Hypothalamic Glial-to-Neuronal Signaling during Puberty: Influence of Alcohol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. EGF/TGFα Family of Growth Factors and Puberty
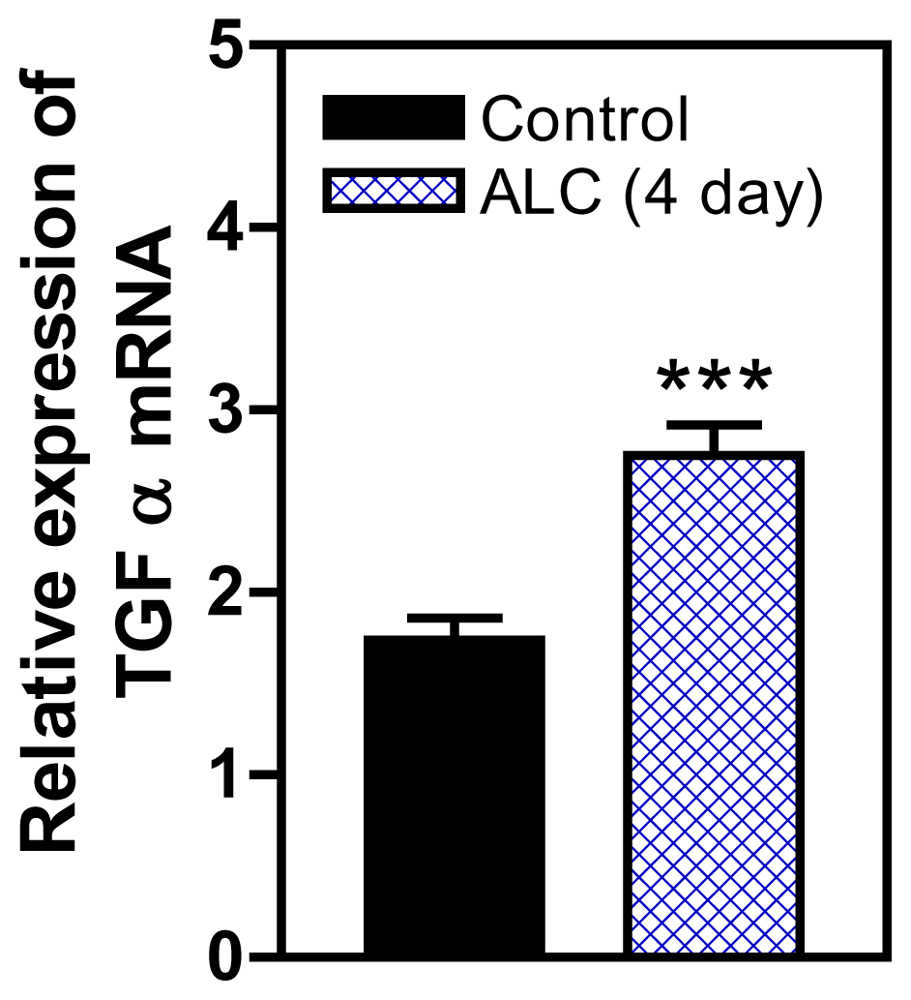
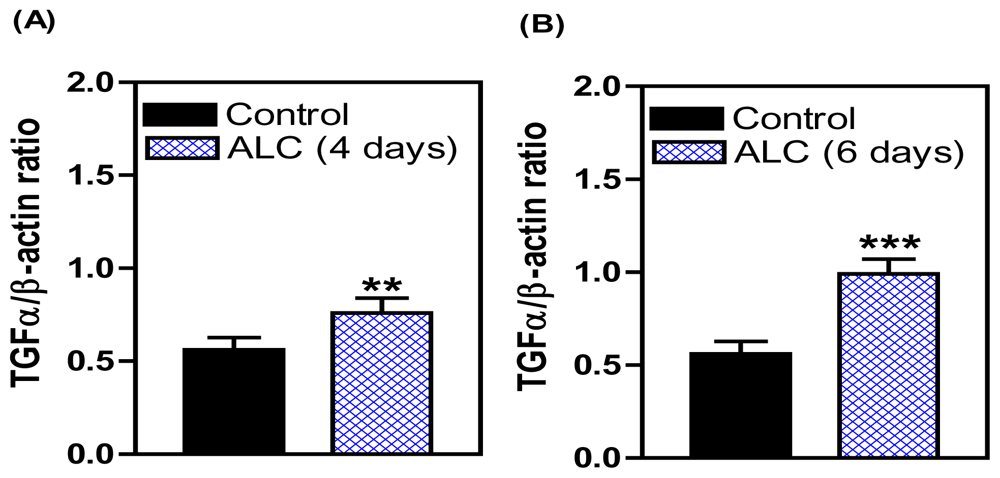
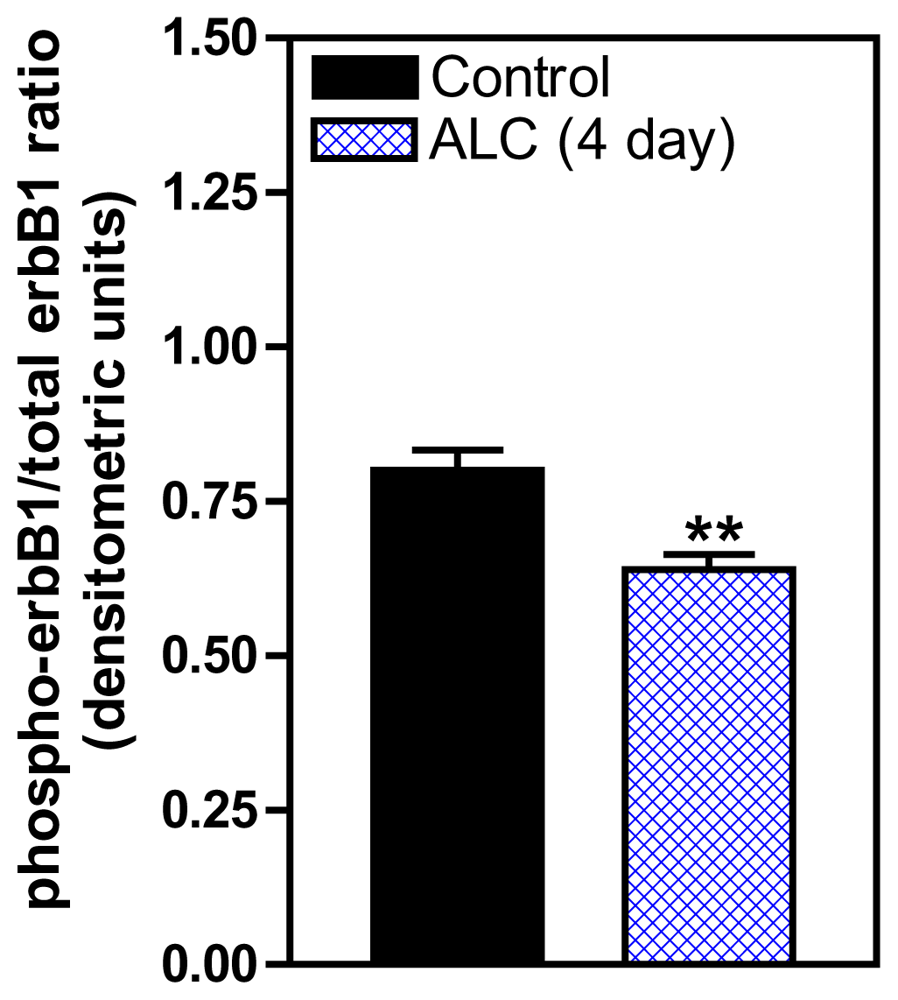
3. Effects of ALC on the TGFα/erbB1 Receptor/PGE2 Pathway
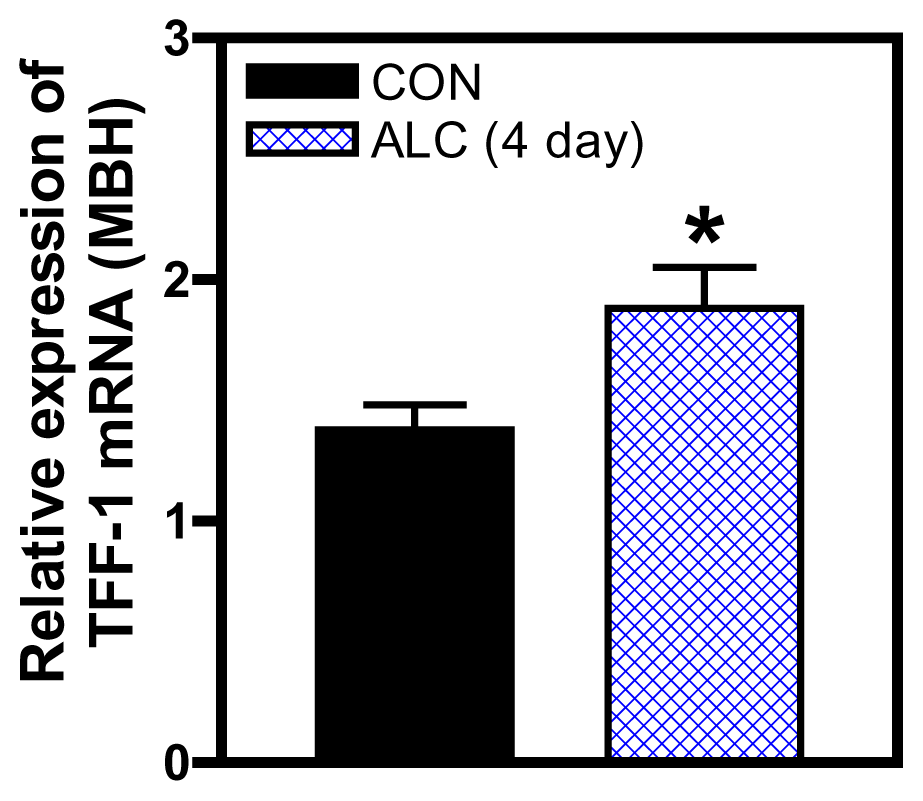
4. Adhesion/Signaling Genes Involved in Glial-Neuronal Communication during Puberty
5. Effects of ALC on Glial-Neuronal Adhesion and Signaling
6. Actions and Interactions of IGF-1 and ALC on Glial-Neuronal Adhesion and Signaling
7. Conclusions
Acknowledgements
References
- Brann, DW; Mahesh, VB. Excitatory amino acids: Function and significance in reproduction and neuroendocrine regulation. Front. Neuroendocrinol 1994, 15, 3–49. [Google Scholar]
- Ojeda, SR; Urbanski, HF. Puberty in the rat. In The Physiology of Reproduction, 2nd ed; Knobil, E, Neill, JD, Eds.; Raven Press: New York NY, USA, 1994; pp. 363–409. [Google Scholar]
- Crowley, WR; Parker, SL; Sahu, A; Kalra, SP. Interacting transmembrane signals regulating GnRH and LH secretion. In The Neurobiology of Puberty; Plant, TM, Lee, PA, Eds.; J Endocrinol: Bristal, UK, 1995; pp. 41–54. [Google Scholar]
- Ojeda, SR. The neurobiology of mammalian puberty: Has the contribution of glial cells been underestimated? J. NIH Res 1994, 6, 51–56. [Google Scholar]
- Terasawa, E. Hypothalamic control of the onset of puberty. Curr. Opin. Endocrinol. Diabetes 1999, 6, 44–49. [Google Scholar]
- Terasawa, E; Fernandez, DL. Neurobiological-mechanisms of the onset of puberty in primates. Endocr. Rev 2001, 22, 111–151. [Google Scholar]
- Claypool, LE; Kasuya, E; Saitoh, Y; Marzban, F; Terasawa, E. N-methyl-d,l-aspartate induces the release of LHRH in the prepubertal and pubertal female rhesus monkey as measured by in vivo push-pull perfusion in the stalk-median eminence. Endocrinology 2000, 141, 219–228. [Google Scholar]
- Urbanski, HF; Ojeda, SR. A role for N-methyl-d-aspartate (NMDA) receptors in the control of LH secretion and initiation of female puberty. Endocrinology 1990, 126, 1774–1776. [Google Scholar]
- Ojeda, SR; Urbanski, HF; Costa, ME; Hill, DF; Moholt-Siebert, M. Involvement of transforming growth factor alpha in the release of luteinizing hormone-releasing hormone from the developing female hypothalamus. Proc. Natl. Acad. Sci. USA 1990, 87, 9698–9702. [Google Scholar]
- Hiney, JK; Ojeda, SR; Dees, WL. Insulin-like growth factor (IGF-1) stimulates LHRH release from the prepubertal female median eminence in vitro. Neuroendocrinology 1991, 54, 420–423. [Google Scholar]
