
Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents
 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material Used in the Research
2.1.1. Plant Material
2.1.2. Potato Phytopathogens
2.1.3. Lactic Acid Bacteria Biopreparation (LpB)
2.1.4. Metschnikowia Pulcherrima Yeast Biopreparation (MpB)
2.1.5. Aqueous Garlic Extract Biopreparation (GB)
2.2. Sample Inoculation with Phytopathogens and Application of Bioprotective Agents
2.3. Fungal Metabolite Analysis by LC–MS/MS
2.4. Growth and Physiological Parameter Analysis
2.4.1. Methods of Plant Cultivation
2.4.2. Assessment of Growth and Physiological Activity Parameters
- Biological condition of the seed potatoes—their vigour, turgor, rotting, and infection by phytopathogens [42];
- The percentage of germinating tubers [42];
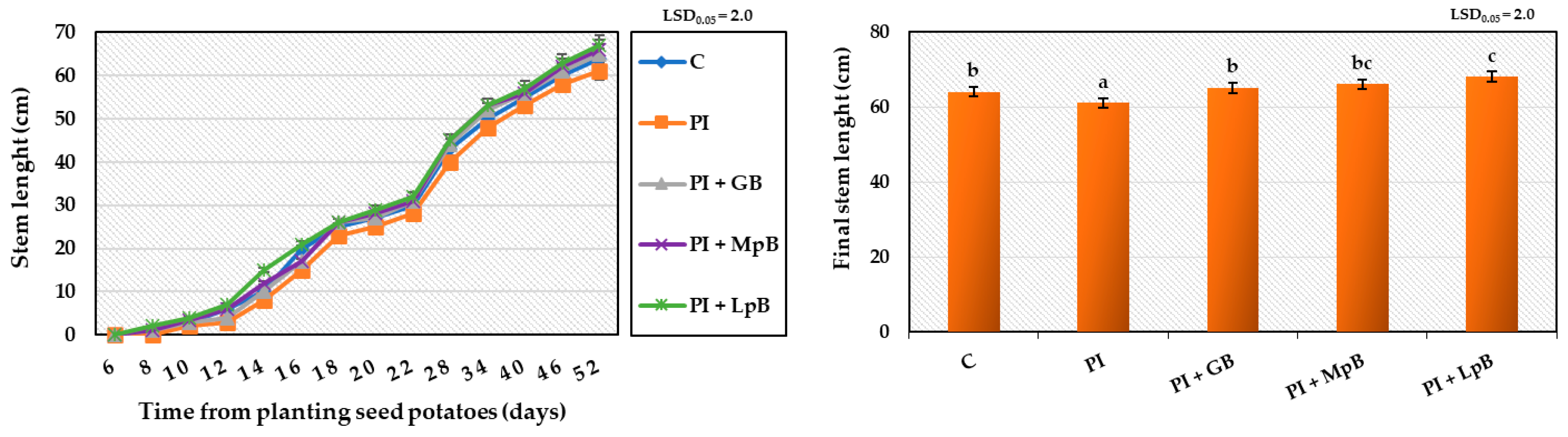
- Growth kinetics of the plant, by measuring the length of shoots every 2 days during the first 24 days, and every 6 days for the remaining period [43];
- Quality of the shoots on a five-point valuation scale, where 5 indicates shoots growing well, full of vigour, while 1 indicates dried out shoots [44];
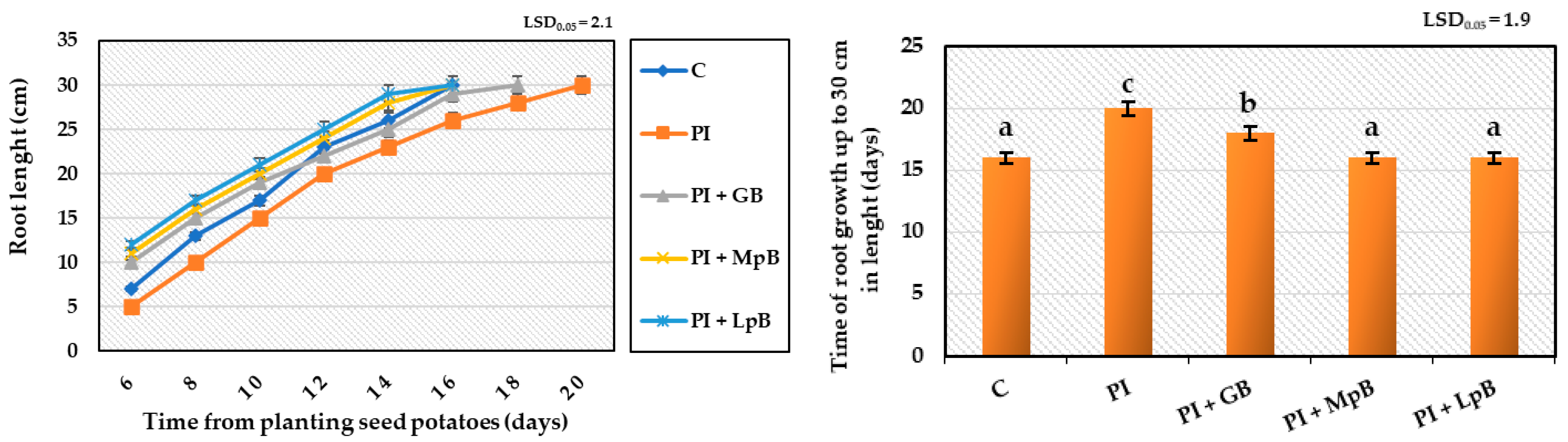
- Quality and growth kinetics of root, by measuring root length every 2 days until they reached 30 cm [42];
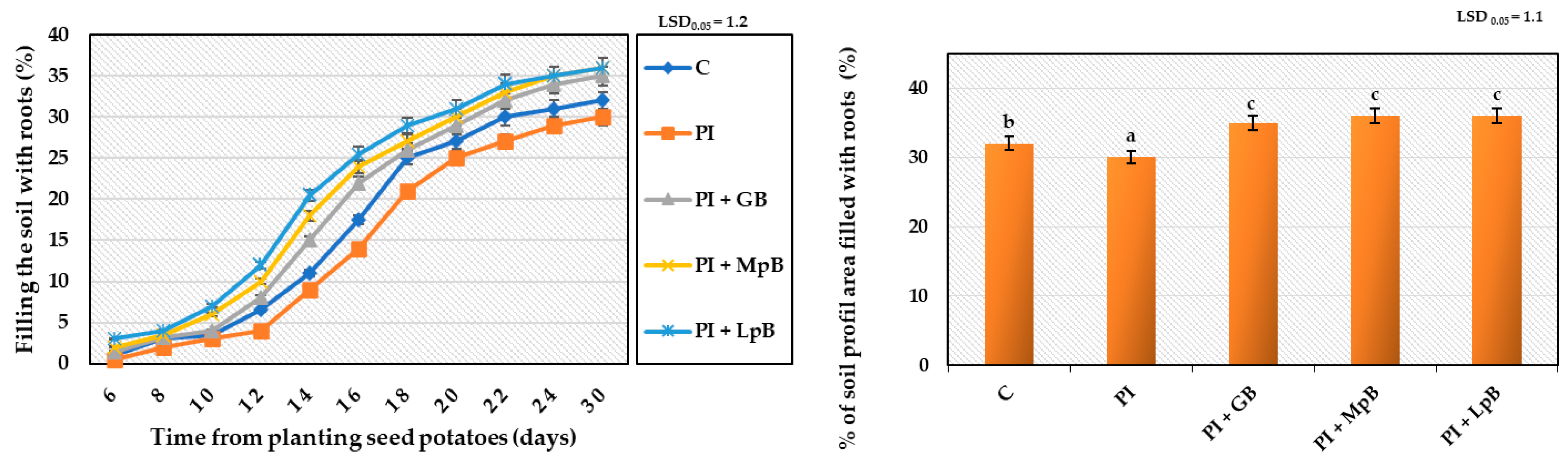
- Growth kinetics of the root system, by assessing every 2 days the percentage of the soil profile area filled by the roots [45];
- Quality of plants on a five-point rating scale, where 5 indicates plants with high vigour, coloured and growing properly, and 1 indicates dried out plants [8];
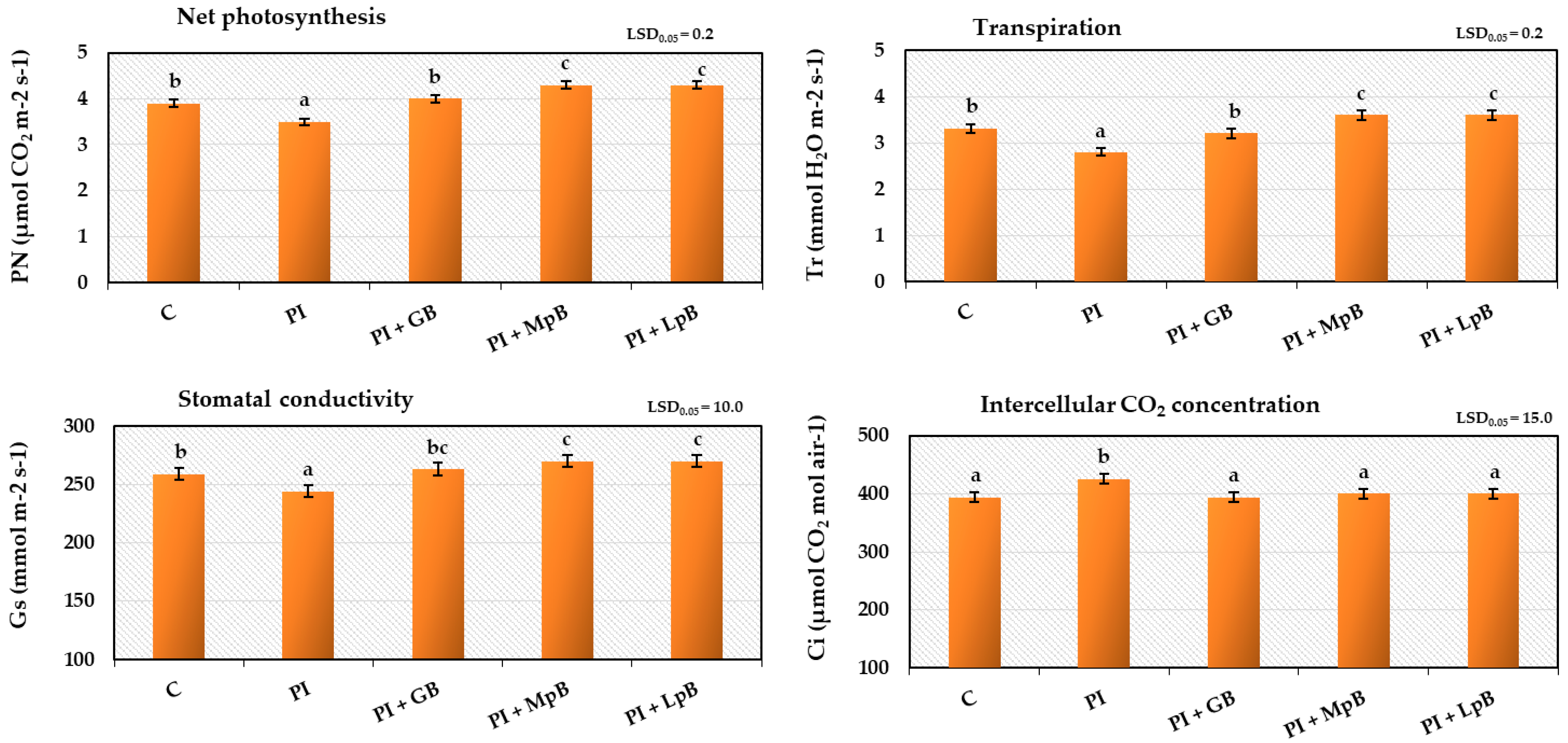
- Intensity of gas exchange (transpiration, net photosynthesis, intercellular CO2 content, and stomata conductivity), measured with a TPS-2 apparatus (PP Systems, USA) in the highest positioned fully developed leaves, during the period of most dynamic plant growth [39];
- Index of chlorophyll content, measured with a Minolta SPAD-502 apparatus (Japan) in the highest positioned fully developed leaves, during the period of most dynamic plant growth [44].
2.5. Statistical Analysis
3. Results and Discussion
3.1. Fungal Metabolite Analysis by LC–MS/MS
3.2. Growth and Physiological Parameter Analysis
3.2.1. Germination of Seed Potatoes and Stem and Root Development
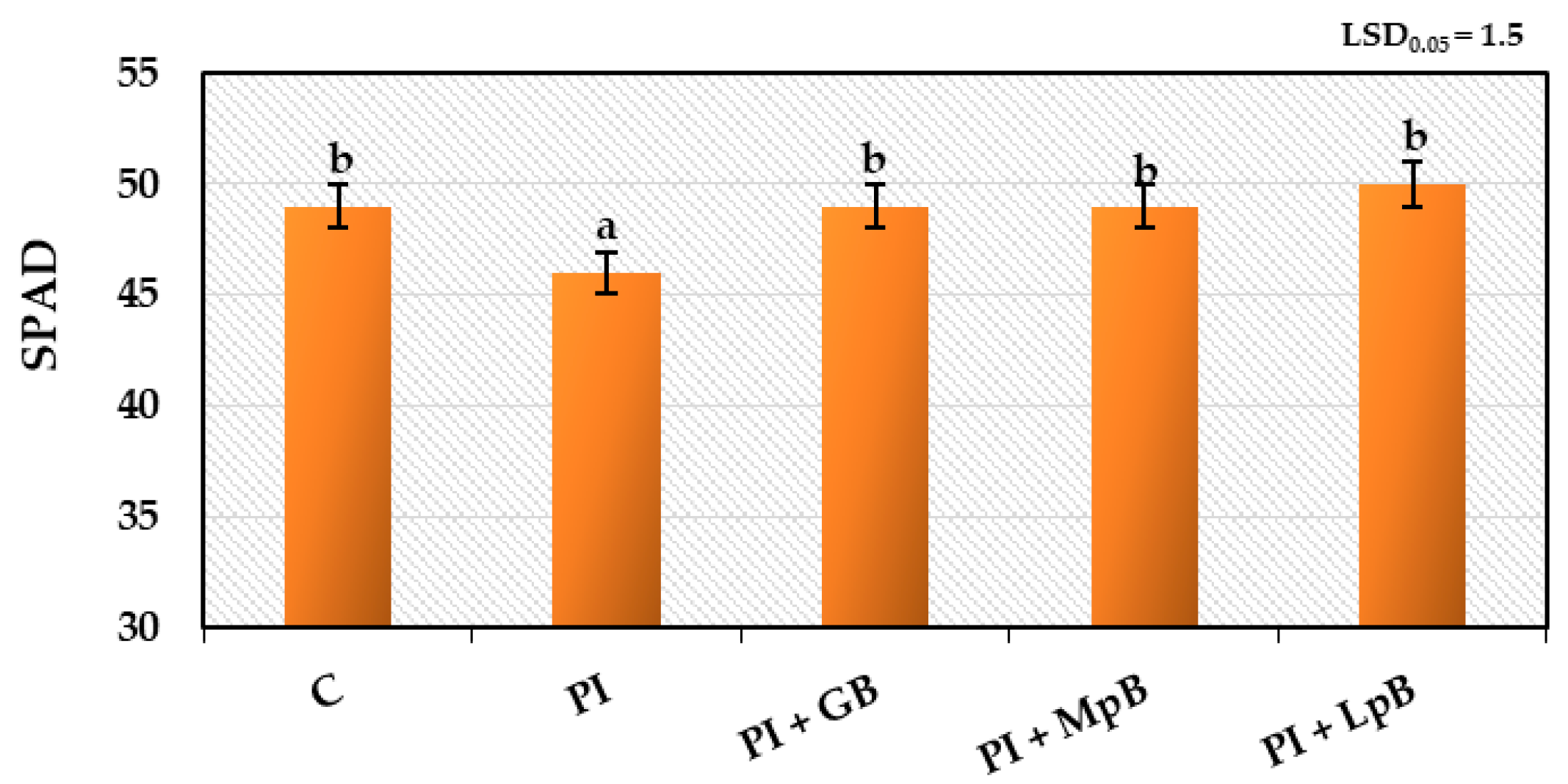
3.2.2. Gas Exchange in Leaves and Index of Chlorophyll Content in Leaves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and Human Health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef]
