
Myocardial Glutathione Synthase and TRXIP Expression Are Significantly Elevated in Hypertension and Diabetes: Influence of Stress on Antioxidant Pathways


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design
2.3. Modeling of Insulin-Dependent DM
2.4. Morphological and Immunohistochemical Study
2.5. Statistics
3. Results
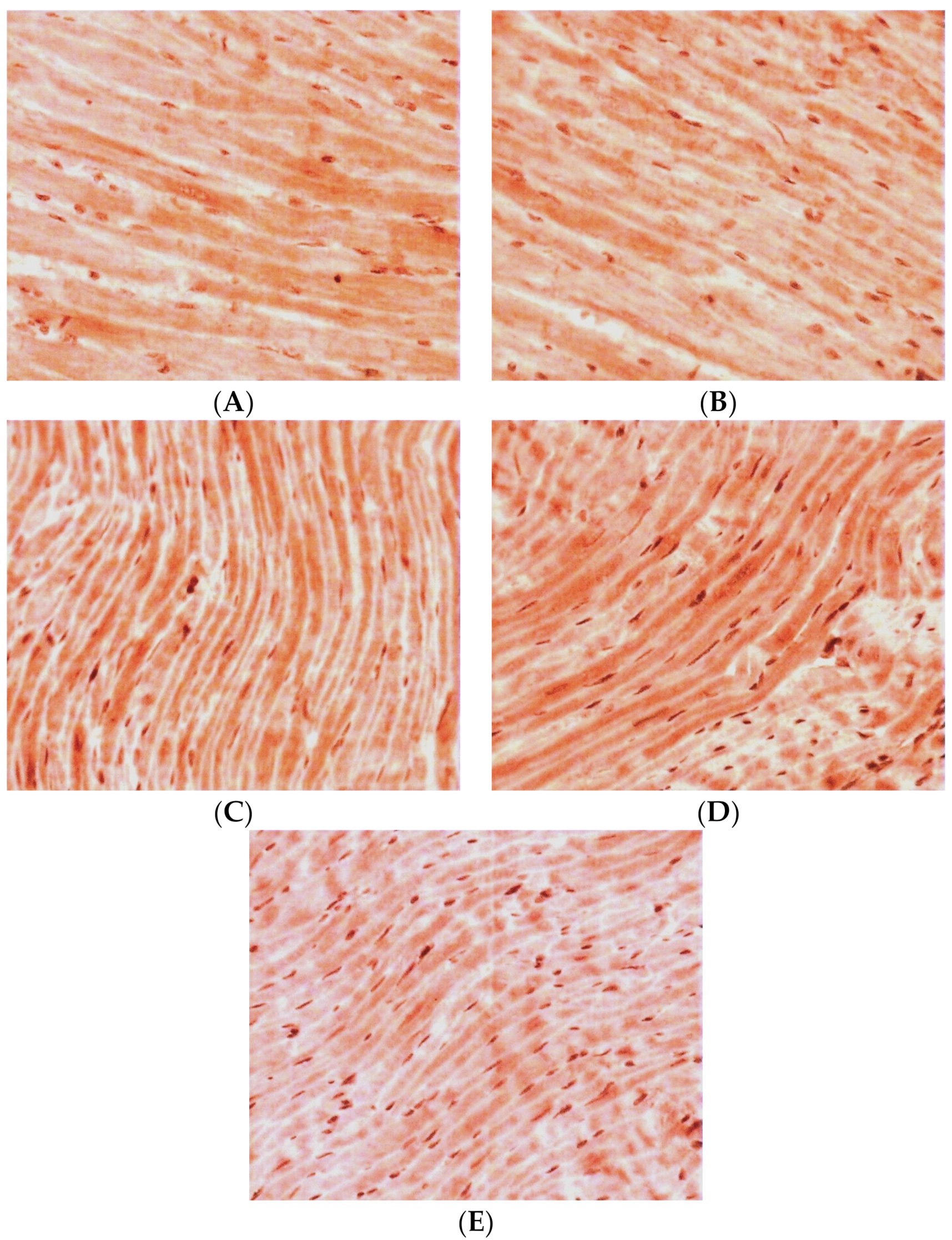
3.1. Immunohistochemical Study of TXNIP Expression in the LV Myocardium
3.2. Immunohistochemical Study of GS Expression in the LV Myocardium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borowiec, A.M.; Właszczuk, A.; Olakowska, E.; Lewin-Kowalik, J. TXNIP inhibition in the treatment of diabetes. Verapamil as a novel therapeutic modality in diabetic patients. Med. Pharm. Rep. 2022, 95, 243–250. [Google Scholar] [CrossRef]
- Yoshihara, E. TXNIP/TBP-2: A master regulator for glucose homeostasis. Antioxidants 2020, 9, 765. [Google Scholar] [CrossRef]
- Alhawiti, N.M.; Al Mahri, S.; Aziz, M.A.; Malik, S.S.; Mohammad, S. TXNIP in metabolic regulation: Physiological role and therapeutic outlook. Curr. Drug Targets 2017, 18, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.J.; Zhang, Y.; Shi, W.J.; Lei, Z.D.; Jiao, X.Y. TXNIP knockout improves cardiac function after myocardial infarction by promoting angiogenesis and reducing cardiomyocyte apoptosis. Cardiovasc. Diagn. Ther. 2022, 12, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, H.; Tooyama, I.; Walker, D.G. Thioredoxin-interacting protein (TXNIP) with focus on brain and neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 9357. [Google Scholar] [CrossRef]
- Jeffrey, S.; Samraj, P.I.; Raj, B.S. The role of alpha-lipoic acid supplementation in the prevention of diabetes complications: A comprehensive review of clinical trials. Curr. Diabetes Rev. 2021, 17, e011821190404. [Google Scholar] [CrossRef]
- Avagimyan, A.; Popov, S.; Shalnova, S. The pathophysiological basis of diabetic cardiomyopathy development. Curr. Probl. Cardiol. 2022, 47, 101156. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tonissen, K.F.; Di Trapani, G. Modulating skin colour: Role of the thioredoxin and glutathione systems in regulating melanogenesis. Biosci. Rep. 2021, 41, BSR20210427. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Vivancos, P.; De Simone, A.; Kiddle, G.; Foyer, C.H. Glutathione–linking cell proliferation to oxidative stress. Free Radic. Biol. Med. 2015, 89, 1154–1164. [Google Scholar] [CrossRef]
- Wu, X.; Li, Y.; Zhang, S.; Zhou, X. Ferroptosis as a novel therapeutic target for cardiovascular disease. Theranostics 2021, 11, 3052–3059. [Google Scholar] [CrossRef]
- Okada, M.; Kimura, Y. Characterization of glutamate-cysteine ligase and glutathione synthetase from the δ-proteobacterium Myxococcus xanthus. Proteins 2022, 90, 1547–1560. [Google Scholar] [CrossRef]
- Zhang, H.; Forman, H.J. Glutathione synthesis and its role in redox signaling. Semin. Cell Dev. Biol. 2012, 23, 722–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, R.; Basnet, T.B.; Basnet, B.B.; Khadka, S. Overview on thioredoxin-interacting protein (TXNIP): A potential target for diabetes intervention. Curr. Drug Targets 2022, 23, 761–767. [Google Scholar] [CrossRef]
