Malignant Transformation of Rat Kidney Induced by Environmental Substances and Estrogen

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Experimental Designs
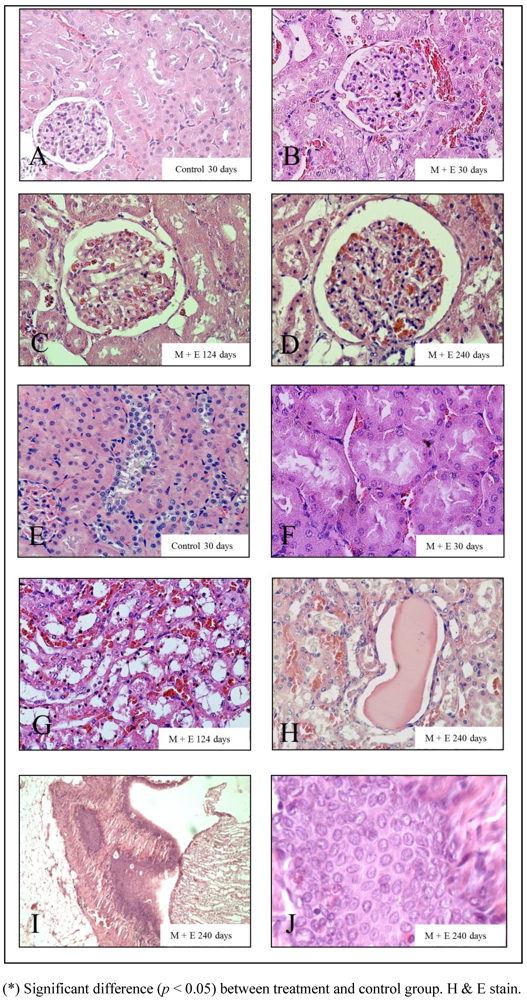
2.2. Morphological Studies
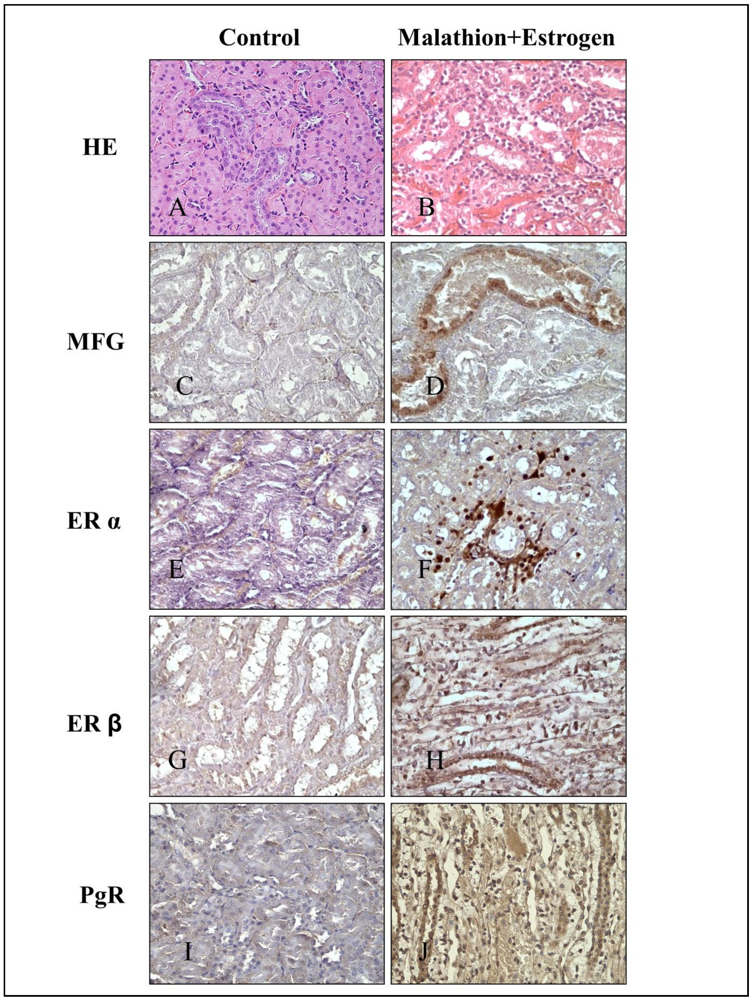
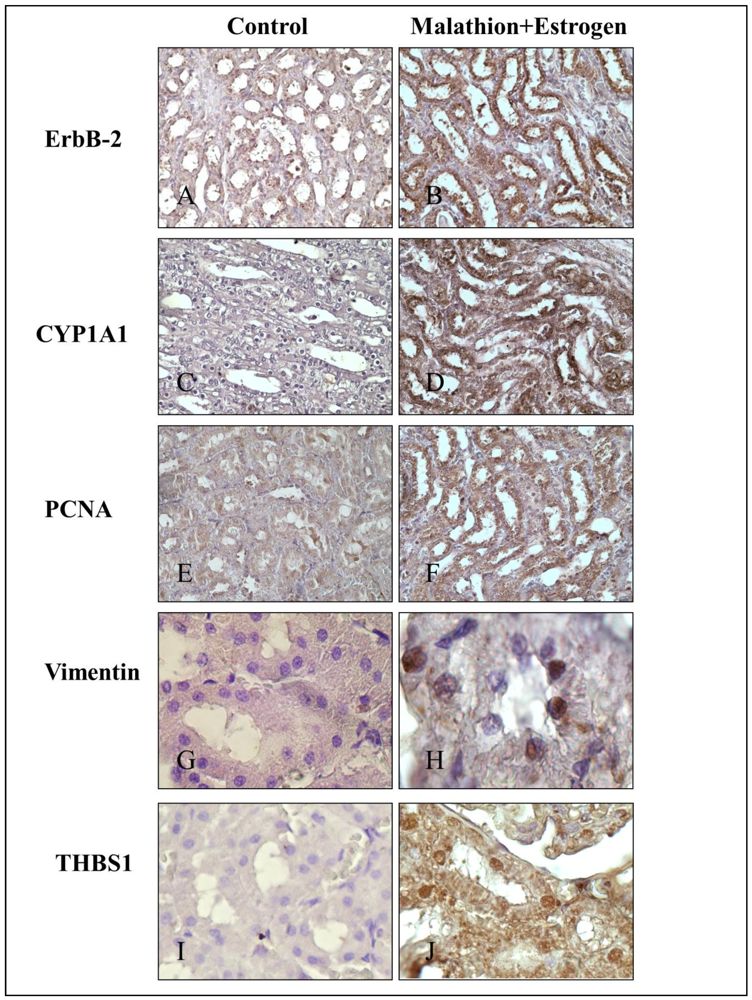
2.3. Immunocytochemical Staining
2.4. Statistical Analysis
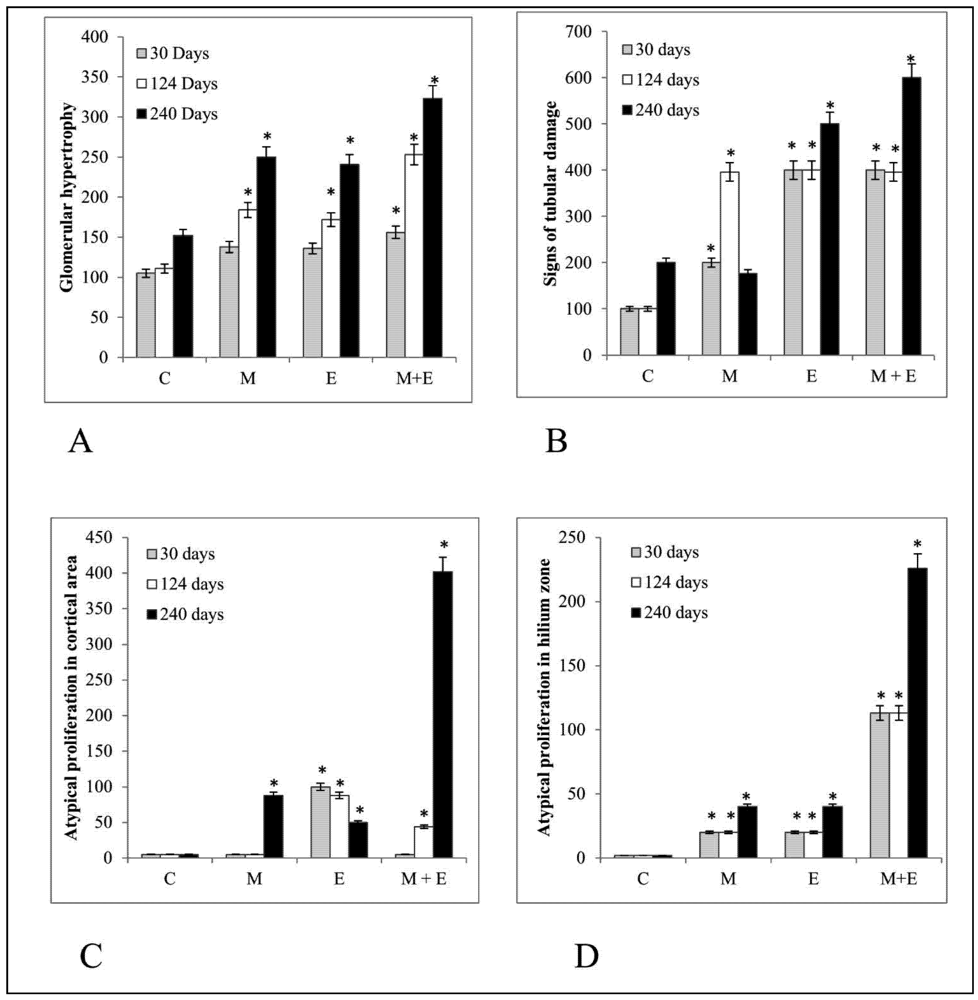
3. Results




4. Discussion
5. Conclusions
Acknowledgements
Conflict of Interest
References and Notes
- Goldsworthy, T.L.; Butterworth, B.E.; Maronpot, R.R. Concepts, labeling procedures, and design of cell proliferation studies relating to carcinogenesis. Environ. Health Perspect. 1993, 101 Suppl 5, 59–65. [Google Scholar]