- FAO. Top Countries in Potatoes Production. Potatoes Production—Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2019; Available online: https://www.fao.org/faostat/en/ (accessed on 6 January 2023).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Z.; Zou, Y.; Li, W.; He, F.; Huang, X.; Lin, C.; Cai, Q.; Wisniewski, M.; Wu, X. Pre- and postharvest measures used to control decay and mycotoxigenic fungi in potato (Solanum tuberosum L.) during storage. Crit. Rev. Food Sci. Nutr. 2021, 62, 415–428. [Google Scholar] [CrossRef]
- Kebede, H.; Liu, X.; Jin, J.; Xing, F. Current status of major mycotoxins contamination in food and feed in Africa. Food Control 2020, 110, 106975. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Saari, N.; Meor Hussin, A.S. Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply. Molecules 2020, 25, 2655. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Galani Yamdeu, J.H.; Gong, Y.Y.; Orfila, C. A review of postharvest approaches to reduce fungal and mycotoxin contamination of foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1521–1560. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on Agriculture, Food, and Human Health and their Management Strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC). No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. 2006. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:364:0005:0024:EN:PDF (accessed on 6 January 2023).
- Xue, H.L.; Bi, Y.; Tang, Y.M.; Zhao, Y.; Wang, Y. Effect of cultivars, Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers. Food Chem. 2014, 151, 236–242. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Diversity of Fusarium spp. associated with dry rot of potato tubers in Poland. Eur. J. Plant Pathol. 2016, 145, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Delgado, J.A.; Schwarz, P.B.; Gillespie, J.; Rivera-Varas, V.V.; Secor, G.A. Trichothecene mycotoxins associated with potato dry rot caused by fusarium graminearum. Phytopathology 2010, 100, 290–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, I.; Naehrer, K. A Three-Year Survey on the Worldwide Occurrence of Mycotoxins in Feedstuffs and Feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-Produced Mycotoxins in Plant-Pathogen Interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canadian Horticultural Council. Control of Potato Storage Conditions for the Management of Post-Harvest Losses due to Diseases; Canadian Horticultural Council: Ottawa, ON, Canada, 2017. [Google Scholar]
- Steglińska, A.; Kołtuniak, A.; Motyl, I.; Berłowska, J.; Czyżowska, A.; Cieciura-Włoch, W.; Okrasa, M.; Kręgiel, D.; Gutarowska, B. Lactic Acid Bacteria as Biocontrol Agents against Potato (Solanum tuberosum L.) Pathogens. Appl. Sci. 2022, 12, 7763. [Google Scholar] [CrossRef]
- Steglińska, A.; Kołtuniak, A.; Berłowska, J.; Czyżowska, A.; Szulc, J.; Cieciura-Włoch, W.; Okrasa, M.; Kręgiel, D.; Gutarowska, B. Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens. Agronomy 2022, 12, 2546. [Google Scholar] [CrossRef]