- Hiney, JK; Srivastava, VK; Nyberg, CL; Ojeda, SR; Dees, WL. Insulin-like growth factor-1 (IGF-1) of peripheral origin acts centrally to accelerate the initiation of female puberty. Endocrinology 1996, 137, 3717–3721. [Google Scholar]
- Navarro, CL; Castellano, JM; Fernandez-Fernandez, R; Barriero, ML; Roa, J; Sanhez-Criado, JE; Aguilar, E; Dieguez, C; Pinilla, L; Tena-Sempere, M. Developmental and hormonally regulated mRNA expression of KiSS-1 and its putative receptor, GPR54, in rat hypothalamus and potent LH-releasing activity of KiSS-1 peptide. Endocrinology 2004, 145, 4565–4574. [Google Scholar]
- Ojeda, SR; Lomniczi, A; Sandau, US. Glial-gonadotropin hormone (GnRH) neuron interactions in the median eminence and the control of GnRH secretion. J. Neuroendocrinol 2008, 20, 732–742. [Google Scholar]
- Ojeda, SR; Lomniczi, A; Sandau, U. Contribution of glial-neuronal interactions to the neuroendocrine control of female puberty. Eur. J. Neurosci 2010, 32, 2003–2010. [Google Scholar]
- Ma, YJ; Berg-von der Emde, K; Moholt-Siebert, M; Hill, DF; Ojeda, SR. Region specific regulation of transforming growth factor-α (TGFα) gene expression in astrocytes of the neuroendocrine brain. J. Neurosci 1994, 14, 5644–5651. [Google Scholar]
- Ma, YJ; Costa, ME; Ojeda, SR. Developmental expression of the genes encoding transforming growth factor alpha and its receptor in the hypothalamus of female rhesus macaques. Neuroendocrinology 1994, 60, 346–359. [Google Scholar]
- Tsai, PS; Werner, S. Basic fibroblast growth factor is a neurotropic factor in GT1 gonadotropin-releasing hormone neuronal cell lines. Endocrinology 1995, 136, 3831–3938. [Google Scholar]
- Voigt, P; Ma, YJ; Gonzales, D; Fahrenbach, WH; Wetsel, WC; Berg-Von der Emde, K; Hill, DF; Taylor, KG; Costa, ME; Seidah, NG; Ojeda, SR. Neural and glial mediated effects of growth factors acting via tyrosine kinase receptor on LHRH neurons. Endocrinology 1996, 137, 2593–2605. [Google Scholar]
- Prevot, V. Glial-neuronal-endothelial interactions are involved in the control of GnRH secretion. J. Neuroendocrinol 2002, 14, 247–255. [Google Scholar]
- Olson, BR. Effects of insulin-like growth factors I and II and insulin on the immortalized hypothalamic GT1-7 cell line. Neuroendocrinology 1995, 62, 155–165. [Google Scholar]
- Prevot, V; Cornea, A; Mungenast, A; Smiley, G; Ojeda, SR. Activation of erb-1 signaling in tanycytes of the median eminence stimulates transforming growth factor β1 release via prostaglandin E2 production and induces cell plasticity. J. Neurosci 2003, 23, 10622–10632. [Google Scholar]