- Bertram, J.S. The molecular biology of cancer. Mol. Aspects Med. 2000, 21, 167–223. [Google Scholar]
- Echiburu-Chau, C.; Calaf, G.M. Rat lung cancer induced by malathion and estrogen. Int. J. Oncol. 2008, 33, 603–611. [Google Scholar]
- Calaf, G.M.; Garrido, F. Catechol estrogens as biomarkers for mammary gland cancer. Int. J. Oncol. 2011, 39, 177–183. [Google Scholar]
- Edwards, J.W.; Lee, S.-G.; Heath, L.M.; Pisaniello, D.L. Worker exposure and a risk assessment of malathion and fenthion used in the control of Mediterranean fruit fly in South Australia. Environ. Res. 2007, 103, 38–45. [Google Scholar]
- Alavanja, M.C.; Samanic, C.; Dosemeci, M.; Lubin, J.; Tarone, R.; Lynch, C.F.; Knott, C.; Thomas, K.; Hoppin, J.A.; Barker, J.; Coble, J.; Sandler, D.P.; Blair, A. Use of agricultural pesticides and prostate cancer risk in the Agricultural Health Study cohort. Am. J. Epidemiol. 2003, 157, 800–814. [Google Scholar]
- Yoder, J.; Watson, M.; Benson, W.W. Lymphocyte chromosome analysis of agricultural workers during extensive occupational exposure to pesticides. Mutat. Res. 1973, 21, 335–340. [Google Scholar]
- Fenske, R.A.; Lu, C.; Barr, D.; Needham, L. Children’s exposure to chlorpyrifos and parathion in an agricultural community in central Washington State. Environ. Health Perspect. 2002, 110, 549–553. [Google Scholar]
- Centers for Disease Control and Prevention (CDC), Second National Report on Human Exposure to Environmental Chemicals; NCEH Publication No. 03-0022; CDC: Atlanta, GA, USA, 2003.
