Forest Dynamics and Their Phenological Response to Climate Warming in the Khingan Mountains, Northeastern China


Abstract
:
1. Introduction
2. Materials and Methods
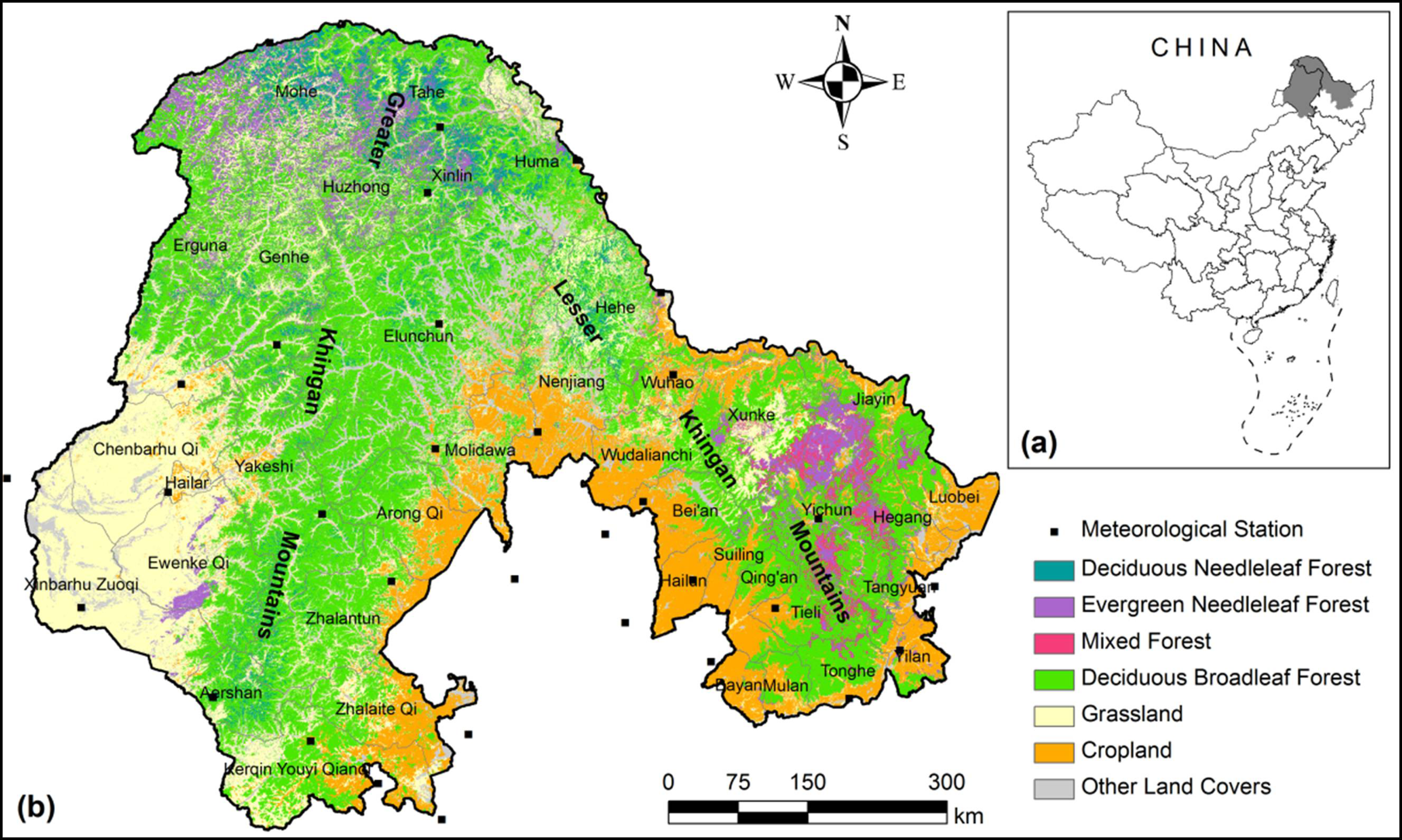
2.1. Study Area

2.2. MODIS Imagery
2.3. Meteorological Data
2.4. Phenological Variables
2.5. Statistical Analysis
3. Results
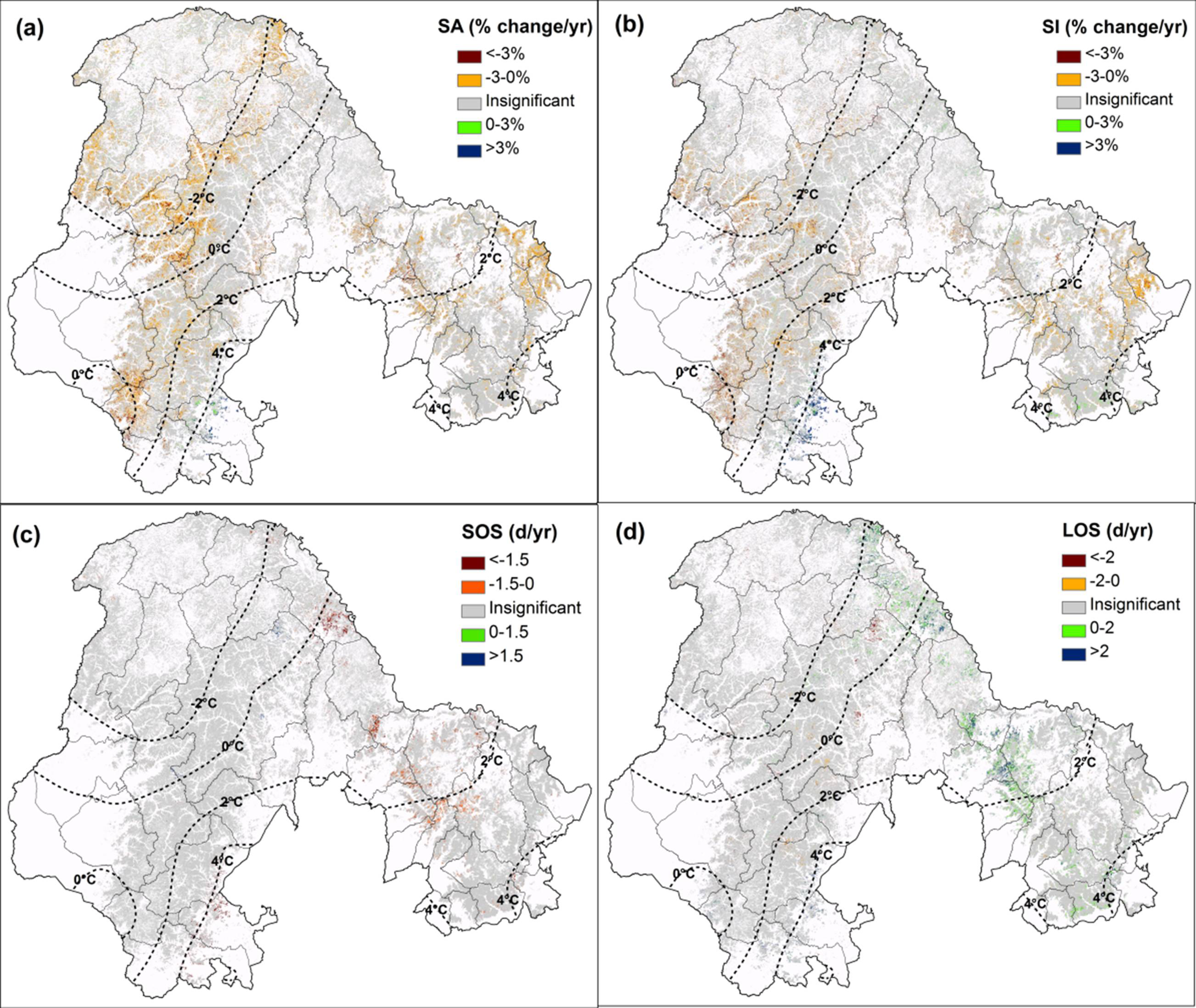
3.1. The Vegetation and Phenology Change of DBF


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Negative trends | Positive trends | Total | |||
|---|---|---|---|---|---|---|
| SA | 13.38 a | 75.32 | 0.93 a | 24.68 | 14.31 a | 100 |
| SI | 7.99 a | 66.39 | 1.86 a | 33.61 | 9.85 a | 100 |
| SOS | 1.93 a | 70.81 | 0.3 a | 29.19 | 2.23 a | 100 |
| LOS | 1.16 a | 39.99 | 3.74 a | 60.01 | 4.90 a | 100 |
3.2. The Impact of Temperature on Phenology Change
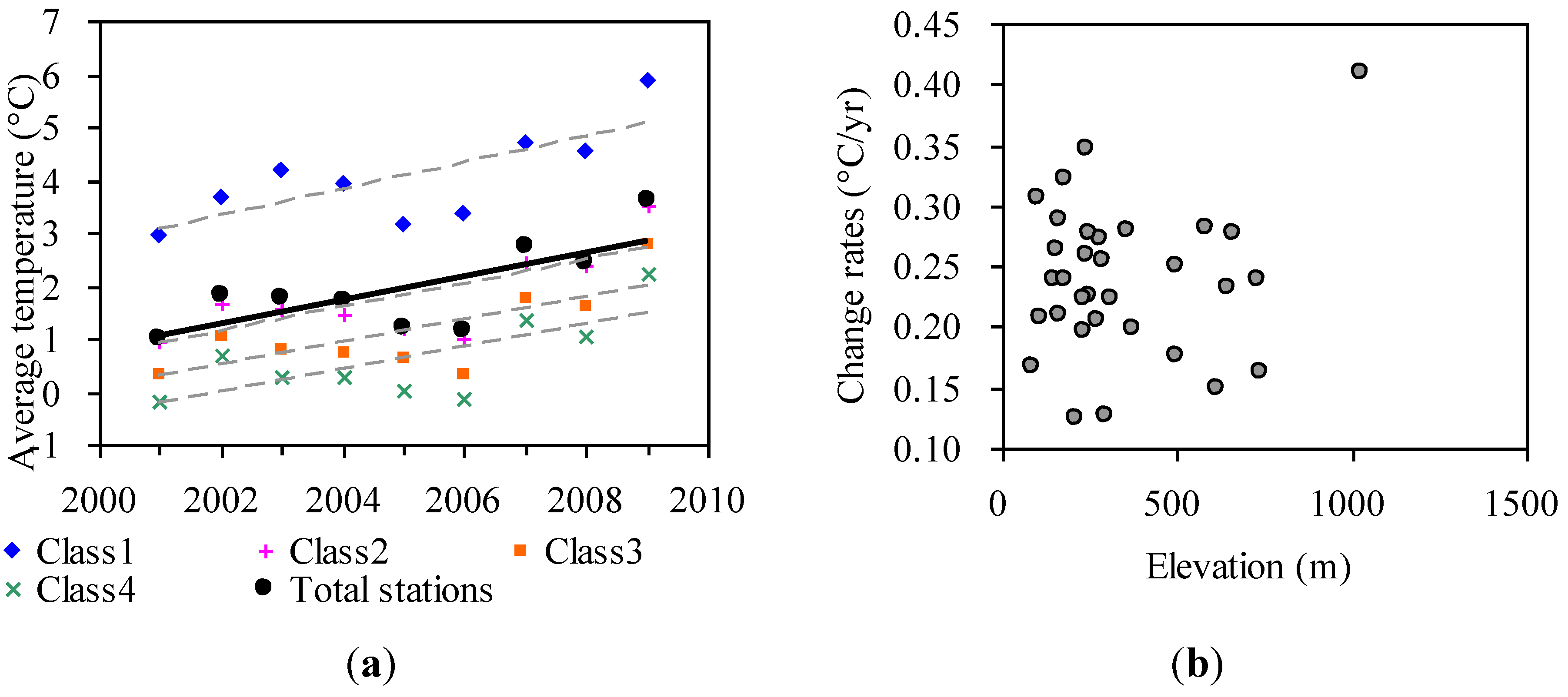
3.2.1. The Temperature Change in Khingan Mountains
| Classes | Slope | R2 | p | Number of stations | Elevation (m) | Latitude |
|---|---|---|---|---|---|---|
| Class1 | 0.245 | 0.55 | 0.022 a | 15 | 0–400 | 45°N–49°N |
| Class2 | 0.228 | 0.56 | 0.021 a | 8 | 0–400 | 49°N–54°N |
| Class3 | 0.215 | 0.53 | 0.025 a | 4 | 400–1,200 | 45°N–49°N |
| Class4 | 0.209 | 0.51 | 0.032 a | 5 | 400–1,200 | 49°N–54°N |
| Total stations | 0.227 | 0.53 | 0.027 a | 32 | 0–1,200 | 45°N–54°N |

3.2.2. Statistical Patterns of Relationships between Phenology and Temperature
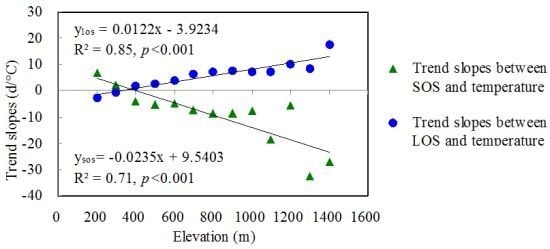
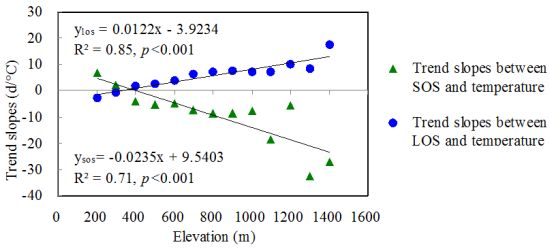
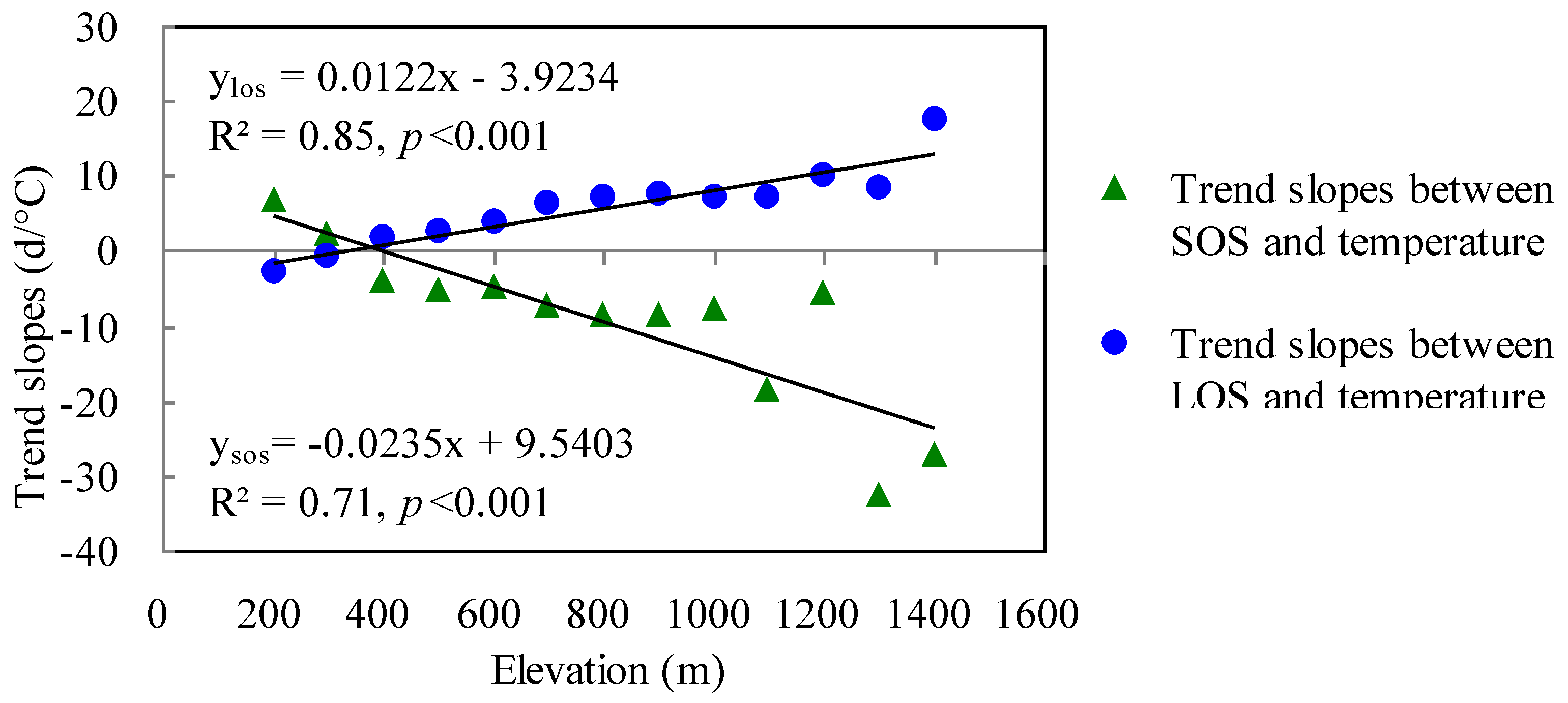
3.2.3. The Effects of Temperature Gradients in Relation to Elevation