- Steglińska, A.; Bekhter, A.; Wawrzyniak, P.; Kunicka-Styczyńska, A.; Jastrząbek, K.; Fidler, M.; Śmigielski, K.; Gutarowska, B. Antimicrobial Activities of Plant Extracts against Solanum tuberosum L. Phytopathogens. Molecules 2022, 27, 1579. [Google Scholar] [CrossRef]
- El-Mabrok, A.S.W.; Hassan, Z.; Mokhtar, A.M.; Hussin, K.M.A. Antifungal activity of Lactobacillus plantarum LAB-C5 and LAB-G7 isolated from Malaysian fruits. Acta Biol. Malaysiana 2013, 2, 22–30. [Google Scholar] [CrossRef]
- Canpolat, E.; Doğaner, M.M.; Derviş, S.; Ulubaş Serçe, Ç. Antifungal Activity of Some Lactic Acid Bacteria Against Several Soil-borne Fungal Pathogens Isolated from Strawberry Plants. Turkish J. Agric.-Food Sci. Technol. 2018, 6, 1163. [Google Scholar] [CrossRef]
- Abdel-Aziz, S.M.; Moustafa, Y.A.; Hamed, H.A. Lactic Acid Bacteria in the Green Biocontrol against some Phytopathogenic Fungi: Treatment of Tomato Seeds. J. Basic. Appl. Sci. Res 2014, 4, 1–9. [Google Scholar]
- Šušković, J.; Kos, B.; Beganović, J.; Pavunc, A.L.; Habjanič, K.; Matoć, S. Antimicrobial activity—The most important property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Gerez, C.L.; Torres, M.J.; Font de Valdez, G.; Rollán, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control 2013, 64, 231–237. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Perczak, A.; Goliński, P.; Bryła, M.; Waśkiewicz, A. The efficiency of lactic acid bacteria against pathogenic fungi and mycotoxins. Arh. Hig. Rada Toksikol. 2018, 69, 32–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendonca-Hagler, L.C.; Hagler, A.N.; Kurtzman, C.P. Phylogeny of Metschnikowia Species Estimated from Partial rRNA Sequences. Int. J. Syst. Bacteriol. 1993, 43, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Zhu, L.; Hou, X. Combination of UV-C Treatment and Metschnikowia pulcherrimas for Controlling Alternaria Rot in Postharvest Winter Jujube Fruit. J. Food Sci. 2015, 80, M137–M141. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Li, W.; Jiang, Z.T.; Jing, M.M.; Shao, Y.Z. The preservation effect of Metschnikowia pulcherrima yeast on anthracnose of postharvest mango fruits and the possible mechanism. Food Sci. Biotechnol. 2018, 27, 95–105. [Google Scholar] [CrossRef]
- No. P.441896, Method of Bio-Protection of Seed Potato Tubers against the Development of Bacterial and Fungal Phytopathogens. Pat. Appl. Pat. Off. Pol. 2022. Available online: https://uprp.gov.pl/pl (accessed on 6 January 2023).
- No. P.441895: Method of Bio-Protection of Seed Potato Tubers against the Development of Fungal Phytopathogens. Pat. Appl. Pat. Off. Pol. 2022. Available online: https://uprp.gov.pl/pl (accessed on 6 January 2023).