- Berkman, C.E.; Thompson, C.M.; Perrin, S.R. Synthesis, absolute configuration, and analysis of malathion, malaoxon, and isomalathion enantiomers. Chem. Res. Toxicol. 1993, 6, 718–723. [Google Scholar] [CrossRef]
- Aldridge, W.N.; Miles, J.W.; Mount, D.L.; Verschoyle, R.D. The toxicological properties of impurities in malathion. Arch. Toxicol. 1979, 42, 95–106. [Google Scholar]
- Durak, D.; Uzun, F.G.; Kalender, S.; Ogutcu, A.; Uzunhisarcikli, M.; Kalender, Y. Malathion-induced oxidative stress in human erythrocytes and the protective effect of vitamins C and E in vitro. Environ. Toxicol. 2009, 24, 235–242. [Google Scholar] [CrossRef]
- Al-Attar, A.M. Physiological and histopathological investigations on the effects of alpha-lipoic acid in rats exposed to malathion. J. Biomed. Biotechnol. 2010. [Google Scholar]
- Raymond, A.; Ensslin, M.A.; Shur, B.D. SED1/MFG-E8: A bi-motif protein that orchestrates diverse cellular interactions. J. Cell Biochem. 2009, 106, 957–966. [Google Scholar]
- Larocca, D.; Peterson, J.A.; Urrea, R.; Kuniyoshi, J.; Bistrain, A.M.; Ceriani, R.L. A Mr 46,000 human milk fat globule protein that is highly expressed in human breast tumors contains factor VIII-like domains. Cancer Res. 1991, 51, 4994–4998. [Google Scholar]
- Zurawa-Janicka, D.; Kobiela, J.; Stefaniak, T.; Wozniak, A.; Narkiewicz, J.; Wozniak, M.; Limon, J.; Lipinska, B. Changes in expression of serine proteases HtrA1 and HtrA2 during estrogen-induced oxidative stress and nephrocarcinogenesis in male Syrian hamster. Acta Biochim. Pol. 2008, 55, 9–19. [Google Scholar]
- Liehr, J.G. Is estradiol a genotoxic mutagenic carcinogen? Endocr. Rev. 2000, 21, 40–54. [Google Scholar] [CrossRef]
- Henderson, B.E.; Feigelson, H.S. Hormonal carcinogenesis. Carcinogenesis 2000, 21, 427–433. [Google Scholar]
- Fernandez, S.V.; Russo, J. Estrogen and xenoestrogens in breast cancer. Toxicol. Pathol. 2010, 38, 110–122. [Google Scholar]
- Harris, J.R.; Lippman, M.E.; Veronesi, U.; Willett, W. Breast cancer (1). N. Engl. J. Med. 1992, 327, 319–328. [Google Scholar]
- Liehr, J.G. Genotoxic effects of estrogens. Mutat. Res. 1990, 238, 269–276. [Google Scholar]
- Duong, B.N.; Elliott, S.; Frigo, D.E.; Melnik, L.I.; Vanhoy, L.; Tomchuck, S.; Lebeau, H.P.; David, O.; Beckman, B.S.; Alam, J.; Bratton, M.R.; McLachlan, J.A.; Burow, M.E. AKT regulation of estrogen receptor beta transcriptional activity in breast cancer. Cancer Res. 2006, 66, 8373–8381. [Google Scholar]
- Leung, Y.K.; Mak, P.; Hassan, S.; Ho, S.M. Estrogen receptor (ER)-beta isoforms: A key to understanding ER-beta signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 13162–13167. [Google Scholar]