4. Discussion and Conclusions
Acknowledgments
Conflict of Interest
References
- Hidalgo, P.J.; Galan, C.; Dominguez, E. Male phenology of three species of Cupressus: Correlation with airborne pollen. Trees-struct. Funct. 2003, 17, 336–344. [Google Scholar]
- Estrella, N.; Menzel, A.; Kramer, U.; Behrendt, H. Integration of flowering dates in phenology and pollen counts in aerobiology: Analysis of their spatial and temporal coherence in Germany (1992-1999). Int. J. Biometeorol. 2006, 51, 49–59. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Y.S. De(re)forestation and climate warming in subarctic China. Appl. Geogr. 2012, 32, 281–290. [Google Scholar] [CrossRef]
- Grasso, M.; Manera, M.; Chiabai, A.; Markandya, A. The health effects of climate change: A survey of recent quantitative research. Int. J. Envrion. Res. Public Health 2012, 9, 1523–1547. [Google Scholar] [CrossRef]
- Huynen, M.M.T.E.; Martens, P.; Schram, D.; Weijenberg, M.P.; Kunst, A.E. The impact of heat waves and cold spells on mortality rates in the Dutch population. Environ. Health Persp. 2001, 109, 463–470. [Google Scholar] [CrossRef]
- Frei, T.; Gassner, E. Climate change and its impact on birch pollen quantities and the start of the pollen season an example from Switzerland for the period 1969-2006. Int. J. Biometeorol. 2008, 52, 667–674. [Google Scholar] [CrossRef]
