

Methamphetamine and Designer Stimulants Modulate Tonic Human Cerebrovascular Smooth Muscle Contractility: Relevance to Drug-Induced Neurovascular Stress

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Rat-Tail Type-1 Collagen Preparation
2.3. Preparation of HBVSMC/Collagen Gels
2.4. Drugs and Chemicals
2.5. Exposure Regimen
2.6. Image Collection and Analysis
2.7. MTT Assay
2.8. Data Analysis
3. Results
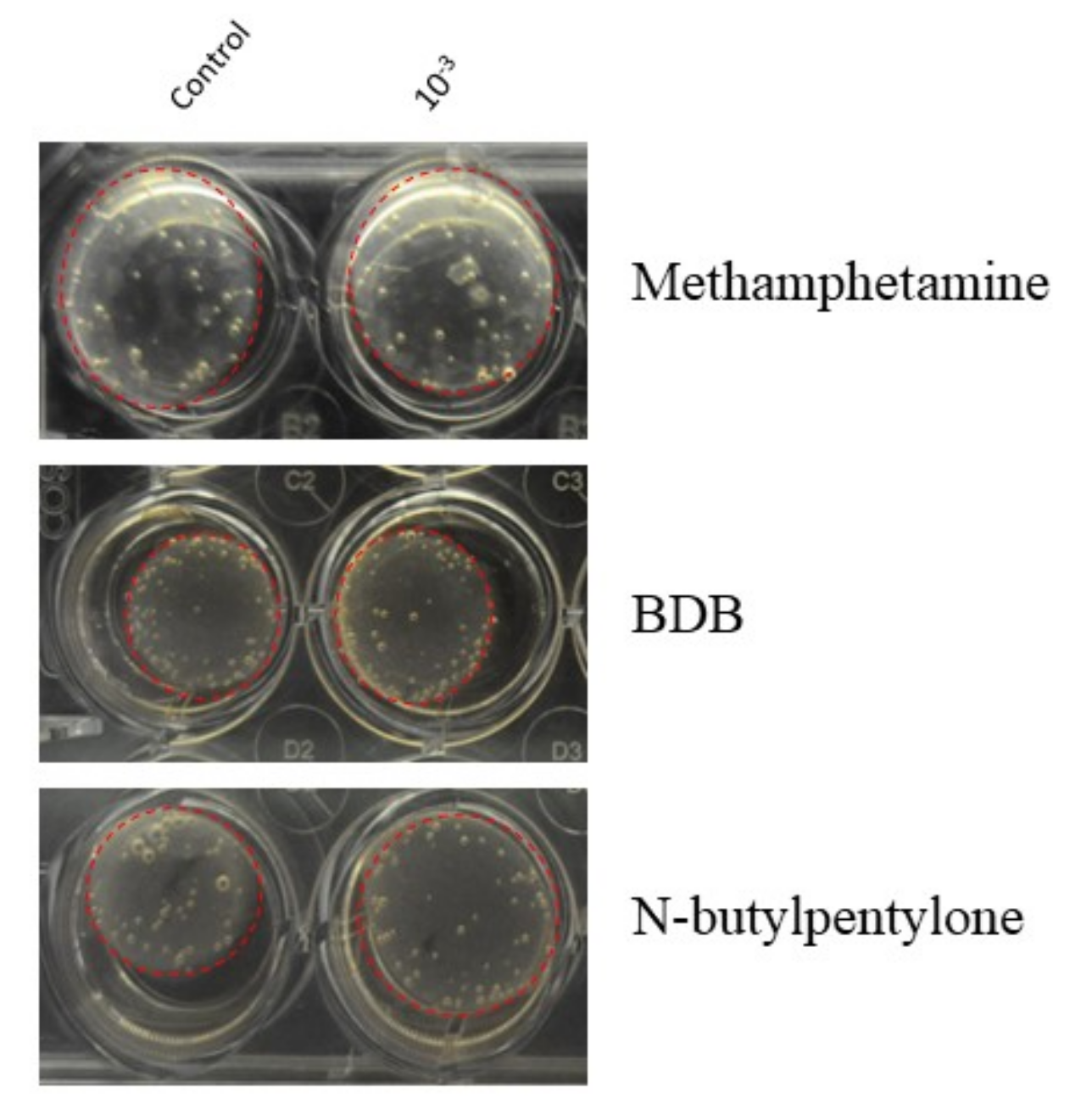
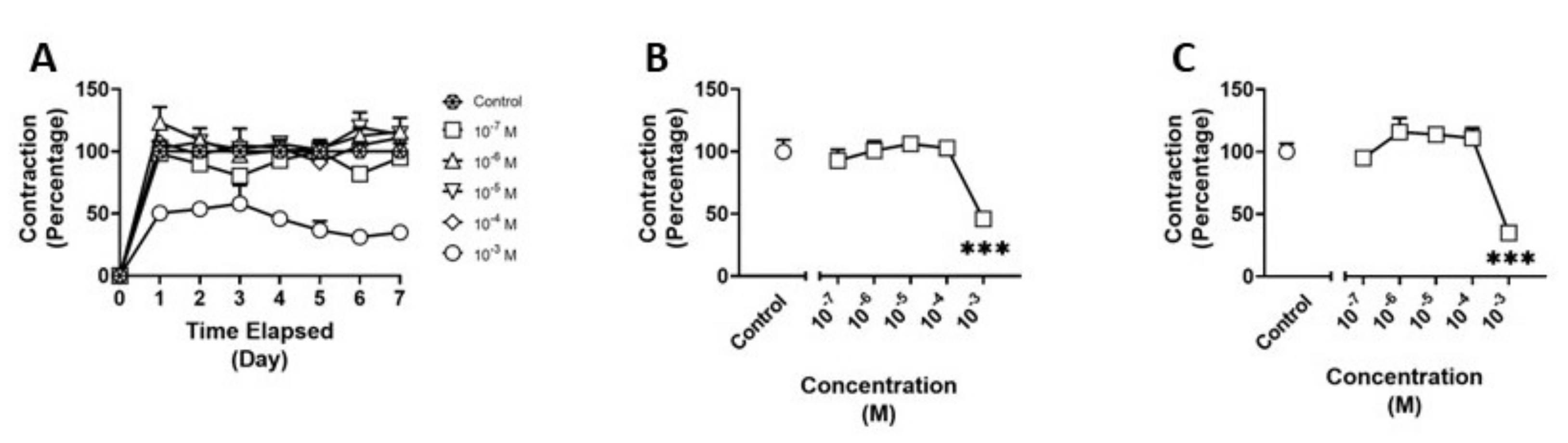
3.1. Effects of Methamphetamine on HBVSMC
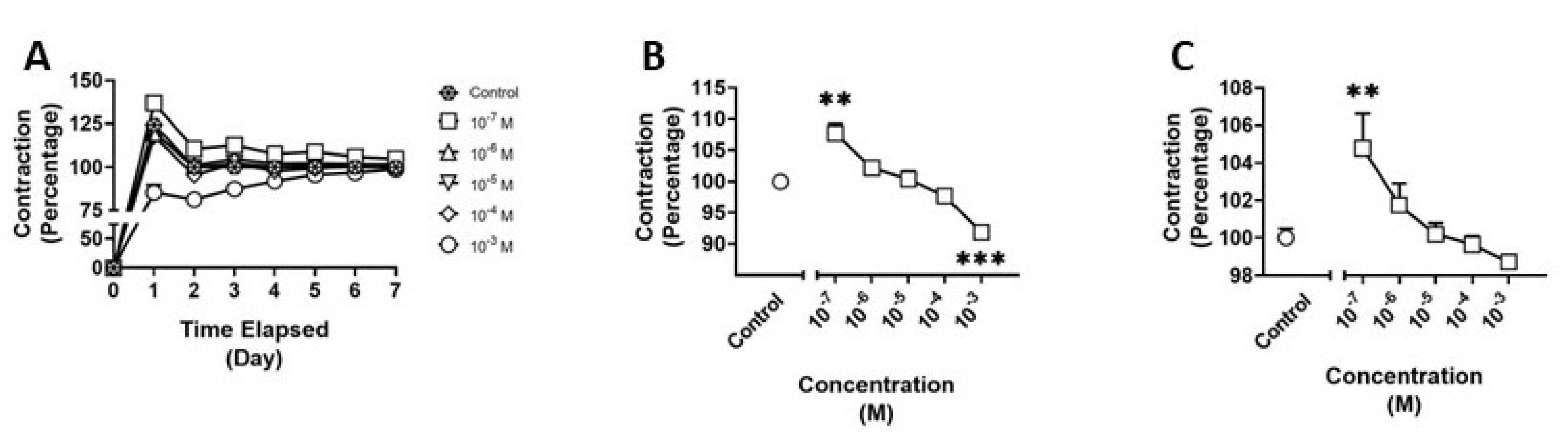
3.2. Effects of BDB on HBVSMC
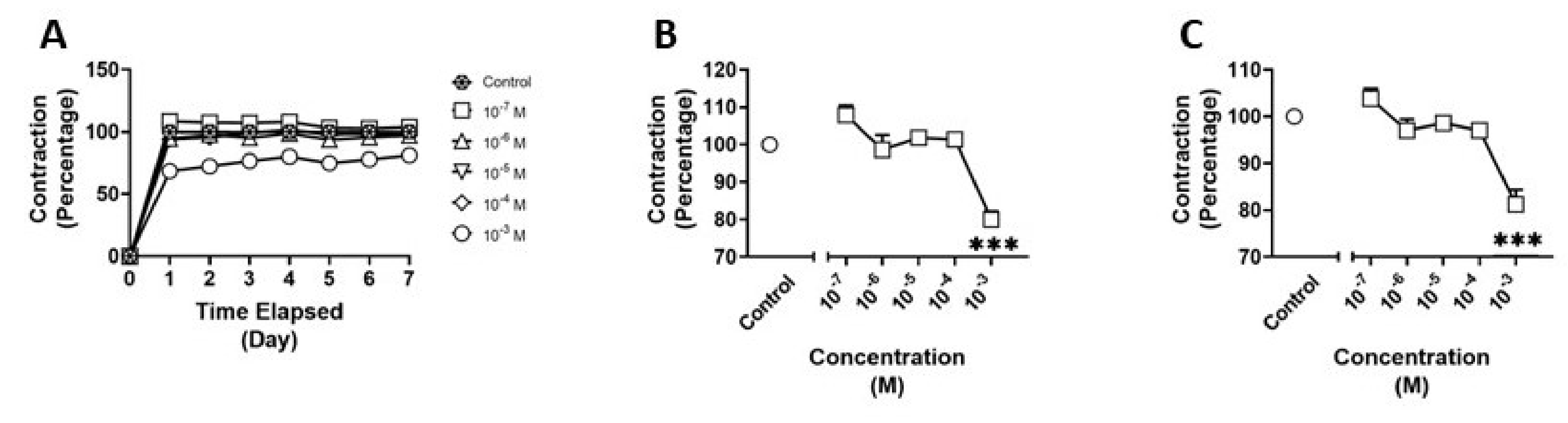
3.3. Effects of N-butylpentylone on HBVSMC
3.4. MTT Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, T.; Spiller, H.A.; Badeti, J.; Funk, A.R.; Zhu, M.; Smith, G.A. Methamphetamine exposures reported to United States poison control centers, 2000–2019. Clin. Toxicol. 2021, 59, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, H.; Bastian, B.A.; Trinidad, J.P.; Spencer, M.R.; Warner, M. Regional Differences in the Drugs Most Frequently Involved in Drug Overdose Deaths: United States, 2017. Natl. Vital-Stat. Rep. 2019, 68, 1–16. [Google Scholar] [PubMed]
- Chitre, N.M.; Bagwell, M.S.; Murnane, K.S. The acute toxic and neurotoxic effects of 3,4-methylenedioxymethamphetamine are more pronounced in adolescent than adult mice. Behav. Brain Res. 2020, 380, 112413. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Canal, C.E.; Ehlen, J.C.; Rice, K.C.; Murnane, K.S. Murnane, M100907 and BD 1047 attenuate the acute toxic effects of methamphetamine. Neurotoxicology 2019, 74, 91–99. [Google Scholar] [CrossRef]
- Andersen, M.L.; Diaz, M.P.; Murnane, K.S.; Howell, L.L. Effects of methamphetamine self-administration on actigraphy-based sleep parameters in rhesus monkeys. Psychopharmacology 2013, 227, 101–107. [Google Scholar] [CrossRef]