- Sharma, A.; Rajendran, S.; Srivastava, A.; Sharma, S.; Kundu, B. Antifungal activities of selected essential oils against Fusarium oxysporum f. sp. lycopersici 1322, with emphasis on Syzygium aromaticum essential oil. J. Biosci. Bioeng. 2017, 123, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Abd-El-Khair, H.; El-Gamal Nadia, G. Effects of aqueous extracts of some plant species against Fusarium solani and Rhizoctonia solani in Phaseolus vulgaris plants. Arch. Phytopathol. Plant Prot. 2011, 44, 1–16. [Google Scholar] [CrossRef]
- Nashwa, S.M.A.; Abo-Elyou, K.A.M. Evaluation of various plant extracts against the early blight disease of tomato plants under greenhouse and field conditions. Plant Prot. Sci. 2012, 48, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Martins, N.; Petropoulos, S.; Ferreira, I.C.F.R. Chemical composition and bioactive compounds of garlic (Allium sativum L.) as affected by pre- and post-harvest conditions: A review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of > 500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef] [Green Version]
- Romanowska-Duda, Z.; Janas, R.; Grzesik, M. Application of Phytotoxkit in the quick assessment of ashes suitability as fertilizers in sorghum crops. Int. Agrophysics 2019, 33, 145–152. [Google Scholar] [CrossRef]
- Grzesik, M.; Janas, R.; Steglińska, A.; Kręgiel, D.; Gutarowska, B. Influence of microclimatic conditions during year-long storage of ‘Impresja’ potato tubers (Solanum tuberosum L.) on the emergence, growth, physiological activity and yield of plants. J. Stored Prod. Res. 2022, 99, 102033. [Google Scholar] [CrossRef]
- Steglińska, A.; Pielech-Przybylska, K.; Janas, R.; Grzesik, M.; Borowski, S.; Kręgiel, D.; Gutarowska, B. Volatile Organic Compounds and Physiological Parameters as Markers of Potato (Solanum tuberosum L.) Infection with Phytopathogens. Molecules 2022, 27, 3708. [Google Scholar] [CrossRef] [PubMed]
- Janas, R.; Górnik, K.; Grzesik, M.; Romanowska-Duda, Z.; van Duijn, B. Effectiveness of pulsed radio frequency in seed quality improvement of vegetable plant species. Int. Agrophysics 2019, 33, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Romanowska-Duda, Z.; Szufa, S.; Grzesik, M.; Piotrowski, K.; Janas, R. The Promotive Effect of Cyanobacteria and Chlorella sp. Foliar Biofertilization on Growth and Metabolic Activities of Willow (Salix viminalis L.) Plants as Feedstock Production, Solid Biofuel and Biochar as C Carrier for Fertilizers via Torrefaction Proce. Energies 2021, 14, 5262. [Google Scholar] [CrossRef]
- Grzesik, M.; Górnik, K.; Janas, R.; Lewandowki, M.; Romanowska-Duda, Z.; Duijn, B. van High efficiency stratification of apple cultivar Ligol seed dormancy by phytohormones, heat shock and pulsed radio frequency. J. Plant Physiol. 2017, 219, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Romanowska-Duda, Z.; Grzesik, M.; Janas, R. Maximal Efficiency of PSII as a Marker of Sorghum Development Fertilized With Waste From a Biomass Biodigestion to Methane. Front. Plant Sci. 2019, 9, 1920. [Google Scholar] [CrossRef] [Green Version]
- Spanic, V.; Katanic, Z.; Sulyok, M.; Krska, R.; Puskas, K.; Vida, G.; Drezner, G.; Šarkanj, B. Multiple fungal metabolites including mycotoxins in naturally infected and fusarium-inoculated wheat samples. Microorganisms 2020, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Kuca, K.; Nepovimova, E.; Wu, W. Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink. Toxins 2020, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Paciolla, C.; Dipierro, N.; Mulè, G.; Logrieco, A.; Dipierro, S. The mycotoxins beauvericin and T-2 induce cell death and alteration to the ascorbate metabolism in tomato protoplasts. Physiol. Mol. Plant Pathol. 2004, 65, 49–56. [Google Scholar] [CrossRef]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, Y.; Yu, S. Sambutoxin-producing isolates of fusarium species and occurrence of sambutoxin in rotten potato tubers. Appl. Environ. Microbiol. 1995, 61, 3750–3751. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.C.; Lee, Y.W. Sambutoxin, a new mycotoxin produced by toxic Fusarium isolates obtained from rotted potato tubers. Appl. Environ. Microbiol. 1994, 60, 4380–4386. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Upadhyay, R.S. Fusaric acid induced cell death and changes in oxidative metabolism of Solanum lycopersicum L. Bot. Stud. 2014, 55, 66. [Google Scholar] [CrossRef] [Green Version]
- Frandsen, R.J.N.; Schütt, C.; Lund, B.W.; Staerk, D.; Nielsen, J.; Olsson, S.; Giese, H. Two Novel Classes of Enzymes Are Required for the Biosynthesis of Aurofusarin in Fusarium graminearum. J. Biol. Chem. 2011, 286, 10419–10428. [Google Scholar] [CrossRef] [Green Version]
- Kokkonen, M.; Jestoi, M.; Rizzo, A. The effect of substrate on mycotoxin production of selected Penicillium strains. Int. J. Food Microbiol. 2005, 99, 207–214. [Google Scholar] [CrossRef]
- Westphal, K.R.; Heidelbach, S.; Zeuner, E.J.; Riisgaard-Jensen, M.; Nielsen, M.E.; Vestergaard, S.Z.; Bekker, N.S.; Skovmark, J.; Olesen, C.K.; Thomsen, K.H.; et al. The effects of different potato dextrose agar media on secondary metabolite production in Fusarium. Int. J. Food Microbiol. 2021, 347, 109171. [Google Scholar] [CrossRef]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary Fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- De Souza, G.D.; Mithöfer, A.; Daolio, C.; Schneider, B.; Rodrigues-Filho, E. Identification of Alternaria alternata mycotoxins by LC-SPE-NMR and their cytotoxic effects to soybean (Glycine max) cell suspension culture. Molecules 2013, 18, 2528–2538. [Google Scholar] [CrossRef] [Green Version]
- Robles, A.J.; Dai, W.; Haldar, S.; Ma, H.; Anderson, V.M.; Overacker, R.D.; Risinger, A.L.; Loesgen, S.; Houghton, P.J.; Cichewicz, R.H.; et al. Altertoxin II, a Highly Effective and Specific Compound against Ewing Sarcoma. Cancers 2021, 13, 6176. [Google Scholar] [CrossRef]
- Wang, H.; Guo, Y.; Luo, Z.; Gao, L.; Li, R.; Zhang, Y.; Kalaji, H.M.; Qiang, S.; Chen, S. Recent Advances in Alternaria Phytotoxins: A Review of Their Occurrence, Structure, Bioactivity and Biosynthesis. J. Fungi 2022, 8, 168. [Google Scholar] [CrossRef]
- Aichinger, G.; Del Favero, G.; Warth, B.; Marko, D. Alternaria toxins—Still emerging? Compr. Rev. Food Sci. Food Saf. 2021, 20, 4390–4406. [Google Scholar] [CrossRef]
- Pusztahelyi, T.; Holb, I.J.; PÃ3csi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Hou, H.; Liu, H.; Wang, H.; Cai, L.; An, M.; Zhang, C. Identification of 3-Methoxyphenylacetic Acid as a Phytotoxin, Produced by Rhizoctonia solani AG-3 TB. Molecules 2023, 28, 790. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-T.; Chou, C.-T.; Chen, I.-S.; Yu, C.-C.; Lu, T.; Hsu, S.-S.; Shieh, P.; Jan, C.-R.; Liang, W.-Z. Mechanisms underlying effect of the mycotoxin cytochalasin B on induction of cytotoxicity, modulation of cell cycle, Ca2+ homeostasis and ROS production in human breast cells. Toxicology 2016, 370, 1–19. [Google Scholar] [CrossRef]
- Mendes, G.; Baltazar, L.M.; Souza, D.G.; Sá, N.P.; Rosa, L.H.; Rosa, C.A.; Souza-Fagundes, E.M.; Ramos, J.P.; Alves-Silva, J.; Cota, B.B.; et al. Effects of cytochalasin E on Paracoccidioides brasiliensis. J. Appl. Microbiol. 2018, 125, 1296–1307. [Google Scholar] [CrossRef]
- Kirk, W.; Gachango, E.; General Seed-Rots Can Pose Problems in Potatoes. Michigan State Univ. Ext. Dep. Plant Pathol. 2011. Available online: https://www.canr.msu.edu/news/general_seed_rots_can_pose_problems_in_potatoes (accessed on 20 December 2022).
