the mesoscale deformation mechanisms of collagen fibrils. J Mech Behav Biomed Mater. 2015, 52, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zuskov, A.; Freedman, B.R.; Gordon, J.A.; Sarver, J.J.; Buckley, M.R.; Soslowsky, L.J. Tendon biomechanics and crimp properties following fatigue loading are influenced by tendon type and age in mice. J. Orthop. Res. 2020, 38, 36. [Google Scholar] [CrossRef]
- Bohm, S.; Mersmann, F.; Arampatzis, A. Human tendon adaptation in response to mechanical loading: A systematic review and meta-analysis of exercise intervention studies on healthy adults. Sport. Med.—Open 2015, 1, 7. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SUP60 | SIT60 | SUP20 | SIT20 | |
|---|---|---|---|---|
| Morphological properties | ||||
| Resting length (mm) | 48.95 (46.79–51.53) | 50.45 (47.92–53.64) | 50,62 (48.40–53.29) | 50,55 (47.94–53.86) |
| CSA (mm2) | 103.33 (97.63–110.45) | 107.18 (102.23–113.16) | 104.03 (98.20–111.14) | 107.12 (103.98–110.68) |
| Mechanical properties | ||||
| Force (N) | 3768.9 (3391.6–4353.1) | 4341.37 (3913.85–5070.55) | 2379.82 (2134.06–2807.60) a,b | 2497.16 (2193.69–3025.21) a,b |
| Elongation (mm) | 6.19 (5.44–7.64) | 6.25 (5.49–7.98) | 5.72 (5.07–7.20) | 7.51 (6.53–9.54) c |
| Stiffness (N/mm) | 771.75 (636.78–1112.95) | 1008.33 (822.89–1674.57) | 600.05 (511.31–816.29) b | 579.92 (492.24–790.34) b |
| TAC Stiffness QF (N/mm) | 174.17 (169.18–189.15) | 176.22 (171.19–195.83) | 158.77 (154.35–179.66) | 114.00 (111.28–130.51) a,b,c |
| Material properties | ||||
| Stress (MPa) | 36.48 (32.61–42.42) | 40.51 (36.32–47.92) | 22.88 (20.65–26.62) a,b | 23.43 (20.61–28.53) a,b |
| Strain (%) | 12.68 (11.15–15.69) | 12.40 (10.83–15.97) | 11.29 (9.97–14.14) | 14.85 (13.02–18.34) |
| Young’s Modulus (Mpa) | 393.65 (320.31–589.37) | 512.28 (434.07–845.60) | 313.63 (271.71–426.35) b | 303.84 (256.47–416.28) b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, A.M.M.; Cavalcante, J.G.T.; Bottaro, M.; Vieira, D.C.L.; Babault, N.; Geremia, J.M.; Corrigan, P.; Silbernagel, K.G.; Durigan, J.L.Q.; Marqueti, R.d.C. The Influence of Hip and Knee Joint Angles on Quadriceps Muscle-Tendon Unit Properties during Maximal Voluntary Isometric Contraction. Int. J. Environ. Res. Public Health 2023, 20, 3947. https://doi.org/10.3390/ijerph20053947
de Sousa AMM, Cavalcante JGT, Bottaro M, Vieira DCL, Babault N, Geremia JM, Corrigan P, Silbernagel KG, Durigan JLQ, Marqueti RdC. The Influence of Hip and Knee Joint Angles on Quadriceps Muscle-Tendon Unit Properties during Maximal Voluntary Isometric Contraction. International Journal of Environmental Research and Public Health. 2023; 20(5):3947. https://doi.org/10.3390/ijerph20053947
Chicago/Turabian Stylede Sousa, Alessandra Martins Melo, Jonathan Galvão Tenório Cavalcante, Martim Bottaro, Denis César Leite Vieira, Nicolas Babault, Jeam Marcel Geremia, Patrick Corrigan, Karin Grävare Silbernagel, João Luiz Quaglioti Durigan, and Rita de Cássia Marqueti. 2023. "The Influence of Hip and Knee Joint Angles on Quadriceps Muscle-Tendon Unit Properties during Maximal Voluntary Isometric Contraction" International Journal of Environmental Research and Public Health 20, no. 5: 3947. https://doi.org/10.3390/ijerph20053947
APA Stylede Sousa, A. M. M., Cavalcante, J. G. T., Bottaro, M., Vieira, D. C. L., Babault, N., Geremia, J. M., Corrigan, P., Silbernagel, K. G., Durigan, J. L. Q., & Marqueti, R. d. C. (2023). The Influence of Hip and Knee Joint Angles on Quadriceps Muscle-Tendon Unit Properties during Maximal Voluntary Isometric Contraction. International Journal of Environmental Research and Public Health, 20(5), 3947. https://doi.org/10.3390/ijerph20053947