- Scardino, A.; Alves, P.; Gross, D.A.; Tourdot, S.; Graff-Dubois, S.; Angevin, E.; Firat, H.; Chouaib, S.; Lemonnier, F.; Nadler, L.M.; Cardoso, A.A.; Kosmatopoulos, K. Identification of HER-2/neu immunogenic epitopes presented by renal cell carcinoma and other human epithelial tumors. Eur. J. Immunol. 2001, 31, 3261–3270. [Google Scholar]
- Goebel, S.U.; Iwamoto, M.; Raffeld, M.; Gibril, F.; Hou, W.; Serrano, J.; Jensen, R.T. Her-2/neu expression and gene amplification in gastrinomas: Correlations with tumor biology, growth, and aggressivenes. Cancer Res. 2002, 62, 3702–3710. [Google Scholar]
- Rintala, S.; Tammela, T.L.; Tuimala, R. CYP1A1 activity in renal cell carcinoma and in adjacent normal renal tissue. Urol. Res. 1998, 26, 117–121. [Google Scholar]
- Agundez, J.A. Cytochrome P450 gene polymorphism and cancer. Curr. Drug Metab. 2004, 5, 211–224. [Google Scholar]
- Kelman, Z. PCNA: Structure, functions and interactions. Oncogene 1997, 14, 629–640. [Google Scholar]
- Bravo, R.; Macdonald-Bravo, H. Existence of two populations of cyclin/proliferating cell nuclear antigen during the cell cycle: Association with DNA replication sites. J. Cell Biol. 1987, 105, 1549–1554. [Google Scholar]
- Woods, A.L.; Hall, P.A.; Shepherd, N.A.; Hanby, A.M.; Waseem, N.H.; Lane, D.P.; Levison, D.A. The assessment of proliferating cell nuclear antigen (PCNA) immunostaining in primary gastrointestinal lymphomas and its relationship to histological grade, S+G2+M phase fraction (flow cytometric analysis) and prognosis. Histopathology 1991, 19, 21–27. [Google Scholar]
- Gilles, C.; Polette, M.; Zahm, J.M.; Tournier, J.M.; Volders, L.; Foidart, J.M.; Birembaut, P. Vimentin contributes to human mammary epithelial cell migration. J. Cell Sci. 1999, 112, 4615–4625. [Google Scholar]
- Whipple, R.A.; Balzer, E.M.; Cho, E.H.; Matrone, M.A.; Yoon, J.R.; Martin, S.S. Vimentin filaments support extension of tubulin-based microtentacles in detached breast tumor cells. Cancer Res. 2008, 68, 5678–5688. [Google Scholar]
- Chen, H.; Herndon, M.E.; Lawler, J. The cell biology of thrombospondin-1. Matrix Biol. 2000, 19, 597–614. [Google Scholar]
- Wang, T.N.; Qian, X.; Granick, M.S.; Solomon, M.P.; Rothman, V.L.; Berger, D.H.; Tuszynski, G.P. Thrombospondin-1 (TSP-1) promotes the invasive properties of human breast cancer. J. Surg. Res. 1996, 63, 39–43. [Google Scholar]
- Yee, K.O.; Connolly, C.M.; Duquette, M.; Kazerounian, S.; Washington, R.; Lawler, J. The effect of thrombospondin-1 on breast cancer metastasis. Breast Cancer Res. Treat. 2009, 114, 85–96. [Google Scholar]
- Institute of Animal Laboratory Resources, Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 1996.