- Masson, E.; Koren, S.; Razik, F.; Goldberg, H.; Kwan, E.P.; Sheu, L.; Gaisano, H.Y.; Fantus, I.G. High beta-cell mass prevents streptozotocin-induced diabetes in thioredoxin-interacting protein-deficient mice. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1251–E1261. [Google Scholar] [CrossRef] [Green Version]
- Parikh, H.; Carlsson, E.; Chutkow, W.A.; Johansson, L.E.; Storgaard, H.; Poulsen, P.; Saxena, R.; Ladd, C.; Schulze, P.C.; Mazzini, M.J.; et al. TXNIP regulates peripheral glucose metabolism in humans. PLoS Med. 2007, 4, e158. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.W.; Lan, T.; Zheng, F.; Huang, H.; Chen, H.F.; Wu, Q.; Zhang, F. Celastrol inhibits TXNIP expression to protect pancreatic β cells in diabetic mice. Phytomedicine 2022, 104, 154316. [Google Scholar] [CrossRef] [PubMed]
- Wondafrash, D.Z.; Nire’a, A.T.; Tafere, G.G.; Desta, D.M.; Berhe, D.A.; Zewdie, K.A. Thioredoxin-interacting protein as a novel potential therapeutic target in diabetes mellitus and its underlying complications. Diabetes Metab. Syndr. Obes. 2020, 13, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, N.; Nakayama, Y.; Abdali, S.A.; Yoshioka, J. Cardiomyocyte-specific Txnip C247S mutation improves left ventricular functional reserve in streptozotocin-induced diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H259–H274. [Google Scholar] [CrossRef]
- Yosri, H.; El-Kashef, D.H.; El-Sherbiny, M.; Said, E.; Salem, H.A. Calycosin modulates NLRP3 and TXNIP-mediated pyroptotic signaling and attenuates diabetic nephropathy progression in diabetic rats; An insight. Biomed. Pharmacother. 2022, 155, 113758. [Google Scholar] [CrossRef]
- Song, S.; Qiu, D.; Shi, Y.; Wang, S.; Zhou, X.; Chen, N.; Wei, J.; Wu, M.; Wu, H.; Duan, H. Thioredoxin-interacting protein deficiency alleviates phenotypic alterations of podocytes via inhibition of mTOR activation in diabetic nephropathy. J. Cell Physiol. 2019, 234, 16485–16502. [Google Scholar] [CrossRef]
- Gao, C.; Wang, R.; Li, B.; Guo, Y.; Yin, T.; Xia, Y.; Zhang, F.; Lian, K.; Liu, Y.; Wang, H.; et al. TXNIP/Redd1 signalling and excessive autophagy: A novel mechanism of myocardial ischaemia/reperfusion injury in mice. Cardiovasc. Res. 2020, 116, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lian, K.; Zhang, L.; Wang, R.; Yi, F.; Gao, C.; Xin, C.; Zhu, D.; Li, Y.; Yan, W.; et al. TXNIP mediates NLRP3 inflammasome activation in cardiac microvascular endothelial cells as a novel mechanism in myocardial ischemia/reperfusion injury. Basic Res. Cardiol. 2014, 109, 415. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Young, M.E.; Chatham, J.C.; Crossman, D.K.; Dell’Italia, L.J.; Shalev, A. TXNIP regulates myocardial fatty acid oxidation via miR-33a signaling. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H64–H75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Yu, Y. The function of thioredoxin-binding protein-2 (TBP-2) in different diseases. Oxidative Med. Cell. Longev. 2018, 2018, 4582130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, A.; Jolibois, J.; Marquet de Rougé, P.; Nivet-Antoine, V. The emerging role of TXNIP in ischemic and cardiovascular diseases; a novel marker and therapeutic target. Int. J. Mol. Sci. 2021, 22, 1693. [Google Scholar] [CrossRef]