- Batra, V.; Murnane, K.S.; Knox, B.; Edinoff, A.N.; Ghaffar, Y.; Nussdorf, L.; Petersen, M.; Kaufman, S.E.; Jiwani, S.; Casey, C.A.; et al. Early onset cardiovascular disease related to methamphetamine use is most striking in individuals under 30: A retrospective chart review. Addict. Behav. Rep. 2022, 15, 100435. [Google Scholar] [CrossRef]
- Dominic, P.; Ahmad, J.; Awwab, H.; Bhuiyan, S.; Kevil, C.G.; Goeders, N.E.; Murnane, K.S.; Patterson, J.C.; Sandau, K.E.; Gopinathannair, R.; et al. Stimulant Drugs of Abuse and Cardiac Arrhythmias. Circ. Arrhythmia Electrophysiol. 2022, 15, e010273. [Google Scholar] [CrossRef]
- Scott, M.L.; Murnane, K.S.; Orr, A.W. Young at heart? Drugs of abuse cause early-onset cardiovascular disease in the young. Heart 2021, 107, 604–606. [Google Scholar] [CrossRef]
- Kevil, C.G.; Goeders, N.E.; Woolard, M.D.; Bhuiyan, S.; Dominic, P.; Kolluru, G.K.; Arnold, C.L.; Traylor, J.G.; Orr, A.W. Methamphetamine Use and Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1739–1746. [Google Scholar] [CrossRef]
- Murnane, K.S.; Howell, L.L. Neuroimaging and drug taking in primates. Psychopharmacology 2011, 216, 153–171. [Google Scholar] [CrossRef]
- Howell, L.L.; Murnane, K.S. Nonhuman primate neuroimaging and the neurobiology of psychostimulant addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 176–194. [Google Scholar] [CrossRef]
- Howell, L.L.; Murnane, K.S. Nonhuman primate positron emission tomography neuroimaging in drug abuse research. J. Pharmacol. Exp. Ther. 2011, 337, 324–334. [Google Scholar] [CrossRef]
- Fowler, J.S.; Volkow, N.D.; Wolf, A.P.; Dewey, S.L.; Schlyer, D.J.; MacGregor, R.R.; Hitzemann, R.; Logan, J.; Bendriem, B.; Gatley, S.J.; et al. Mapping cocaine binding sites in human and baboon brain in vivo. Synapse 1989, 4, 371–377. [Google Scholar] [CrossRef]