- Birkeland, S.A.; Storm, H.H. Glomerulonephritis and malignancy: A population-based analysis. Kidney Int. 2003, 63, 716–721. [Google Scholar]
- Rodgers, K.E. Effects of oral administration of malathion on the course of disease in MRL-lpr mice. J. Autoimmun. 1997, 10, 367–373. [Google Scholar]
- Oestreicher, E.M.; Guo, C.; Seely, E.W.; Kikuchi, T.; Martinez-Vasquez, D.; Jonasson, L.; Yao, T.; Burr, D.; Mayoral, S.; Roubsanthisuk, W.; Ricchiuti, V.; Adler, G.K. Estradiol increases proteinuria and angiotensin II type 1 receptor in kidneys of rats receiving L-NAME and angiotensin II. Kidney Int. 2006, 70, 1759–1768. [Google Scholar]
- Buratti, F.M.; D’Aniello, A.; Volpe, M.T.; Meneguz, A.; Testai, E. Malathion bioactivation in the human liver: The contribution of different cytochrome p450 isoforms. Drug Metab. Dispos. 2005, 33, 295–302. [Google Scholar]
- Prabhakaran, S.; Shameem, F.; Devi, K.S. Influence of protein deficiency on hexachlorocyclohexane and malathion toxicity in pregnant rats. Vet. Hum. Toxicol. 1993, 35, 429–433. [Google Scholar]
- Reus, G.Z.; Valvassori, S.S.; Nuernberg, H.; Comim, C.M.; Stringari, R.B.; Padilha, P.T.; Leffa, D.D.; Tavares, P.; Dagostim, G.; Paula, M.M.; Andrade, V.M.; Quevedo, J. DNA damage after acute and chronic treatment with malathion in rats. J. Agr. Food Chem. 2008, 56, 7560–7565. [Google Scholar]
- Possamai, F.P.; Fortunato, J.J.; Feier, G.; Agostinho, F.R.; Quevedo, J.; Wilhelm Filho, D.; Dal-Pizzol, F. Oxidative stress after acute and sub-chronic malathion intoxication in Wistar rats. Environ. Toxicol. Pharmacol. 2007, 23, 198–204. [Google Scholar]
- Uzun, F.G.; Kalender, S.; Durak, D.; Demir, F.; Kalender, Y. Malathion-induced testicular toxicity in male rats and the protective effect of vitamins C and E. Food Chem. Toxicol. 2009, 47, 1903–1908. [Google Scholar]
- Choudhary, N.; Goyal, R.; Joshi, S.C. Effect of malathion on reproductive system of male rats. J. Environ. Biol. 2008, 29, 259–262. [Google Scholar]
- Bosco, C.; Rodrigo, R.; Diaz, S.; Borax, J. Renal effects of chronic exposure to malathion in Octodon degus. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1997, 118, 247–253. [Google Scholar] [CrossRef]
- Bonner, M.R.; Coble, J.; Blair, A.; Beane Freeman, L.E.; Hoppin, J.A.; Sandler, D.P.; Alavanja, M.C. Malathion exposure and the incidence of cancer in the agricultural health study. Am. J. Epidemiol. 2007, 166, 1023–1034. [Google Scholar]
- Blair, A.; Dosemeci, M.; Heineman, E.F. Cancer and other causes of death among male and female farmers from twenty-three states. Am. J. Ind. Med. 1993, 23, 729–742. [Google Scholar]
- Zahm, S.H.; Ward, M.H.; Blair, A. Pesticides and cancer. Occup. Med. 1997, 12, 269–289. [Google Scholar]
- Bhat, H.K.; Calaf, G.; Hei, T.K.; Loya, T.; Vadgama, J.V. Critical role of oxidative stress in estrogen-induced carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 3913–3918. [Google Scholar]
- Liehr, J.G.; Chiappetta, C.; Roy, D.; Stancel, G.M. Elevation of protooncogene messenger RNAs in estrogen-induced kidney tumors in the hamster. Carcinogenesis 1992, 13, 601–604. [Google Scholar]
- Silbiger, S.; Lei, J.; Neugarten, J. Estradiol suppresses type I collagen synthesis in mesangial cells via activation of activator protein-1. Kidney Int. 1999, 55, 1268–1276. [Google Scholar]