- Barrios, J.M.; Verstraeten, W.W.; Maes, P.; Clement, J.; Aerts, J.M.; Haredasht, S.A.; Wambacq, J.; Lagrou, K.; Ducoffre, G.; Van Ranst, M.; et al. Satellite derived forest phenology and its relation with Nephropathia Epidemica in Belgium. Int. J. Environ. Res. Public Health 2010, 7, 2486–2500. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Ciais, P.; Li, J.; Friedlingstein, P.; Koven, C.; Chen, A. Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. PNAS 2011, 108, 1240–1245. [Google Scholar]
- Piao, S.L.; Cui, M.D.; Chen, A.P.; Wang, X.H.; Ciais, P.; Liu, J.; Tang, Y.H. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agr. Forest. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Chen, X.Q.; Pan, W.F. Relationships among phenological growing season, time-integrated normalized difference vegetation index and climate forcing in the temperate region of eastern China. Int. J. Climatol. 2002, 22, 1781–1792. [Google Scholar] [CrossRef]
- Avolio, E.; Orlandi, F.; Bellecci, C.; Fornaciari, M.; Federico, S. Assessment of the impact of climate change on the olive flowering in Calabria (southern Italy). Theor. Appl. Climatol. 2012, 107, 531–540. [Google Scholar] [CrossRef]
- Zhou, L.M.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. 2001, 106, 20069–20083. [Google Scholar]
- Badeck, F.W.; Bondeau, A.; Bottcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Vitasse, Y.; Delzon, S.; Dufrene, E.; Pontailler, J.Y.; Louvet, J.M.; Kremer, A.; Michalet, R. Leaf phenology sensitivity to temperature in European trees: Do within-species populations exhibit similar responses? Agr. Forest. Meteorol. 2009, 149, 735–744. [Google Scholar] [CrossRef]
- Zheng, J. Relationships of the distribution of schistosomiasis and elevation in highland moutains in Yunnan. Strait J. Prev. Med. 2000, 6, 17–18. (in Chinese). [Google Scholar]