- Volkow, N.D.; Ding, Y.S.; Fowler, J.S.; Wang, G.J.; Logan, J.; Gatley, J.S.; Dewey, S.; Ashby, C.; Liebermann, J.; Hitzemann, R.; et al. Is methylphenidate like cocaine? Studies on their pharmacokinetics and distribution in the human brain. Arch. Gen. Psychiatry 1995, 52, 456–463. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Fischman, M.W.; Foltin, R.W.; Fowler, J.S.; Abumrad, N.N.; Vitkun, S.; Logan, J.; Gatley, S.J.; Pappas, N.; et al. Relationship between subjective effects of cocaine and dopamine transporter occupancy. Nature 1997, 386, 827–830. [Google Scholar] [CrossRef]
- Iversen, L.; Gibbons, S.; Treble, R.; Setola, V.; Huang, X.-P.; Roth, B.L. Neurochemical profiles of some novel psychoactive substances. Eur. J. Pharmacol. 2013, 700, 147–151. [Google Scholar] [CrossRef]
- Marusich, J.A.; Antonazzo, K.R.; Wiley, J.L.; Blough, B.E.; Partilla, J.S.; Baumann, M.H. Pharmacology of novel synthetic stimulants structurally related to the "bath salts" constituent 3,4-methylenedioxypyrovalerone (MDPV). Neuropharmacology 2014, 87, 206–213. [Google Scholar] [CrossRef]
- Reith, M.E.; Blough, B.E.; Hong, W.C.; Jones, K.T.; Schmitt, K.C.; Baumann, M.H.; Partilla, J.S.; Rothman, R.B.; Katz, J.L. Behavioral, biological, and chemical perspectives on atypical agents targeting the dopamine transporter. Drug Alcohol Depend. 2015, 147, 1–19. [Google Scholar] [CrossRef]
- Baumann, M.H.; Solis, E.; Watterson, L.R.; Marusich, J.A.; Fantegrossi, W.E.; Wiley, J.L. Baths salts, spice, and related designer drugs: The science behind the headlines. J. Neurosci. 2014, 34, 15150–15158. [Google Scholar] [CrossRef]
- Angoa-Pérez, M.; Anneken, J.H.; Kuhn, D.M. Neurotoxicology of Synthetic Cathinone Analogs. Curr. Top Behav. Neurosci. 2017, 32, 209–230. [Google Scholar]
- Simmler, L.; Buser, T.; Donzelli, M.; Schramm, Y.; Dieu, L.; Huwyler, J.; Chaboz, S.; Hoener, M.; Liechti, M. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Simmler, L.; Rickli, A.; Hoener, M.; Liechti, M. Monoamine transporter and receptor interaction profiles of a new series of designer cathinones. Neuropharmacology 2014, 79, 152–160. [Google Scholar] [CrossRef] [PubMed]