- Rage, F; Lee, BJ; Ma, YJ; Ojeda, SR. Estradiol enhances prostaglandin E2 (PGE2) receptor gene expression in luteinizing hormone-releasing hormone (LHRH) neurons and facilitates the LHRH response to PGE2 by activating a glia-to-neuron signaling pathway. J. Neurosci 1997, 17, 9145–9156. [Google Scholar]
- Mungenast, AE; Ojeda, SR. Expression of three gene families encoding cell-cell communication molecules in the prepubertal nonhuman primate hypothalamus. J Neuroendocrinol 2005, 17, 208–219. [Google Scholar]
- Ojeda, SR; Dubay, C; Lomniczi, A; Kaidar, G; Matagne, V; Sandau, US; Dissen, GA. Gene networks and the neuroendocrine regulation of puberty. Mol. Cell. Endocrinol. 2010, 324, 3–11. [Google Scholar]
- Peles, E; Nativ, M; Campbell, PL; Sakuria, T; Martinez, R; Lev, S; Clary, DO; Schilling, J; Barnea, G; Plowman, GD; Gurmet, M; Schlessinger, J. The carbonic anhydrase domain of receptor tyrosine phosphatase ß is a functional ligand for the recognition molecule contactin. Cell 1995, 82, 251–260. [Google Scholar]
- Peles, E; Schlessinger, J; Grumet, M. Multi-ligand interactions with receptor-like protein tyrosine phosphatase β: implications for intercellular signaling. Trends Biochem Sci 1998, 2(3), 121–124. [Google Scholar]
- Pierre, K; Rougon, G; Allard, M; Bonhomme, R; Gennarini, G; Poulain, DA; Theodosis, DT. Regulated expression of the cell adhesion glucoprotein F3 in adult hypothalamic magnocellular neurons. J. Neurosci 1998, 18, 5333–5343. [Google Scholar]
- Dees, WL; Skelley, CW. The effects of ethanol during the onset of female puberty. Neuroendocrinology 1990, 51, 64–69. [Google Scholar]
- Srivastava, VK; Hiney, JK; Nyberg, CL; Dees, WL. Effect of ethanol on the synthesis of insulin-like growth factor-1(IGF-1) and the IGF-1 receptor in late prepubertal female rats: a correlation with serum IGF-1. Alc. Clin. Exp. Res 1995, 19, 1467–1473. [Google Scholar]
- Srivastava, VK; Hiney, JK; Dearth, RK; Dees, WL. Chronic effects of prepubertal ethanol administration on steroidogenic acute regulatory protein in the rat ovary. Alc. Clin. Exp. Res 2002, 26, 107–113. [Google Scholar]
- Emanuele, N; Ren, J; LaPaglia, N; Steiner, J. EtOH disrupts female mammalian puberty: age and opiate dependence. Endocrine 2002, 18, 247–254. [Google Scholar]
- Anderson, RA; Willis, BR; Oswald, C; Gupta, A; Zaneveld, L. Delayed male sexual maturation induced by chronic ethanol ingestion. Fed. Proc 1981, 40, 825–829. [Google Scholar]
- Ramaley, JA. The regulation of gonadotropin secretion in immature ethanol-treated male rats. J. Androl 1982, 3, 248–252. [Google Scholar]