- Lu, Z.; Broker, M.; Liang, G.D. Tick-borne encephalitis in mainland China. Vector-Borne Zoonotic Dis. 2008, 8, 713–720. [Google Scholar] [CrossRef]
- Bi, W.M.; Deng, H.P.; Bu, X.Y. Studies on regionalization of natural foci of tick-borne encephalitis. J. Capital Norm. Univ. 1997, 18, 100–107. (in Chinese). [Google Scholar]
- Zhang, J.; Chen, S.; Zhao, X. Spatial divergency of temperature change during 1951-2000 in China and its correlation with global climate change. J. Arid Land Resour. Environ. 2006, 20, 1–6. [Google Scholar] [CrossRef]
- Cai, H.Y.; Zhang, S.W.; Bu, K.; Yang, J.C.; Chang, L.P. Integrating geographical data and phenological characteristics derived from MODIS data for improving land cover mapping. J. Geogr. Sci. 2011, 21, 705–718. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Sirikul, N. Comparison of MODIS Vegetation Indices with Biophysical and Flux Measurement. Ph.D Dissertation, The University of Arizona, Tucson, AZ, USA, 2006. [Google Scholar]
- Labus, M.P.; Nielsen, G.A.; Lawrence, R.L.; Engel, R.; Long, D.S. Wheat yield estimates using multi-temporal NDVI satellite imagery. Int. J. Remote. Sens. 2002, 23, 4169–4180. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP estimates from NDVI for the Central Grassland Region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, D. Monitoring forest phenophases of Northeast China based on MODIS NDVI data. Resour. Sci. 2006, 28, 111–117. [Google Scholar]
- Jönsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remot. Sen. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Eklundh, L.; Jönsson, P. TIMESAT 3.0 Software Mannual; Lund University: Lund, Sweden, 2009. [Google Scholar]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jönsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982-2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Olsson, L.; Eklundh, L.; Ardo, J. A recent greening of the Sahel—Trends, patterns and potential causes. J. Arid Environ. 2005, 63, 556–566. [Google Scholar] [CrossRef]
- Dong, J.H.; Zhu, J.H.; Yin, F.R. Epidemiologic investigations of tick-borne encephalitis and lyme disease in Great Khingan Mountains in Inner Mongolia. Chin. Prev. Med. 2007, 8, 718–719. [Google Scholar]
- Guo, Z.X. Research on Vegetation Changing and Reaction to Climate in Northeast China from 1982 to 2003; Northeast Institute of Geography and Agroecology: Changchun, China, 2008. [Google Scholar]
- Sun, F.H.; Yang, S.Y.; Chen, P.S. Climatic warming-drying trend in Northeastern China during the last 44 years and its effects. Chin. J. Ecol. 2005, 24, 751–755. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar]
- Wang, J.; Ji, L.Z.; Khomutova, M. Relationships between forest insect pest occurrence area and meteorological factors in Great Xing’an Mountains of Heilongjiang Province, Northeast China. Chin. J. Ecol. 2007, 26, 673–677. [Google Scholar]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659–659. [Google Scholar] [CrossRef]