- López-Arnau, R.; Martínez-Clemente, J.; Pubill, D.; Escubedo, E.; Camarasa, J. Comparative neuropharmacology of three psychostimulant cathinone derivatives: Butylone, mephedrone and methylone. Br. J. Pharmacol. 2012, 167, 407–420. [Google Scholar] [CrossRef]
- Eshleman, A.J.; Wolfrum, K.M.; Hatfield, M.G.; Johnson, R.A.; Murphy, K.V.; Janowsky, A. Substituted methcathinones differ in transporter and receptor interactions. Biochem. Pharmacol. 2013, 85, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Svensson, T.H. Alpha-adrenoceptor modulation hypothesis of antipsychotic atypicality. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 1145–1158. [Google Scholar] [CrossRef]
- Carter, O.L.; Pettigrew, J.D.; Hasler, F.; Wallis, G.; Liu, G.B.; Hell, D.; Vollenweider, F.X. Modulating the rate and rhythmicity of perceptual rivalry alternations with the mixed 5-HT2A and 5-HT1A agonist psilocybin. Neuropsychopharmacology 2005, 30, 1154–1162. [Google Scholar] [CrossRef]
- Stratman, A.N.; Pezoa, S.A.; Farrelly, O.M.; Castranova, D.; Dye, L.E., 3rd; Butler, M.G.; Sidik, H.; Talbot, W.S.; Weinstein, B.M. Interactions between mural cells and endothelial cells stabilize the developing zebrafish dorsal aorta. Development 2017, 144, 115–127. [Google Scholar]
- Tan, X.; Cai, D.; Chen, N.; Du, S.; Qiao, D.; Yue, X.; Wang, T.; Li, J.; Xie, W.; Wang, H. Methamphetamine mediates apoptosis of vascular smooth muscle cells via the chop-related endoplasmic reticulum stress pathway. Toxicol. Lett. 2021, 350, 98–110. [Google Scholar] [CrossRef]
- Liang, L.-Y.; Wang, M.-M.; Liu, M.; Zhao, W.; Wang, X.; Shi, L.; Zhu, M.-J.; Zhao, Y.-L.; Liu, L.; Maurya, P.; et al. Chronic toxicity of methamphetamine: Oxidative remodeling of pulmonary arteries. Toxicol. Vitr. 2020, 62, 104668. [Google Scholar] [CrossRef]