- Hamed, H.A.; Moustafa, Y.A.; Abdel-Aziz, S.M. In vivo efficacy of lactic acid bacteria in biological control against Fusarium oxysporum for protection of tomato plant. Life Sci. J. 2011, 8, 462–468. [Google Scholar]
- Koroney, A.S.; Plasson, C.; Pawlak, B.; Sidikou, R.; Driouich, A.; Menu-Bouaouiche, L.; Vicré-Gibouin, M. Root exudate of Solanum tuberosum is enriched in galactose-containing molecules and impacts the growth of Pectobacterium atrosepticum. Ann. Bot. 2016, 118, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Badek, B.; Romanowska-Duda, Z.; Grzesik, M.; Duijn, B. van Rapid Evaluation of Germinability of Primed China Aster (Callistephus Chinensis Ness.) Seeds with Physiological and Biochemical Markers. J. Hortic. Res. 2014, 22, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Badek, B.; Romanowska-Duda, Z.; Grzesik, M.; Kuras, A. Physiological Markers for Assessing Germinability of Lycopersicon esculentum Seeds Primed by Environment-Friendly Methods. Polish J. Environ. Stud. 2016, 25, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Fu, J.; Chen, Z.; Chen, C.; Zhang, J.; Ren, L. Measurement and Analysis of Root Anchorage Effect on Stalk Forces in Lodged Corn Harvesting. Front. Plant Sci. 2022, 13, 852375. [Google Scholar] [CrossRef] [PubMed]
- Grzesik, M.; Romanowska-Duda, Z.; Kalaji, H.M. Effectiveness of cyanobacteria and green algae in enhancing the photosynthetic performance and growth of willow (Salix viminalis L.) plants under limited synthetic fertilizers application. Photosynthetica 2017, 55, 510–521. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.C.; Asanuma, K.; Kusutani, A.; Toyota, M. Gas Exchange Properties of Potato Plants under Different Nitrogen Regimes. Pakistan J. Biol. Sci. 2000, 3, 1654–1657. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Concentration Mean ± SD [ng/g] | ||||
|---|---|---|---|---|---|
| F. sambucinum Culture on MEA Plate (n = 3) | F. sambucinum Culture on Potato (n = 3) | Potatoes Treated with Biological Preparations and Infected with F. sambucinum | |||
| GB (n = 3) | LpB (n = 3) | MpB (n = 3) | |||
| Abscisic acid | <LOD b | <LOD b | <LOD b | <LOD b | 30.2 ± 20.4 a |
| Acuminatum B | 20,500 ± 584 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Acuminatum C | 9380 ± 862 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Asterric acid | <LOD a | 83.3 ± 112 a | <LOD a | 50.9 ± 9.73 a | <LOD a |
| Aurofusarin | <LOD b | 7.27 ± 2.49 a | <LOD b | 3.50 ± 0.0718 a | <LOD b |
| Beauvericin1 | 6.3 ± 2 a | 27.5 ± 17.8 a | 73.8 ± 63.6 a | 11.8 ± 15.9 a | 2.21 ± 1.55 a |
| Bikaverin 1 | <LOD a | 505 ± 510 a | 5.04 ± 1.40 a | <LOD a | 31.1 ± 29.5 a |
| Brevianamide F 2 | 20.8 ± 1.13 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Chaconin | <LOD a | <LOD a | 1320 ± 1070 a | 2240 ± 1740 ab | 5560 ± 2080 b |
| Chrysogin | 14.8 ± 4.59 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Citreorosein | <LOD b | <LOD b | <LOD b | 1.54 ± 0.128 a | <LOD b |
| Cordycepin 1,2 | <LOD a | 65.6 ± 11 a | 106 ± 120 a | 169 ± 144 a | 43.5 ± 7.32 a |
| cyclo(L-Pro-L-Tyr) 2 | 67.2 ± 14.9 a | <LOD b | <LOD b | <LOD b | <LOD b |
| cyclo(L-Pro-L-Val) 2 | 216 ± 10.7 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Cyclosporin A | <LOD b | <LOD b | 948 ± 407 a | <LOD b | <LOD b |
| Cyclosporin B | <LOD b | <LOD b | 114 ± 80.2 a | <LOD b | <LOD b |
| Cyclosporin D | <LOD b | <LOD b | 50.8 ± 27.4 a | <LOD b | <LOD b |
| Cyclosporin H | <LOD b | <LOD b | 783 ± 338 a | <LOD b | <LOD b |
| Deoxygerfelin 1 | <LOD a | 34.3 ± 33.2 a | <LOD a | 0.668 ± 0.679 a | 10.8 ± 10.6 a |
| Diacetoxyscirpenol | 147.1 ± 17.3 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Dinactin | <LODb | <LOD b | <LOD b | <LOD b | 3.04 ± 1.72 a |
| Emodin 1 | <LOD a | 4.31 ± 2.90 a | <LOD a | 1.65 ± 2.35 a | 7.64 ± 7.36 a |
| Endocrocin | <LOD ab | 203 ± 113 cd | <LOD ae | 31.9 ± 3.61 ace | 63.7 ± 64.8 bde |
| Equisetin | <LOD b | <LOD b | <LOD b | <LOD b | 1.67 ± 1.07 a |
| Fallacinol | <LOD b | <LOD b | <LOD b | 2.78 ± 2.08 a | <LOD b |
| Fusaric acid | <LOD a | 370 ± 133 a | 7870 ± 6990 a | <LOD a | <LOD a |
| Gibepyron D | <LOD b | <LOD b | 46.5 ± 34.3 a | <LOD b | <LOD b |
| Iso-Rhodoptilometrin | <LOD a | 24.4 ± 21 a | <LOD a | 3.92 ± 2.56 a | 4.87 ± 5.47 a |
| Isosulochrin 1 | <LOD a | 25.9 ± 26.8 a | <LOD a | 28.2 ± 40.7 a | <LOD a |
| Lecanoric acid 1 | <LOD a | 232 ± 262 a | <LOD a | 27.6 ± 34.1 a | 218 ± 305 a |
| Monactin | <LOD b | <LOD b | <LOD b | <LOD b | 26.7 ± 256 a |
| Nonactin | <LOD a | <LOD a | <LOD a | <LOD a | 2.61 ± 2.04 a |
| Radicinin | <LOD b | <LOD b | 1.14 ± 0.16 a | <LOD b | <LOD b |