- Dees, WL; Dissen, GA; Hiney, JK; Lara, F; Ojeda, SR. Alcohol ingestion inhibits the increased secretion of puberty-related hormones in the developing female rhesus monkey. Endocrinology 2000, 141, 1325–1331. [Google Scholar]
- Dissen, GA; Dearth, RK; Scott, HM; Ojeda, SR; Dees, WL. Alcohol alters prepubertal luteinizing hormone secretion in immature female rhesus monkeys by a hypothalamic action. Endocrinology 2004, 145, 4558–4564. [Google Scholar]
- Cardona-Gomez, GP; Doncarlos, L; Garcia-Segura, LM. Insulin-like growth factor I receptors and estrogen receptors colocalize in female rat brain. Neuroscience 2000, 99, 751–760. [Google Scholar]
- Ma, YJ; Berg-von der Emde, K; Rage, F; Wetsel, WC; Ojeda, SR. Hypothalamic astrocytes respond to transforming growth factor alpha with the secretion of neuroactive substances that stimulate the release of luteinizing hormone-releasing hormone. Endocrinology 1997, 138, 19–25. [Google Scholar]
- Ojeda, SR; Ma, YJ; Lee, BJ; Prevot, V. Glia to neuron signaling and the neuroendocrine control of female puberty. Recent Prog. Horm. Res 2000, 55, 197–224. [Google Scholar]
- Ma, YJ; Junier, MP; Costa, ME; Ojeda, SR. Transforming growth factor α (TGFα) gene expression in the hypothalamus is developmentally regulated and linked to sexual maturation. Neuron 1992, 9, 657–670. [Google Scholar]
- Yaish, P; Gazit, A; Gilon, C; Levitzki, A. Blocking of EGF-dependent cell proliferation by EGF receptor kinase inhibitors. Science 1988, 242, 933–935. [Google Scholar]
- Junier, MP; Ma, YJ; Costa, ME; Hoffman, G; Hill, DF; Ojeda, SR. Transforming growth factor α contributes to the mechanism by which hypothalamic injury induces precocious puberty. Proc. Natl. Acad. Sci. USA 1991, 88, 9743–9747. [Google Scholar]
- Junier, MP; Hill, DF; Costa, ME; Felder, S; Ojeda, SR. Hypothalamic lesions that induce female precocious puberty activate glial expression of the epidermal growth factor receptor gene: differential regulation of alternatively spliced transcripts. J. Neurosci 1993, 13, 703–713. [Google Scholar]
- Ma, YJ; Dissen, GA; Merlino, G; Coquelin, A; Ojeda, SR. Overexpression of a human transforming growth factor-α (TGFα) transgene reveals a dual antagonistic role of TGFα in female sexual development. Endocrinology 1994, 135, 1392–1400. [Google Scholar]
- Rage, F; Hill, DF; Sena-Esteves, M; Breakfield, XO; Coffey, RJ; Costa, ME; McCann, SM; Ojeda, SR. Targeting transforming growth factor α expression to discrete loci of the neuroendocrine brain induces female sexual precocity. Proc. Natl. Acad. Sci. USA 1997, 94, 2735–2740. [Google Scholar]