- Xiang, G.; Seki, T.; Schuster, M.D.; Witkowski, P.; Boyle, A.J.; See, F.; Martens, T.P.; Kocher, A.; Sondermeijer, H.; Krum, H.; et al. Catalytic degradation of vitamin D up-regulated protein 1 mRNA enhances cardiomyocyte survival and prevents left ventricular remodeling after myocardial ischemia. J. Biol. Chem. 2005, 280, 39394–39402. [Google Scholar] [CrossRef] [Green Version]
- Brondum, E.; Nilsson, H.; Aalkjaer, C. Functional abnormalities in isolated arteries from Goto-Kakizaki and streptozotocin-treated diabetic rat models. Horm. Metab. Res. 2005, 37, 56–60. [Google Scholar] [CrossRef]
- Bond, J.S.; Failla, M.L.; Unger, D.F. Elevated manganese concentration and arginase activity in livers of streptozotocin-induced diabetic rats. J. Biol. Chem. 1983, 258, 8004–8009. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Current Protocol. 2021, 1, e78. [Google Scholar] [CrossRef]
- Avtandilov, G.G. Pathologicoanatomical diagnosis in the light of medical informatics requirements. Arkhiv Patol. 1990, 52, 63–67. [Google Scholar]
- Blagonravov, M.L.; Korshunova, A.Y.; Azova, M.M.; Bondar, S.A.; Frolov, V.A. Cardiomyocyte autophagia and morphological alterations in the left ventricular myocardium during acute focal ischemia. Bull. Exp. Biol. Med. 2016, 160, 398–400. [Google Scholar] [CrossRef]
- Byon, C.H.; Han, T.; Wu, J.; Hui, S.T. Txnip ablation reduces vascular smooth muscle cell inflammation and ameliorates atherosclerosis in apolipoprotein E knockout mice. Atherosclerosis 2015, 241, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Zhang, Y.; Kelly, D.J.; Tan, C.Y.; Gill, A.; Cheng, D.; Braet, F.; Park, J.S.; Sue, C.M.; Pollock, C.A.; et al. Thioredoxin interacting protein (TXNIP) regulates tubular autophagy and mitophagy in diabetic nephropathy through the mTOR signaling pathway. Sci. Rep. 2016, 6, 29196. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Liao, R.; Liu, C.; Liu, S.; Huang, H.; Liu, J.; Jin, T.; Guo, H.; Zheng, Z.; Xia, M.; et al. Epigenetic regulation of TXNIP-mediated oxidative stress and NLRP3 inflammasome activation contributes to SAHH inhibition-aggravated diabetic nephropathy. Redox Biol. 2021, 45, 102033. [Google Scholar] [CrossRef]
- Luo, T.; Zhou, X.; Qin, M.; Lin, Y.; Lin, J.; Chen, G.; Liu, A.; Ouyang, D.; Chen, D.; Pan, H. Corilagin restrains NLRP3 inflammasome activation and pyroptosis through the ROS/TXNIP/NLRP3 pathway to prevent inflammation. Oxid. Med. Cell Longev. 2022, 2022, 1652244. [Google Scholar] [CrossRef]
- An, X.; Zhang, Y.; Cao, Y.; Chen, J.; Qin, H.; Yang, L. Punicalagin protects diabetic nephropathy by inhibiting pyroptosis based on TXNIP/NLRP3 pathway. Nutrients 2020, 12, 1516. [Google Scholar] [CrossRef]
- Ouyang, Y.; Peng, Y.; Li, J.; Holmgren, A.; Lu, J. Modulation of thiol-dependent redox system by metal ions via thioredoxin and glutaredoxin systems. Metallomics 2018, 10, 218–228. [Google Scholar] [CrossRef]
- Ogata, F.T.; Simões Sato, A.Y.; Coppo, L.; Arai, R.J.; Stern, A.I.; Pequeno Monteiro, H. Thiol-based antioxidants and the epithelial/mesenchymal transition in cancer. Antioxid. Redox Signal. 2022, 36, 1037–1050. [Google Scholar] [CrossRef]