| Sambutoxin | <LOD b | 0.29 ± 0.15 a | 0.0220 ± 0.0113 b | <LOD b | <LOD b |
| Secalonic acid D | <LOD a | <LOD a | <LOD a | 3.53 ± 1.93 b | 2.57 ± 2.24 ab |
| Solanin 1 | <LOD a | 3250 ± 2000 ab | 462 ± 393 a | 1970 ± 1670 ab | 7820 ± 5010 b |
| Sterigmatocystin | <LOD b | <LOD b | <LOD b | <LOD b | 0.238 ± 0.228 a |
| Sulochrin 1 | <LOD a | 60.9 ± 68.2 a | <LOD a | 91.9 ± 108 a | <LOD a |
| Terragine 1 | <LOD a | 1310 ± 795 a | 2830 ± 3080 a | 8350 ± 13,300 a | 944 ± 181 a |
| Tryptophol 1,2 | <LOD b | <LOD b | <LOD b | <LOD b | 14.2 ± 4.75 a |
| Violaceol I | <LOD a | 1670 ± 1970 a | <LOD a | 1120 ± 1380 a | 6080 ± 7270 a |
| Violaceol II 1 | <LOD a | 17.7 ± 20.3 a | <LOD a | <LOD a | <LOD a |
| Vulpinic acid | <LOD b | <LOD b | <LOD b | <LOD b | 0.0993 ± 0.0542 a |
| Xanthoquinoidin A1 | <LOD a | <LOD a | <LOD a | <LOD a | 24.8 ± 18.5 a |
| Components | Concentration Mean ± SD [ng/g] | ||||
|---|---|---|---|---|---|
| A. tenuissima Culture on MEA Plate (n = 3) | A. tenuissima Culture on Potato (n = 3) | Potatoes Treated with Biological Preparations and Infected with A. tenuissima | |||
| GB (n = 3) | LpB (n = 3) | MpB (n = 3) | |||
| 4-Hydroxyalternariol | 32.7 ± 10.2 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Abscisic acid | <LOD a | <LOD a | 40.6 ± 10.2 d | 21.9 ± 14.4 ad | 33.9 ± 4.57 d |
| Alteichin | <LOD c | <LOD c | 8.98 ± 1.14 a | 3.97 ± 1.83 b | <LOD c |
| Alternariol (AOH) | 935 ± 90.7 a | <LOD b | 11.8 ± 12.2 b | <LOD b | 0.451 ± 0.222 b |
| Alternariolmethylether (AME) | 18.6 ± 4.54 a | 0.57 ± 0.55 b | 11.4 ± 12.6 ab | <LOD b | 0.889 ± 0.352 b |
| Altertoxin-I (ALX-1) | 1030 ± 9.32 a | <LOD c | 15.4 ± 2.79 b | 2.98 ± 0.85 bc | 5.45 ± 3.44 bc |
| Asterric acid | <LOD a | <LOD a | 58.3 ± 75.1 a | 192.5 ± 148.6 a | 91.7 ± 111 a |
| Beauvericin1 | <LOD a | 46.3 ± 68.6 a | 8.98 ± 8.08 a | 6.41 ± 6.18 a | 4.91 ± 3.23 a |
| Bikaverin1 | <LOD b | 16.6 ± 19.2 b | 265 ± 79.4 a | 78.1 ± 109.2 b | 86.9 ± 53.8 b |
| Brevianamid F 2 | 9.36 ± 0.353 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Chaconin | <LOD a | <LOD a | 5710 ± 700 b | 5460 ± 2060 b | 7680 ± 2510 b |
| Citreorosein | <LOD b | <LOD b | 1.87 ± 1.54 ab | 2.34 ± 0.20 ab | 0.993 ± 0.0613 ab |
| Cordycepin 1,2 | 13.8 ± 1.68 a | 128 ± 120 a | 44.0 ± 4.51 a | 50.6 ± 7.87 a | 32.9 ± 4.91 a |
| cyclo(L-Leu-L-Pro) | 4.57 ± 1.54 a | <LOD b | <LOD b | <LOD b | <LOD b |
| cyclo(L-Pro-L-Tyr) 2 | 58.6 ± 10.2 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Cytochalasin B | 5.81 ± 3.27 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Deoxygerfelin 1 | <LOD a | <LOD a | 7.46 ± 7.71 a | 4.28 ± 5.23 a | 3.60 ± 2.96 a |
| Dinactin | <LOD a | <LOD a | <LOD a | <LOD a | 0.871 ± 1.08 a |
| Emodin 1 | <LOD b | <LOD b | 2.92 ± 2.40 ab | 3.92 ± 1.20 a | 2.34 ± 1.58 ab |
| Endocrocin | <LOD a | <LOD a | <LOD a | 54.7 ± 52.3 a | <LOD a |
| Fallacinol | <LOD b | <LOD b | 1.20 ± 1.63 ab | 2.57 ± 0.38 a | 1.50 ± 0.975 ab |
| Fusaric acid | <LOD a | 3820 ± 3850 a | 23.4 ± 15.1 a | 82.2 ± 65.2 a | <LOD a |
| Iso-Rhodoptilometrin | <LOD b | 0.478 ± 0.0493 bc | 0.91 ± 0.65 bc | 2.66 ± 1.49 ac | 1.07 ± 1.10 bc |
| Isosulochrin 1 | <LOD a | <LOD a | 7.26 ± 9.28 a | 31.2 ± 39.1 a | 0.80 ± 0.70 a |
| Lecanoric acid1 | <LOD a | <LOD a | 66.6 ± 42.3 a | 32.4 ± 24.1 a | 43.8 ± 56.9 a |
| Sambutoxin | <LOD a | 0.0484 ± 0.0235 a | 0.0511 ± 0.045 a | <LOD a | 0.0290 ± 0.0222 a |
| Secalonic acid D | <LOD a | <LOD a | 12.7 ± 13.5 a | <LOD a | 15.6 ± 11.7 a |
| Solanin 1 | <LOD a | 31,500 ± 38,500 a | 7090 ± 1310 a | 9110 ± 6350 a | 19,100 ± 17,000 a |
| Sulochrin 1 | <LOD a | <LOD a | 13.0 ± 17.1 a | 65.0 ± 82.9 a | 12.8 ± 15.0 a |
| Tentoxin (TX) | <LOD a | 27.2 ± 30 a | 12.3 ± 6.79 a | 11.3 ± 5.52 a | 31.4 ± 45.2 a |
| Terragine 1 | <LOD a | <LOD a | <LOD a | 716 ± 179 b | 685 ± 25.5 b |
| Tryptophol 1,2 | <LOD a | <LOD a | 18.0 ± 5.83 b | 9.42 ± 4.42 ab | 17.0 ± 2.95 b |
| Violaceol I | <LOD a | <LOD a | 5730 ± 6340 a | 1850 ± 1570 a | 8280 ± 13,100 a |
| Components | Concentration Mean ± SD [ng/g] | ||||
|---|---|---|---|---|---|
| R. solani Culture on MEA Plate (n = 3) | R. solani Culture on Potato (n = 3) | Potatoes Treated with Biological Preparations and Infected with R. solani | |||
| GB (n = 3) | LpB (n = 3) | MpB (n = 3) | |||
| Abscisic acid | <LOD a | <LOD a | 30.8 ± 11.5 a | 37.5 ± 43.0 a | 27.2 ± 6.88 a |
| Beauvericin A | <LOD b | <LOD b | 0.33 ± 0.20 b | 1.27 ± 0.39 a | 0.299 ± 0.102 b |
| Beauvericin 1 | <LOD a | 36.1 ± 31.8 a | 7.15 ± 3.43 a | 53.7 ± 50.7 a | 24.5 ± 24.6 a |
