Molecular Detection and Drug Resistance of Mycobacterium tuberculosis Complex from Cattle at a Dairy Farm in the Nkonkobe Region of South Africa: A Pilot Study

Abstract
:1. Introduction
2. Results and Discussion
2.1. Prevalence of MTBC amongst Different Cattle Breeds

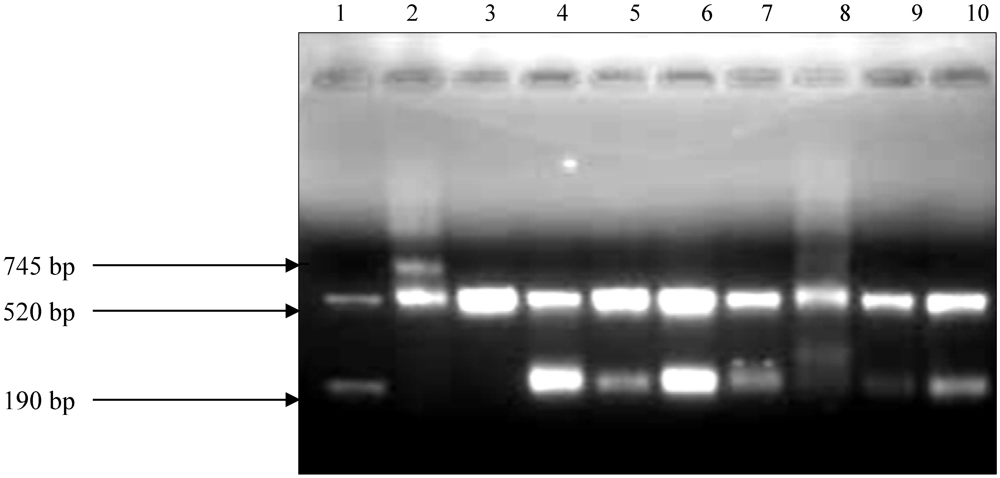{kind=link}
| Total herd | Breed | Age | Sex | Body condition | TB history | Vaccinated against BTB | Number positive for MTBC | Overall positivity per breed |
|---|---|---|---|---|---|---|---|---|
| 50 | Jersey | <3 years | F | Good | Unknown | No | 1 | 3 |
| >3 years | F | Good | Unknown | No | 2 | |||
| 50 | Friesland | <3 years | F | Good | Unknown | No | 1 | 4 |
| >3 years | F | Good | Unknown | No | 3 | |||
| 50 | Guernsey | <3 years | F | Good | Unknown | No | 1 | 2 |
| >3 years | F | Good | Unknown | No | 1 | |||
| 50 | Ayrshire | <3 years | F | Good | Unknown | No | 1 | 2 |
| >3 years | F | Good | Unknown | No | 1 | |||
| Total (200) | 11 | |||||||
2.2. Amplification of DNA Samples

2.3.Drug Susceptibility Results
| Antibiotic Susceptibility pattern | Genotype MTBDR plus assay | ||
|---|---|---|---|
| Number of samples | |||
| RIF | Resistant | 10 | |
| INH | Resistant | 10 | |
| RIF & INH | MDR | 10 | |
| RIF & INH | Sensitive | 1 | |
2.4. Mutations Conferring INH and RIF Resistance
| Mutation probe | Codon analyzed | Type of mutation | Number of samples |
|---|---|---|---|
| rpoB (RIF resistance ) | |||
| MUT1 | 513–519 | D516V | 2 |
| MUT2A | 526–529 | H526Y | 9 |
| MUT2B | 526–529 | H526D | 8 |
| MUT3 | 530–533 | S531L | 6 |
| katG (High level INH resistance) | |||
| MUT1 | 315 | S315T1 | 0 |
| MUT2 | 315 | S315T2 | 0 |
| inhA (Low level INH resistance) | |||
| MUT1 | –15 | C15T | 6 |
| MUT2 | –16 | A16G | 6 |
| MUT3A | –8 | T8C | 6 |
| MUT3B | –8 | T8A | 10 |
3. Experimental Section
3.1. Sampling
3.2. Decontamination of the Samples
3.3. DNA Extraction and Amplification
3.4. GenoType® MTBDRplus Assay
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Du, Y.; Qi, Y.; Yu, L.; Lin, J.; Liu, S.; Ni, H.; Pang, H.; Liu, H.; Si, W.; Zhao, H.; Wang, C. Molecular characterization of Mycobacterium tuberculosis complex (MTBC) isolated from cattle in northeast and northwest China. Res. Vet. Sci. 2011, 90, 385–391. [Google Scholar]
- Michel, A.L.; Hlokwe, T.M.; Coetzee, M.L.; Mare, L.; Connoway, L.; Rutten, V.P.M.G.; Kremer, K. High Mycobacterium bovis genetic diversity in a low prevalence setting. Vet. Microbiol. 2008, 126, 151–159. [Google Scholar] [CrossRef]