- Nabaei, G.; Oveisgharan, S.; Ghorbani, A.; Fatehi, F. Impaired arterial smooth muscle cell vasodilatory function in methamphetamine users. J. Neurol. Sci. 2016, 370, 107–111. [Google Scholar] [CrossRef]
- Zolkowska, D.; Rothman, R.B.; Baumann, M.H. Amphetamine analogs increase plasma serotonin: Implications for cardiac and pulmonary disease. J. Pharmacol. Exp. Ther. 2006, 318, 604–610. [Google Scholar] [CrossRef]
- Ford, C.L.; Wang, Y.; Morgan, K.; Boktor, M.; Jordan, P.; Castor, T.P.; Alexander, J.S. Interferon-gamma depresses human intestinal smooth muscle cell contractility: Relevance to inflammatory gut motility disturbances. Life Sci. 2019, 222, 69–77. [Google Scholar] [CrossRef]
- Al-Kofahi, M.; Becker, F.; Gavins, F.N.; Woolard, M.D.; Tsunoda, I.; Wang, Y.; Ostanin, D.; Zawieja, D.C.; Muthuchamy, M.; von der Weid, P.Y.; et al. IL-1beta reduces tonic contraction of mesenteric lymphatic muscle cells, with the involvement of cycloxygenase-2 and prostaglandin E2. Br. J. Pharmacol. 2015, 172, 4038–4051. [Google Scholar] [CrossRef]
- Seo, J.-W.; Jones, S.M.; Hostetter, T.A.; Iliff, J.J.; West, G.A. Methamphetamine induces the release of endothelin. J. Neurosci. Res. 2016, 94, 170–178. [Google Scholar] [CrossRef]
- Polesskaya, O.; Silva, J.; Sanfilippo, C.; Desrosiers, T.; Sun, A.; Shen, J.; Feng, C.; Polesskiy, A.; Deane, R.; Zlokovic, B.; et al. Methamphetamine causes sustained depression in cerebral blood flow. Brain Res. 2011, 1373, 91–100. [Google Scholar] [CrossRef]
- Kousik, S.M.; Graves, S.M.; Napier, T.C.; Zhao, C.; Carvey, P.M. Methamphetamine-induced vascular changes lead to striatal hypoxia and dopamine reduction. Neuroreport 2011, 22, 923–928. [Google Scholar] [CrossRef]