| Bikaverin 1 | <LOD a | 23.6 ± 26.3 a | 116 ± 131 a | 48.8 ± 59.9 a | 164 ± 77.1 a |
| Brevianamide F 2 | 2.47 ± 1.69 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Chaconine | <LOD b | <LOD b | 9320 ± 3060 a | 7890 ± 1910 a | 7760 ± 3100 a |
| Citreorosein | <LOD b | <LOD b | <LOD b | <LOD b | 2.17 ± 0.49 a |
| Cordycepin 1,2 | <LOD cd | 124 ± 34.1 a | 43.9 ± 6.3 b c | 60.2 ± 28.1 b | 38.8 ± 17.3 bd |
| cyclo(L-Leu-L-Pro) | 7.97 ± 0.92 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Deoxygerfelin 1 | <LOD b | <LOD b | 11.8 ± 6.83 a | 1.03 ± 0.76 b | 6.53 ± 3.54 ab |
| Dinactin | <LOD b | <LOD b | <LOD b | 0.71 ± 0.13 a | 0.16 ± 0.13 b |
| Emodin 1 | <LOD a | <LOD a | 1.74 ± 0.89 a | 0.80 ± 0.77 a | 2.08 ± 1.26 a |
| Endocrocin | <LOD a | 49 ± 49.3 a | 29.2 ± 7.76 a | <LOD a | 39.4 ± 14.1 a |
| Enniatin B | <LOD b | <LOD b | 0.111 ± 0.0169 a | <LOD b | <LOD b |
| Fallacinol | <LOD b | <LOD b | <LOD b | <LOD b | 2.45 ± 0.403 a |
| Fusaric acid | <LOD a | 2730 ± 3720 a | <LOD a | 265.9 ± 116.3 a | 114 ± 63.1 a |
| Iso-Rhodoptilometrin | <LOD b | 2.32 ± 2.52 ab | 2.53 ± 1.20 ab | 1.32 ± 1.14 b | 5.55 ± 1.37 a |
| Isosulochrin 1 | <LOD a | <LOD a | 1.96 ± 2.25 a | <LOD a | 9.69 ± 14.7 a |
| Lecanoric acid 1 | <LOD a | <LOD a | 21 ± 22.6 a | 6.15 ± 4.10 a | 37.8 ± 34.8 a |
| Monactin | <LOD b | <LOD b | 3.37 ± 4.26 ab | 11.6 ± 5.40 a | 2.25 ± 1.91 ab |
| Nonactin 1 | <LOD b | <LOD b | 0.60 ± 0.34 b | 1.43 ± 0.47 a | <LOD b |
| Sambutoxin | <LOD a | 0.45 ± 0.44 a | <LOD a | 0.0691 ± 0.0620 a | 0.149 ± 0.133 a |
| Secalonic acid D | <LOD a | <LOD a | <LOD a | 25.7 ± 17.8 a | 46.9 ± 45.3 a |
| Solanine 1 | <LOD a | 17,400 ± 10,300 a | 27,700 ± 17,500 a | 11,000 ± 5030 a | 22,500 ± 23,700 a |
| Sulochrin 1 | <LOD a | <LOD a | <LOD a | <LOD a | 26.0 ± 31.4 a |
| Terragine 1 | <LOD a | 2640 ± 1800 a | 1780 ± 1270 a | 1520 ± 1480 a | 1390 ± 1360 a |
| Tryptophol 1,2 | <LOD b | 13.8 ± 5.52 ab | 15.0 ± 4.31 ab | 25.1 ± 15.1 a | 10.4 ± 5.16 a b |
| Violaceol I | <LOD a | <LOD a | 1870 ± 1720 a | 246 ± 29.2 a | 2070 ± 2480 a |
| Violaceol II 1 | <LOD a | <LOD a | 41.6 ± 30.5 a | 10.2 ± 0.739 a | 30.7 ± 28.9 a |
| Xanthoquinoidin A1 | <LOD a | <LOD a | <LOD a | 22.9 ± 31.1 a | <LOD a |
| Components | Concentration Mean ± SD [ng/g] | ||||
|---|---|---|---|---|---|
| P. exigua Culture on MEA Plate (n = 3) | P. exigua Culture on Potato (n = 3) | Potatoes Treated with Biological Preparations and Infected with P. exigua | |||
| GB (n = 3) | LpB (n = 3) | MpB (n = 3) | |||
| Abscisic acid | <LOD c | <LOD c | 33.2 ± 7.68 ab | 46.3 ± 7.62 a | 29.0 ± 3.03 b |
| Beauvericin A | <LOD a | <LOD a | <LOD a | 0.334 ± 0.314 a | <LOD a |
| Beauvericin 1 | <LOD a | 21.7 ± 32.1 a | 11.1 ± 1.27 a | 7.60 ± 11.9 a | 1.49 ± 0.461 a |
| Bikaverin 1 | <LOD a | 103 ± 142 a | 137 ± 55.4 a | 17.0 ± 5.17 a | 42.45 ± 2.96 a |
| Brevianamide F 2 | 11.0 ± 0.67 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Chaconine | <LOD c | <LOD c | 10,400 ± 662 ab | 7700 ± 1010 a | 13,500 ± 3720 b |
| Citreorosein | <LOD c | <LOD c | 2.34 ± 0.944 b | 4.70 ± 1.18 a | <LOD c |
| Cordycepin 1,2 | 2.76 ± 0.562 a | 86.0 ± 90.7 a | 35.4 ± 3.97 a | 23.2 ± 8.53 a | 38.1 ± 9.51 a |
| cyclo(L-Pro-L-Tyr) 2 | 347 ± 42.9 a | <LOD b | <LOD b | <LOD b | <LOD b |
| cyclo(L-Pro-L-Val) 2 | 269 ± 11.7 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Cytochalasin B | 75,000 ± 5890 a | 238 ± 305 b | 9.98 ± 4.06 b | 552 ± 857 b | <LOD b |
| Deoxygerfelin 1 | <LOD a | 1.83 ± 0.304 a | 7.36 ± 7.81 a | 2.26 ± 2.60 a | 3.41 ± 3.87 a |
| Dinactin | <LOD a | <LOD a | 1.51 ± 1.19 a | 0.186 ± 0.104 a | 1.37 ± 0.613 a |
| Emodin 1 | <LOD b | 0.830 ± 0.349 ab | 1.67 ± 1.11 ab | 1.17 ± 0.49 a | 2.07 ± 1.08 a b |
| Endocrocin | <LOD b | <LOD b | <LOD b | <LOD b | 49.5 ± 7.45 a |
| Enniatin B | <LOD b | <LOD b | <LOD b | 9.30 ± 0.0543 a | <LOD b |
| Fallacinol | <LOD a | <LOD a | <LOD a | 2.07 ± 2.76 a | <LOD a |
| Iso-Rhodoptilometrin | <LOD b | 1.98 ± 1.67 ab | 0.98 ± 0.69 b | 1.05 ± 0.792 ab | 6.92 ± 4.23 a |
| Isosulochrin 1 | <LOD a | <LOD a | 2.98 ± 3.73 a | <LOD a | 1.42 ± 0.60 a |
| Lecanoric acid 1 | <LOD a | 8.51 ± 3.38 a | 34.2 ± 47.1 a | 16.0 ± 14.3 a | 5.44 ± 4.13 a |
| Monactin | <LOD b | <LOD b | 19.8 ± 14.2 a | 12.4 ± 2.56 ab | 6.05 ± 0.88 ab |
| Nonactin 1 | <LOD a | 0.97 ± 1.27 a | 2.42 ± 1.55 a | 1.40 ± 1.30 a | 0.52 ± 0.05 a |
| Secalonic acid D | <LOD d | <LOD cd | 5.94 ± 2.96 b c d | 5.89 ± 3.93 b | 23.6 ± 6.92 a |
| Siccanol | <LOD b | <LOD b | 69.8 ± 17.7 b | 190 ± 77.1 a | <LOD b |
| Solanine 1 | <LOD b | 14,800 ± 20,800 ab | 31,000 ± 13,900 ab | 8890 ± 2700 ab | 44,300 ± 9400 a |