- Hlokwe, T.M.; Jenkins, A.O.; Streicher, E.M.; Venter, E.H.; Cooper, D.; Godfroid, J.; Michel, A.L. Molecular characterisation of Mycobacterium bovis isolated from African buffaloes (Syncerus caffer) in Hluhluwe-iMfolozi Park in KwaZulu-Natal, South Africa. Onderstepoort. J. Vet. Res. 2011, 78. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.L.; Bengis, R.G.; Keet, D.F.; Hofmeyr, M.; de Klerk, L.M.; Cross, P.C.; Jolles, A.E.; Cooper, D.; Whyte, I.J.; Buss, P.; Godfroid, J. Wildlife tuberculosis in South African conservation areas: Implications and challenges. Vet. Microbiol. 2006, 112, 91–100. [Google Scholar] [CrossRef]
- Lyashchenko, K.; Whelan, A.O.; Greenwald, R.; Pollock, J.M.; Andersen, P.; Hewinson, R.G.; Vordermeier, H.M. Association of tuberculin-boosted antibody responses with pathology and cell-mediated immunity in cattle vaccinated with Mycobacterium bovis BCG and infected with M. bovis. Infect. Immun. 2004, 72, 2462–2467. [Google Scholar]
- Angela, D.P.; Giuseppina, C.; Tony, F.V.; Bijo, B.; Fatmira, S.; Giuseppina, T. Detection of Mycobacterium tuberculosis complex in milk using polymerase chain reaction (PCR). Food Control 2006, 17, 776–780. [Google Scholar] [CrossRef]
- Chen, Y.; Chao, Y.; Deng, Q.; Liu, T.; Xiang, J.; Chen, J.; Zhou, J.; Zhan, Z.; Kuang, Y.; Cai, H.; Chen, H.; Guo, A. Potential challenges to the stop TB plan for humans in China; Cattle maintain M. bovis & M. tuberculosis. Tuberculosis 2009, 89, 95–100. [Google Scholar] [CrossRef]
- Jeon, B.Y.; Kim, S.C.; Je, S.; Kwak, J.; Cho, J.E.; Woo, J.T.; Seo, S.; Shim, H.S.; Park, B.O.; Lee, S.S.; Cho, S.N. Evaluation of enzyme-linked immunosorbent assay using milk samples as a potential screening test of bovine tuberculosis of dairy cows in Korea. Res. Vet. Sci. 2010, 88, 390–393. [Google Scholar] [CrossRef]
- Cosivi, O.; Grange, J.M.; Daborn, C.J.; Raviglione, M.C.; Fujikura, T.; Cousins, D.; Robinson, R.A.; Huchzermeyer, H.F.A.K.; de Kantor, I.; Meslin, F.X. Zoonotic tuberculosis due to Mycobacterium bovis in developing countries. Emerg. Infect. Dis. 1998, 4, 59–68. [Google Scholar] [CrossRef]
- Al-Saqur, I.M.; Al-Thwani, A.N.; Al-Attar, I.M. Detection of Mycobacteria spp in cow’s milk using conventional methods and PCR. J. Vet. Sci. 2009, 23, 259–262. [Google Scholar]
- Gibson, A.L.; Hewinson, G.; Goodchild, T.; Watt, B.; Story, A.; Inwald, J.; Drobniewski, F.A. Molecular epidemiology of disease due to Mycobacterium bovis in humans in the United Kingdom. J. Clin. Microbiol. 2004, 42, 431–434. [Google Scholar]
- Campbell, P.J.; Morlock, G.P.; Sikes, R.D.; Dalton, T.L.; Metchock, B.; Starks, A.M.; Hooks, D.P.; Cowan, L.S.; Plikaytis, B.B.; Posey, J.E. Molecular detection of mutations associated with first and second-line drug resistance compared with conventional drug susceptibility testing of Mycobacterium tuberculosis. Antimicrob. Agents. Chemother. 2011, 55, 2032–2041. [Google Scholar]