- Tucker, C.J.; Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Myneni, R.B.; Taylor, M.G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int. J. Biometeorol. 2001, 45, 184–190. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Global. Biogeochem. Cy. 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Chen, X.Q.; Hu, B.; Yu, R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Global. Change. Biol. 2005, 11, 1118–1130. [Google Scholar] [CrossRef]
- Klemola, T.; Pettersen, T.; Stenseth, N.C. Trophic interactions in population cycles of voles and lemmings: A model-based synthesis. Adv. Ecol. Res. 2003, 33, 75–160. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cai, H.; Zhang, S.; Yang, X. Forest Dynamics and Their Phenological Response to Climate Warming in the Khingan Mountains, Northeastern China. Int. J. Environ. Res. Public Health 2012, 9, 3943-3953. https://doi.org/10.3390/ijerph9113943
Cai H, Zhang S, Yang X. Forest Dynamics and Their Phenological Response to Climate Warming in the Khingan Mountains, Northeastern China. International Journal of Environmental Research and Public Health. 2012; 9(11):3943-3953. https://doi.org/10.3390/ijerph9113943
Chicago/Turabian StyleCai, Hongyan, Shuwen Zhang, and Xiaohuan Yang. 2012. "Forest Dynamics and Their Phenological Response to Climate Warming in the Khingan Mountains, Northeastern China" International Journal of Environmental Research and Public Health 9, no. 11: 3943-3953. https://doi.org/10.3390/ijerph9113943
APA StyleCai, H., Zhang, S., & Yang, X. (2012). Forest Dynamics and Their Phenological Response to Climate Warming in the Khingan Mountains, Northeastern China. International Journal of Environmental Research and Public Health, 9(11), 3943-3953. https://doi.org/10.3390/ijerph9113943