| Sulochrin 1 | <LOD b | <LOD b | <LOD b | <LOD b | 2.07 ± 0.68 a |
| Terragine 1 | <LOD a | 2400 ± 3020 a | 1130 ± 518 a | 718 ± 320 a | <LOD a |
| Tryptophol 1,2 | <LOD a | 69.9 ± 85.1 a | 12.7 ± 2.76 a | 18.5 ± 4.31 a | 18.3 ± 4.44 a |
| Violaceol I | <LOD a | <LOD a | 3460 ± 5550 a | 453 ± 606 a | 326 ± 279 a |
| Violaceol II 1 | <LOD a | <LOD a | 93.9 ± 113 a | 547 ± 10.5 a | <LOD a |
| Xanthoquinoidin A1 | <LOD a | 45.0 ± 57.3 a | <LOD a | <LOD a | <LOD a |
| Components | Concentration Mean ± SD [ng/g] | ||||
|---|---|---|---|---|---|
| C. coccodes Culture on MEA Plate (n = 3) | C. coccodes Culture on Potato (n = 3) | Potatoes Treated with Biological Preparations and Infected with C. coccodes | |||
| GB (n = 3) | LpB (n = 3) | MpB (n = 3) | |||
| Abscisic acid | <LOD a | <LOD a | <LOD a | <LOD a | 26.4 ± 17.1 a |
| Asterric acid | <LOD a | <LOD a | <LOD a | <LOD a | 98.7 ± 158 a |
| Beauvericin 1 | <LOD a | 1.15 ± 0.984 a | 11.5 ± 9.68 a | 13.0 ± 13.9 a | 3.85 ± 4.31 a |
| Bikaverin 1 | <LOD b | 23.7 ± 23.4 ab | 99.4 ± 49.8 a | 44.5 ± 40.1 ab | 14.6 ± 1.92 b |
| Brevianamid F 2 | 31.2 ± 2.81 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Chaconin | <LOD b | <LOD b | 5110 ± 1100 a | 3480 ± 618 a | 5450 ± 2230 a |
| Citreorosein | <LOD b | <LOD b | <LOD b | <LOD b | 2.10 ± 0.479 a |
| Cordycepin 1,2 | <LOD b | 58.1 ± 23.5 ab | 92.3 ± 61.3 a | 88.3 ± 4.15 a | 33.2 ± 5.96 ab |
| cyclo(L-Pro-L-Tyr) 2 | 490 ± 229 a | <LOD b | <LOD b | <LOD b | <LOD b |
| cyclo(L-Pro-L-Val) 2 | 297 ± 81.3 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Cytochalasin E 1 | 38,000 ± 12,800 a | <LOD b | <LOD b | <LOD b | <LOD b |
| Deoxygerfelin 1 | <LOD a | 1.55 ± 0.359 a | 12.0 ± 14.4 a | 3.88 ± 5.11 a | 3.06 ± 3.24 a |
| Dinactin | <LOD a | <LOD a | <LOD a | <LOD a | 1.78 ± 1.71 a |
| Emodin 1 | 6.13 ± 4.42 a | <LOD a | 8.66 ± 11.5 a | 0.47 ± 0.29 a | 5.45 ± 3.98 a |
| Endocrocin | 370 ± 18.1 a | <LOD b | <LOD b | 28.2 ± 15.3 b | 23.8 ± 19.5 b |
| Fallacinol | <LOD b | <LOD b | <LOD b | <LOD b | 1.33 ± 1.07 a |
| Fusaric acid | <LOD a | <LOD a | <LOD a | 4090 ± 5540 a | <LOD a |
| Iso-Rhodoptilometrin | 0.384 ± 0.181 a | 0.413 ± 0.561 a | 8.96 ± 11.2 a | 1.36 ± 1.32 a | 1.88 ± 0.49 a |
| Isosulochrin 1 | <LOD a | <LOD a | <LOD a | 6.33 ± 10.4 a | 10.5 ± 12.3 a |
| Lecanoric acid 1 | <LOD a | <LOD a | 31.1 ± 49.0 a | 13.40 ± 16.6 a | 27.6 ± 31.1 a |
| Monactin | <LOD a | 10.3 ± 11.3 a | <LOD a | <LOD a | 18.6 ± 14.8 a |
| Monocerin 1 | 1760 ± 487 a | 4.24 ± 5.32 b | 24.0 ± 9.74 b | 41.6 ± 33.3 b | 4.15 ± 3.62 b |
| Nonactin 1 | <LOD a | 1.11 ± 1.42 a | <LOD a | <LOD a | 1.59 ± 1.12 a |
| Rosellichalasin 1 | 458 ± 65.0 a | <LOD b | <LOD b | 11.0 ± 0.65 b | <LOD b |
| Sambutoxin | <LOD a | <LOD a | <LOD a | 0.0807 ± 0.113 a | <LOD a |
| Solanin 1 | <LOD a | 6140 ± 1690 a | 5250 ± 1870 a | 2950 ± 908 a | 9590 ± 9020 a |
| Sulochrin 1 | <LOD a | <LOD a | <LOD a | <LOD a | 11.1 ± 15.6 a |
| Terragine 1 | <LOD b | 780 ± 830 ab | 3110 ± 1740 a | 1430 ± 156 ab | 1060 ± 578 ab |
| Trichodimerol | <LOD b | <LOD b | <LOD b | 486 ± 298 a | 22.5 ± 18.9 b |
| Trichotetronine | <LOD b | <LOD b | <LOD b | 40.3 ± 9.79 a | <LOD b |
| Tryptophol 1,2 | <LOD c | 6.07 ± 2.16 ab | 10.4 ± 2.88 a | <LOD c | 5.52 ± 0.89 b |
| Violaceol I | <LOD a | <LOD a | 2330 ± 3680 a | 1090 ± 1650 a | 1820 ± 2090 a |
| Violaceol II 1 | <LOD a | <LOD a | <LOD a | 38.8 ± 43.2 a | 47.0 ± 45.6 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steglińska, A.; Sulyok, M.; Janas, R.; Grzesik, M.; Liszkowska, W.; Kręgiel, D.; Gutarowska, B. Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents. Int. J. Environ. Res. Public Health 2023, 20, 5221. https://doi.org/10.3390/ijerph20065221
Steglińska A, Sulyok M, Janas R, Grzesik M, Liszkowska W, Kręgiel D, Gutarowska B. Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents. International Journal of Environmental Research and Public Health. 2023; 20(6):5221. https://doi.org/10.3390/ijerph20065221
Chicago/Turabian StyleSteglińska, Aleksandra, Michael Sulyok, Regina Janas, Mieczysław Grzesik, Wiktoria Liszkowska, Dorota Kręgiel, and Beata Gutarowska. 2023. "Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents" International Journal of Environmental Research and Public Health 20, no. 6: 5221. https://doi.org/10.3390/ijerph20065221
APA StyleSteglińska, A., Sulyok, M., Janas, R., Grzesik, M., Liszkowska, W., Kręgiel, D., & Gutarowska, B. (2023). Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents. International Journal of Environmental Research and Public Health, 20(6), 5221. https://doi.org/10.3390/ijerph20065221