- Ojeda, SR; Ma, YJ. Glial-neuronal interactions in the neuroendocrine control of mammalian puberty: facilitatory effects of gonadal steroids. J. Neurobiol 1999, 40, 528–540. [Google Scholar]
- Dees, WL; Hiney, JK; Nyberg, CL. Effects of ethanol on the reproductive neuroendocrine axis of prepubertal and adult female rats. In The Reproductive Neuroendocrinology of Aging and Drug Abuse; Sarkar, DK, Barnes, CD, Eds.; CRC Press: Boca Raton, FL USA, 1995; pp. 301–334. [Google Scholar]
- Dees, WL; Srivastava, VK; Hiney, JK. Actions and interactions of alcohol and insulin-like growth factor-1 on female pubertal development. Alc. Clin. Exp. Res 2009, 11, 1847–1856. [Google Scholar]
- Ojeda, SR; Urbanski, HF; Katz, KH; Costa, ME; Conn, PM. Activation of two different but complementary biochemical pathways stimulates release of hypothalamic luteinizing hormone releasing hormone. Proc. Natl. Acad. Sci. USA 1986, 83, 4932–4936. [Google Scholar]
- Ojeda, SR; Urbanski, HF; Katz, KH; Costa, ME. Prostaglandin E2 releases luteinizing hormone releasing hormone for the female juvenile hypothalamus through Ca2+ dependent, calmodulin independent mechanism. Brain Res 1988, 441, 339–351. [Google Scholar]
- Hiney, JK; Dearth, RK; Srivastava, V; Rettori, V; Dees, WL. Actions of ethanol on epidermal growth factor receptor activated luteinizing hormone secretion. J. Stud. Alcohol 2003, 64, 809–816. [Google Scholar]
- Hiney, JK; Dees, WL. Ethanol inhibits LHRH release from the median eminence of prepubertal female rats in vitro: investigation of its action on norepinephrine and prostaglandin E2. Endocrinology 1991, 128, 1404–1408. [Google Scholar]
- Srivastava, VK; Hiney, JK; Dees, WL. Prepubertal ethanol exposure alters hypothalamic transforming growth factor α and erbB1 receptor signaling in the female rat. Alcohol 2011, 45, 173–181. [Google Scholar]
- Carpenter, G; Wahl, MI. The epidermal growth factor family. In Peptide Growth Factors and their Receptors; Sporn, MB, Roberts, AB, Eds.; Springer-Verlag: Berlin, Germany, 1990; pp. 69–171. [Google Scholar]
- Ebner, R; Derynck, R. Epidermal growth factor and transforming growth factor α: Differential intracellular routing and processing of ligand-receptor complexes. Cell Regul 1991, 2, 599–612. [Google Scholar]
- Tuma, DJ; Todero, SL; Barak-Bernihagen, M; Casey, CA; Sorrell, MF. Chronic ethanol ingestion impairs TGf α stimulated receptor autophosphorylation. Alcohol 1998, 15, 233–238. [Google Scholar]
- Wang, S; Feng, J; Ying, C; Wang, W. Time-dependent alteration of epidermal growth factor receptor in rat stomach by ethanol feeding. Toxicol. Lett 1997, 90, 115–123. [Google Scholar]