- Terawaki, H.; Yoshimura, K.; Hasegawa, T.; Matsuyama, Y.; Negawa, T.; Yamada, K.; Matsushima, M.; Nakayama, M.; Hosoya, T.; Era, S. Oxidative stress is enhanced in correlation with renal dysfunction: Examination with the redox state of albumin. Kidney Int. 2004, 66, 1988–1993. [Google Scholar]
- Kobiela, J.; Krajewski, J.; Kalinska-Blach, B.; Stefaniak, T. Selectivity of oxidative stress targeting in estrogen-induced experimental nephrocarcinogenesis. Acta Biochim. Pol. 2002, 49, 51–58. [Google Scholar]
- Devanesan, P.; Todorovic, R.; Zhao, J.; Gross, M.L.; Rogan, E.G.; Cavalieri, E.L. Catechol estrogen conjugates and DNA adducts in the kidney of male Syrian golden hamsters treated with 4-hydroxyestradiol: Potential biomarkers for estrogen-initiated cancer. Carcinogenesis 2001, 22, 489–497. [Google Scholar]
- Liehr, J.G.; Roy, D. Free radical generation by redox cycling of estrogens. Free Rad. Biol. Med. 1990, 8, 415–423. [Google Scholar]
- Mense, S.M.; Remotti, F.; Bhan, A.; Singh, B.; El-Tamer, M.; Hei, T.K.; Bhat, H.K. Estrogen-induced breast cancer: Alterations in breast morphology and oxidative stress as a function of estrogen exposure. Toxicol. Appl. Pharmacol. 2008, 232, 78–85. [Google Scholar]
- Moore, P.D.; Yedjou, C.G.; Tchounwou, P.B. Malathion-induced oxidative stress, cytotoxicity, and genotoxicity in human liver carcinoma (HepG2) cells. Environ. Toxicol. 2010, 25, 221–226. [Google Scholar]
- Fortunato, J.J.; Feier, G.; Vitali, A.M.; Petronilho, F.C.; Dal-Pizzol, F.; Quevedo, J. Malathion-induced oxidative stress in rat brain regions. Neurochem. Res. 2006, 31, 671–678. [Google Scholar]
- Calaf, G.M.; Roy, D. Gene and protein expressions induced by 17beta-estradiol and parathion in cultured breast epithelial cells. Mol. Med. 2007, 13, 255–265. [Google Scholar]
- Calaf, G.M.; Roy, D. Cancer genes induced by malathion and parathion in the presence of estrogen in breast cells. Int. J. Mol. Med. 2008, 21, 261–268. [Google Scholar]
- Bretveld, R.W.; Thomas, C.M.; Scheepers, P.T.; Zielhuis, G.A.; Roeleveld, N. Pesticide exposure: The hormonal function of the female reproductive system disrupted? Reprod. Biol. Endocrinol. 2006, 4. [Google Scholar]
- Tanaka, Y.; Sasaki, M.; Kaneuchi, M.; Fujimoto, S.; Dahiya, R. Estrogen receptor alpha polymorphisms and renal cell carcinoma—A possible risk. Mol. Cell Endocrinol. 2003, 202, 109–116. [Google Scholar]
- Hopp, T.A.; Weiss, H.L.; Parra, I.S.; Cui, Y.; Osborne, C.K.; Fuqua, S.A. Low levels of estrogen receptor beta protein predict resistance to tamoxifen therapy in breast cancer. Clin. Cancer Res. 2004, 10, 7490–7499. [Google Scholar]
- Dubey, R.K.; Jackson, E.K. Estrogen-induced cardiorenal protection: Potential cellular, biochemical, and molecular mechanisms. Am. J. Physiol. Renal. Physiol. 2001, 280, F365–F388. [Google Scholar]
- Yager, J.D. Endogenous estrogens as carcinogens through metabolic activation. J. Natl. Cancer Inst. Monogr. 2000, 27, 67–73. [Google Scholar]
- Hong, Z.J.; Chu, C.H.; Fan, H.L.; Hsu, K.F.; Hsu, G.C.; Yu, C.P.; Char, D.L.; Yu, J.C. A progesterone-receptor-positive huge retroperitoneal tumour mimics metastasis in a breast cancer patient: Sarcomatoid renal cell carcinoma. Acta Chir. Belg. 2011, 111, 94–96. [Google Scholar]
- Meric, F.; Hung, M.C.; Hortobagyi, G.N.; Hunt, K.K. HER2/neu in the management of invasive breast cancer. J. Am. Coll. Surg. 2002, 194, 488–501. [Google Scholar]