- Nagai, F.; Nonaka, R.; Kamimura, K.S.H. The effects of non-medically used psychoactive drugs on monoamine neurotransmission in rat brain. Eur. J. Pharmacol. 2007, 559, 132–137. [Google Scholar] [CrossRef]
- Meyer, M.R.; Peters, F.T.; Maurer, H.H. Investigations on the human hepatic cytochrome P450 isozymes involved in the metabolism of 3,4-methylenedioxy-amphetamine (MDA) and benzodioxolyl-butanamine (BDB) enantiomers. Toxicol. Lett. 2009, 190, 54–60. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, N.; Dao, N.; Hewett, C.; Oberle, S.; Minagar, A.; Lamon, K.; Ford, C.; Blough, B.E.; Alexander, J.S.; Murnane, K.S. Methamphetamine and Designer Stimulants Modulate Tonic Human Cerebrovascular Smooth Muscle Contractility: Relevance to Drug-Induced Neurovascular Stress. Pathophysiology 2023, 30, 144-154. https://doi.org/10.3390/pathophysiology30020013
Hall N, Dao N, Hewett C, Oberle S, Minagar A, Lamon K, Ford C, Blough BE, Alexander JS, Murnane KS. Methamphetamine and Designer Stimulants Modulate Tonic Human Cerebrovascular Smooth Muscle Contractility: Relevance to Drug-Induced Neurovascular Stress. Pathophysiology. 2023; 30(2):144-154. https://doi.org/10.3390/pathophysiology30020013
Chicago/Turabian StyleHall, Nicole, Nhi Dao, Cameron Hewett, Sara Oberle, Andrew Minagar, Kariann Lamon, Carey Ford, Bruce E. Blough, J. Steven Alexander, and Kevin S. Murnane. 2023. "Methamphetamine and Designer Stimulants Modulate Tonic Human Cerebrovascular Smooth Muscle Contractility: Relevance to Drug-Induced Neurovascular Stress" Pathophysiology 30, no. 2: 144-154. https://doi.org/10.3390/pathophysiology30020013
APA StyleHall, N., Dao, N., Hewett, C., Oberle, S., Minagar, A., Lamon, K., Ford, C., Blough, B. E., Alexander, J. S., & Murnane, K. S. (2023). Methamphetamine and Designer Stimulants Modulate Tonic Human Cerebrovascular Smooth Muscle Contractility: Relevance to Drug-Induced Neurovascular Stress. Pathophysiology, 30(2), 144-154. https://doi.org/10.3390/pathophysiology30020013