- Nyberg, CL; Hiney, JK; Minks, JE; Dees, WL. Ethanol alters N-methyl-dl-Aspartic acid-induced LH secretion of luteinizing hormone releasing hormone and the onset of puberty in the female rats. Neuroendocrinology 1993, 57, 863–868. [Google Scholar]
- Lomniczi, A; Mastronardi, CA; Faletti, AG; Seilicovich, A; De Laurentiis, A; McCann, SM; Rettori, V. Inhibitory pathways and the inhibition of luteinizing hormone-releasing hormone release by alcohol. Proc. Natl. Acad. Sci. USA 2000, 97, 2337–2342. [Google Scholar]
- Ojeda, SR; Hill, J; Hill, DF; Costa, ME; Tapia, V; Cornea, A; Ma, YJ. The Oct-2 POU domain gene in the neuroendocrine brain: A transcriptional regulator of mammalian puberty. Endocrinology 1999, 140, 3774–3789. [Google Scholar]
- Dees, WL; Srivastava, VK; Hiney, JK. Alcohol alters IGF-1 activated Oct-2 POU gene expression in the immature female hypothalamus. J. Stud. Alc 2005, 666, 35–45. [Google Scholar]
- Kim, HJ; Sohn, HJ; Ha, M; Han, JY; Kang, SS; Choi, WS; Cho, JG. Prepubertal chronic ethanol administration alters TTF-1 and Oct-2 expression in the hypothalamus of female rats. Mol. Brain Res 2005, 136, 262–266. [Google Scholar]
- Lee, BJ; Cho, GJ; Norgren, RB, Jr; Junier, MP; Hill, DF; Tapia, V; Costa, ME; Ojeda, SR. TTF-1, a homeodomain gene required for diencephalic morphogenesis, is postnatally expressed in the neuroendocrine brain in a developmentally regulated and cell-specific fashion. Mol. Cell Biol. 2001, 17, 107–126. [Google Scholar]
- Peles, E; Native, M; Lustig, M; Grumet, M; Schilling, J; Martinez, R; Plowman, GD; Schlissinger, J. Identification of a novel contactin-associated transmembrane receptor with multiple domains implicated in protein-protein interactions. EMBO J 1997, 16, 978–988. [Google Scholar]
- Lomniczi, A; Ojeda, SR. A role for glial cells of the neuroendocrine brain in the central control of female sexual development. In Astrocytes in Pathophysiology of the Nervous System; Purpura, V, Haydon, P, Eds.; Springer: New York, NY, USA, 2009; pp. 487–510. [Google Scholar]
- Sandau, US; Mungenast, AE; McCarthy, J; Biederer, T; Corfas, G; Ojeda, SR. The synaptic cell adhesion molecule, SynCAM1, mediates astrocyte-to astrocyte and astrocyte-to-GnRH neuron adhesiveness in the mouse hypothalamus. Endocrinology 2011, 152, 2353–2363. [Google Scholar]
- Ma, YJ; Hill, DF; Creswick, KE; Costa, ME; Cornea, A; Lioubin, MN; Plowman, GD; Ojeda, SR. Neuregulins signaling via a glial erb-2-erb-4 receptor complex contribute to the neuroendocrine control of mammalian sexual development. J. Neurosci 1999, 19, 9913–9927. [Google Scholar]