- Gutierrez, C.; Schiff, R. HER2: Biology, detection, and clinical implications. Arch. Pathol. Lab Med. 2010, 135, 55–62. [Google Scholar]
- Imai, T.; Kimura, M.; Takeda, M.; Tomita, Y. Significance of epidermal growth factor receptor and c-erbB-2 protein expression in transitional cell cancer of the upper urinary tract for tumour recurrence at the urinary bladder. Br. J. Cancer 1995, 71, 69–72. [Google Scholar]
- Barouki, R.; Morel, Y. Repression of cytochrome P450 1A1 gene expression by oxidative stress: Mechanisms and biological implications. Biochem. Pharmacol. 2001, 61, 511–516. [Google Scholar]
- Meyer, R.P.; Podvinec, M.; Meyer, U.A. Cytochrome P450 CYP1A1 accumulates in the cytosol of kidney and brain and is activated by heme. Mol. Pharmacol. 2002, 62, 1061–1067. [Google Scholar]
- Hall, P.A.; Levison, D.A.; Woods, A.L.; Yu, C.C.; Kellock, D.B.; Watkins, J.A.; Barnes, D.M.; Gillett, C.E.; Camplejohn, R.; Dover, R.; et al. Proliferating cell nuclear antigen (PCNA) immunolocalization in paraffin sections: An index of cell proliferation with evidence of deregulated expression in some neoplasms. J. Pathol. 1990, 162, 285–294. [Google Scholar] [CrossRef]
- Kazerounian, S.; Yee, K.O.; Lawler, J. Thrombospondins in cancer. Cell Mol. Life Sci. 2008, 65, 700–712. [Google Scholar]
- Albo, D.; Arnoletti, J.P.; Castiglioni, A.; Granick, M.S.; Solomon, M.P.; Rothman, V.L.; Tuszynski, G.P. Thrombospondin (TSP) and transforming growth factor beta 1 (TGF-beta) promote human A549 lung carcinoma cell plasminogen activator inhibitor type 1 (PAI-1) production and stimulate tumor cell attachment in vitro. Biochem. Biophys. Res. Commun. 1994, 203, 857–865. [Google Scholar] [CrossRef]
- Cabello, G.; Valenzuela, M.; Vilaxa, A.; Durán, V.; Rudolph, I.; Hrepic, N.; Calaf, G.M. A rat mammary tumor model induced by the organophosphorous pesticides parathion and malathion, possibly through acetylcholinesterase inhibition. Environ. Health Perspect. 2001, 109, 471–479. [Google Scholar]
- Calaf, G.M.; Parra, E.; Garrido, F. Cell Proliferation and tumor formation induced by eserine, an acetylcholinesterase inhibitor, in rat mammary gland. Oncol. Rep. 2007, 17, 25–33. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alfaro-Lira, S.; Pizarro-Ortiz, M.; Calaf, G.M. Malignant Transformation of Rat Kidney Induced by Environmental Substances and Estrogen. Int. J. Environ. Res. Public Health 2012, 9, 1630-1648. https://doi.org/10.3390/ijerph9051630
Alfaro-Lira S, Pizarro-Ortiz M, Calaf GM. Malignant Transformation of Rat Kidney Induced by Environmental Substances and Estrogen. International Journal of Environmental Research and Public Health. 2012; 9(5):1630-1648. https://doi.org/10.3390/ijerph9051630
Chicago/Turabian StyleAlfaro-Lira, Susana, María Pizarro-Ortiz, and Gloria M. Calaf. 2012. "Malignant Transformation of Rat Kidney Induced by Environmental Substances and Estrogen" International Journal of Environmental Research and Public Health 9, no. 5: 1630-1648. https://doi.org/10.3390/ijerph9051630
APA StyleAlfaro-Lira, S., Pizarro-Ortiz, M., & Calaf, G. M. (2012). Malignant Transformation of Rat Kidney Induced by Environmental Substances and Estrogen. International Journal of Environmental Research and Public Health, 9(5), 1630-1648. https://doi.org/10.3390/ijerph9051630