- Green, E.; Obi, L.; Nchabeleng, M.; de Villiers, B.E.; Sein, P.P.; Letsoalo, T.; Hoosen, A.A.; Bessong, P.O.; Ndip, R.N. Drug-susceptibility patterns of Mycobacterium tuberculosis in Mpumalanga Province, South Africa: Possible guiding design of retreatment regimen. J. Health. Popul. Nutr. 2010, 28, 7–13. [Google Scholar]
- Da Silva, P.E.A.; Palomino, J.C. Molecular basis and mechanisms of drug resistance in Mycobacterium tuberculosis: Classical and new drugs. J. Antimicrob. Chemother. 2011, 66, 1417–1430. [Google Scholar] [CrossRef]
- Wilson, M.L. Recent advances in the laboratory detection of Mycobacterium tuberculosis complex and drug resistance. Clin. Infect. Dis. 2011, 52, 1350–1353. [Google Scholar] [CrossRef]
- Aslan, G.; Tezcan, S.; Serin, M.S.; Emekdas, G. Genotypic analysis of isoniazid and rifampin resistance in drug resistant clinical Mycobacterium tuberculosis complex isolates in southern Turkey. Jpn. J. Infect. Dis. 2008, 61, 255–260. [Google Scholar]
- Mokrousov, I.; Filliol, I.; Legrand, E.; Sola, C.; Otten, T.; Vyshnevskaya, E.; Limeschenko, E.; Vyshnevskiy, B.; Narvskaya, O.; Rastogi, N. Molecular characterization of multiple-drug-resistant Mycobacterium tuberculosis isolates from northwestern Russia and analysis of rifampin resistance using RNA/RNA mismatch analysis as compared to the line probe assay and sequencing of the rpoB gene. Res. Microbiol. 2002, 153, 213–219. [Google Scholar] [CrossRef]
- Lacoma, A.; Sierra, N.G.; Prat, C.; Manzano, J.R.; Haba, L.; Roses, S.; Maldonado, J.; Dominguez, J. GenoType MTBDRplus assay for molecular detection of rifampin and isoniazid resistance in Mycobacterium tuberculosis strains and clinical samples. J. Clin. Microbiol. 2008, 46, 3660–3667. [Google Scholar]
- Vitale, F.; Capra, G.; Maxia, L.; Reale, S.; Vesco, G.; Caracappa, S. Detection of Mycobacterium tuberculosis complex in cattle by PCR using milk, lymph node aspirates, and nasal swabs. J. Clin. Microbiol. 1998, 36, 1050–1055. [Google Scholar]
- Humblet, M.F.; Boschiroli, M.L.; Saegerman, C. Classification of worldwide bovine tuberculosis risk factors in cattle: A stratified approach. Vet. Res. 2009, 40. [Google Scholar] [CrossRef]
- Kazwala, R.R.; Kambarage, D.M.; Daborn, C.J.; Nyange, J.; Jiwa, S.F.H.; Sharp, J.M. Risk factors associated with bovine tuberculosis in cattle in the southern highlands of Tanzania. Vet. Res. Commun. 2001, 25, 609–614. [Google Scholar] [CrossRef]
- Munyeme, M.; Muma, J.B.; Skjerve, E.; Nambota, A.M.; Phiri, I.G.K.; Samui, K.L.; Dorny, P.; Tryland, M. Risk factors associated with bovine tuberculosis in traditional cattle of the livestock/wildlife interface areas in the Kafue basin of Zambia. Prev.Vet. Med. 2008, 85, 317–328. [Google Scholar] [CrossRef]
- Ameni, G.; Erkihun, A. Bovine tuberculosis on small-scale dairy farms in Adama Town, central Ethiopia, and farmer awareness of the disease. Rev. Sci. Tech. Off. Int. Epiz. 2007, 26, 711–719. [Google Scholar]