- Prevot, V; Rio, C; Cho, GJ; Lomniczi, A; Heger, S; Neville, CM; Rosenthal, NA; Ojeda, SR; Corfas, G. Normal female sexual development requires neuregulin-erbB receptor signaling in hypothalamic astrocytes. J. Neurosci 2003, 23, 230–239. [Google Scholar]
- Sandau, US; Mungenast, AE; Alderman, Z; Pablo Sardi, S; Fogel, AI; Taylor, B; Parent, A; Biederer, T; Corfas, G; Ojeda, SR. SynCAM1, a synaptic adhesion molecule, is expressed in astrocytes and contributes to erbB4 receptor-mediated control of female sexual development. Endocrinology 2011, 152, 2364–2376. [Google Scholar]
- King, JC; Letourneau, RL. Luteinizing hormone releasing hormone terminals in the median eminence of rats undergo dynamic changes after gonadectomy, as revealed by electron microscopic image analysis. Endocrinology 1994, 134, 1340–1351. [Google Scholar]
- Prevot, V; Bouret, S; Croix, D; Alonso, G; Jennes, L; Mitchell, V; Routtenberg, A; Beayvillain, JC. Growth associated protein-43 messenger ribonucleic acid expression in gonadotropin releasing hormone neurons during the rat estrous cycle. Endocrinology 2000, 141, 1648–1657. [Google Scholar]
- Theodosis, DT; Poulain, DA. Neuronal-glial and synaptic plasticity of the adult oxytocinergic system. Ann. N. Y. Acad. Sci 1992, 652, 303–325. [Google Scholar]
- Witkin, JW; O’Sullivan, H; Ferin, M. Glial ensheathment of GnRH neurons in prepubertal female rhesus macaques. J. Neuroendocrinol 1995, 7, 665–671. [Google Scholar]
- Srivastava, VK; Hiney, JK; Dees, WL. Hypothalamic actions and interactions of alcohol and IGF-1 on the expression of glial receptor protein tyrosine phosphatase-ß during female pubertal development. Alc Clin Exp Res 2011. [Google Scholar] [CrossRef]
- Wilson, ME. Premature elevation in serum insulin-like growth factor-1 advances first ovulation in rhesus monkeys. J. Endocrinol 1998, 158, 247–257. [Google Scholar]
- Handelsman, DJ; Spaliviero, JA; Scott, CD; Baxter, RC. Hormonal regulation of the peripubertal stage of insulin-like growth factor-I in the rat. Endocrinology 1987, 120, 491–496. [Google Scholar]
- Copeland, KC; Kuehl, TJ; Castracane, VD. Pubertal endocrinology of the baboon: elevated somatomedin-C/insulin-like growth factor I at puberty. J. Clin. Endocrinol. Metab 1982, 55, 1198–1201. [Google Scholar]
- Bondy, C; Werner, H; Roberts, CT; Leroith, D. Cellular pattern of type-I insulin-like growth factor receptor gene expression during maturation of the rat brain: comparison with insulin-like growth factors I and II. Neuroscience 1992, 46, 909–923. [Google Scholar]
- Vendola, K; Zhou, J; Wang, J; Bondy, CA. Androgens promote insulin-like growth factor-1 and insulin-like growth factor-1 receptor gene expression in the primate ovary. Hum. Reprod 1999, 14, 2328–2332. [Google Scholar]
- DiVall, SA; Williams, TR; Carver, SE; Koch, L; Bruning, JC; Kahn, CR; Wondisford, F; Radovick, S; Wolfe, A. Divergent roles of growth factors in the GnRH regulation of puberty in mice. J. Clin. Invest 2010, 120, 2900–2909. [Google Scholar]
- Parent, AS; Mungenast, AE; Lomniczi, A; Sandau, US; Peles, E; Bosch, MA; Ronnekliev, OK; Ojeda, SR. A contactin receptor-like protein tyrosine phosphatase beta complex mediates adhesive communication between astroglial cell and gonadotropin releasing hormone neurons. J. Neuroendocrinol 2007, 19, 847–859. [Google Scholar]
- Duenas, M; Luquin, S; Chowen, JA; Torres-Aleman, I; Naftolin, F; Garcia-Segura, LM. Gonadal hormone regulation of insulin-like growth factor-I like immunoreactivity in hypothalamic astroglia of developing and adult rats. Neuroendocrinology 1994, 59, 528–538. [Google Scholar]
- Srivastava, VK; Hiney, JK; Dees, WL. Short term alcohol administration alters KiSS-1 gene expression in the reproductive hypothalamus of prepubertal female rats. Alcohol. Clin. Exp. Res 2009, 33, 1605–1614. [Google Scholar]
- Hiney, JK; Srivastava, VK; Dees, WL. Insulin-like growth factor-1 stimulates hypothalamic KiSS-1 gene expression by activating Akt: Effect of alcohol. Neuroscience 2010, 166, 625–632. [Google Scholar]











© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Srivastava, V.K.; Hiney, J.K.; Les Dees, W. Hypothalamic Glial-to-Neuronal Signaling during Puberty: Influence of Alcohol. Int. J. Environ. Res. Public Health 2011, 8, 2876-2894. https://doi.org/10.3390/ijerph8072894
Srivastava VK, Hiney JK, Les Dees W. Hypothalamic Glial-to-Neuronal Signaling during Puberty: Influence of Alcohol. International Journal of Environmental Research and Public Health. 2011; 8(7):2876-2894. https://doi.org/10.3390/ijerph8072894
Chicago/Turabian StyleSrivastava, Vinod K., Jill K. Hiney, and W. Les Dees. 2011. "Hypothalamic Glial-to-Neuronal Signaling during Puberty: Influence of Alcohol" International Journal of Environmental Research and Public Health 8, no. 7: 2876-2894. https://doi.org/10.3390/ijerph8072894
APA StyleSrivastava, V. K., Hiney, J. K., & Les Dees, W. (2011). Hypothalamic Glial-to-Neuronal Signaling during Puberty: Influence of Alcohol. International Journal of Environmental Research and Public Health, 8(7), 2876-2894. https://doi.org/10.3390/ijerph8072894