- Ameni, G.; Vordermeier, M.; Firdessa, R.; Aseffa, A.; Hewinson, G.; Gordon, S.V.; Berg, S. Mycobacterium tuberculosis infection in grazing cattle in central Ethiopia. Vet. J. 2011, 188, 359–361. [Google Scholar] [CrossRef]
- Srivastava, K.; Chauhan, D.S.; Gupta, P.; Singh, H.B.; Sharma, V.D.; Yadav, V.S.; Sreekumaran, P.; Thakral, S.S.; Dharamdheeran, J.S.; Nigam, P.; Prasad, H.K.; Katoch, V.M. Isolation of Mycobacterium bovis & M. tuberculosis from cattle of some farms in north India—Possible relevance in human health. Indian J. Med. Res. 2008, 128, 26–31. [Google Scholar]
- Sechi, L.A.; Zanetti, S.; Sanguinetti, M.; Molicotti, P.; Romano, L.; Leori, G.; Delogu, G.; Boccia, S.; La Sorda, M.; Fadda, G. Molecular basis of rifampin and isoniazid resistance in Mycobacterium bovis strains isolated in Sardinia, Italy. Antimicrob. Agents. Chemother. 2001, 45, 1645–1647. [Google Scholar]
- Hofmann-Thiel, S.; van Ingen, J.; Feldmann, K.; Turaev, L.; Uzakova, G.T.; Murmusaeva, G.; van Soolingen, D.; Hoffmann, H. Mechanisms of heteroresistance to isoniazid and rifampin of Mycobacterium tuberculosis in Tashkent, Uzbekistan. Eur. Respir. J. 2009, 33, 368–374. [Google Scholar]
- Bazira, J.; Asiimwe, B.B.; Joloba, M.L.; Bwanga, F.; Matee, M.I. Use of the GenoType® MTBDRplus assay to assess drug resistance of Mycobacterium tuberculosis isolates from patients in rural Uganda. BMC Clin. Pathol. 2010, 10. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silaigwana, B.; Green, E.; Ndip, R.N. Molecular Detection and Drug Resistance of Mycobacterium tuberculosis Complex from Cattle at a Dairy Farm in the Nkonkobe Region of South Africa: A Pilot Study. Int. J. Environ. Res. Public Health 2012, 9, 2045-2056. https://doi.org/10.3390/ijerph9062045
Silaigwana B, Green E, Ndip RN. Molecular Detection and Drug Resistance of Mycobacterium tuberculosis Complex from Cattle at a Dairy Farm in the Nkonkobe Region of South Africa: A Pilot Study. International Journal of Environmental Research and Public Health. 2012; 9(6):2045-2056. https://doi.org/10.3390/ijerph9062045
Chicago/Turabian StyleSilaigwana, Blessing, Ezekiel Green, and Roland N. Ndip. 2012. "Molecular Detection and Drug Resistance of Mycobacterium tuberculosis Complex from Cattle at a Dairy Farm in the Nkonkobe Region of South Africa: A Pilot Study" International Journal of Environmental Research and Public Health 9, no. 6: 2045-2056. https://doi.org/10.3390/ijerph9062045
APA StyleSilaigwana, B., Green, E., & Ndip, R. N. (2012). Molecular Detection and Drug Resistance of Mycobacterium tuberculosis Complex from Cattle at a Dairy Farm in the Nkonkobe Region of South Africa: A Pilot Study. International Journal of Environmental Research and Public Health, 9(6), 2045-2056. https://doi.org/10.3390/ijerph9062045