- Darenskaya, M.A.; Chugunova, E.V.; Kolesnikov, S.I.; Grebenkina, L.A.; Semenova, N.V.; Nikitina, O.A.; Kolesnikova, L.I. Content of carbonyl compounds and parameters of glutathione metabolism in men with type 1 diabetes mellitus at preclinical stages of diabetic nephropathy. Bull. Exp. Biol. Med. 2021, 171, 592–595. [Google Scholar] [CrossRef]
- Wang, B.F.; Yoshioka, J. The emerging role of thioredoxin-interacting protein in myocardial ischemia/reperfusion injury. J. Cardiovasc. Pharmacol. Ther. 2017, 22, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Ji, L.; Xing, W.; Zhang, W.; Zhou, H.; Qian, X.; Wang, X.; Gao, F.; Sun, X.; Zhang, H. Acute hyperglycaemia enhances oxidative stress and aggravates myocardial ischaemia/reperfusion injury: Role of thioredoxin-interacting protein. J. Cell Mol. Med. 2013, 17, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, J.; Yang, X.; Sun, X.; Xu, Q.; Wang, B.; Zhong, P.; Wei, Z. Altered expression of TXNIP in the peripheral leukocytes of patients with coronary atherosclerotic heart disease. Medicine 2017, 96, e9108. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, C.; Lei, Y.; Wu, W.; Liu, W. Inhibition of EZH2 attenuates coronary heart disease by interacting with microRNA-22 to regulate the TXNIP/nuclear factor-κB pathway. Exp. Physiol. 2020, 105, 2038–2050. [Google Scholar] [CrossRef] [PubMed]
- Devi, T.; Somayajulu, M.; Kowluru, R.; Singh, L.P. TXNIP regulates mitophagy in retinal Müller cells under high-glucose conditions: Implications for diabetic retinopathy. Cell Death Dis. 2017, 8, e2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.J.; Shen, Z.; Cui, R.X.; Huang, Y.; Xu, D.L.; Zhao, F.L.; Pan, J.; Shi, A.M.; Liu, T.; Yu, Y.L. Thioredoxin-interacting protein (TXNIP) regulates Parkin/PINK1-mediated mitophagy in dopaminergic neurons under high-glucose conditions: Implications for molecular links between Parkinson’s disease and diabetes. Neurosci. Bull. 2020, 36, 346–358. [Google Scholar] [CrossRef]
- Rodriguez, Y.A.; Kaur, S.; Nolte, E.; Zheng, Z.; Blagg, B.S.J.; Dobrowsky, R.T. Novologue therapy requires heat shock protein 70 and thioredoxin-interacting protein to improve mitochondrial bioenergetics and decrease mitophagy in diabetic sensory neurons. ACS Chem. Neurosci. 2021, 12, 3049–3059. [Google Scholar] [CrossRef]
- Robaczewska, J.; Kedziora-Kornatowska, K.; Kozakiewicz, M.; Zary-Sikorska, E.; Pawluk, H.; Pawliszak, W.; Kedziora, J. Role of glutathione metabolism and glutathione-related antioxidant defense systems in hypertension. J. Physiol. Pharmacol. 2016, 67, 331–337. [Google Scholar]
- Yu, Q.; Lee, C.F.; Wang, W.; Karamanlidis, G.; Kuroda, J.; Matsushima, S.; Sadoshima, J.; Tian, R. Elimination of NADPH oxidase activity promotes reductive stress and sensitizes the heart to ischemic injury. J. Am. Heart Assoc. 2014, 3, e000555. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.; Li, B.; Wang, W.; Liu, X.; Xia, Y.; Zhang, C.; Zhang, M.; Zhang, Y.; An, F. NLRP3 gene silencing ameliorates diabetic cardiomyopathy in a type 2 diabetes rat model. PLoS ONE 2014, 9, e104771. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Han, B.; Zhang, B.; Chu, Z.; Zhang, X.; Lu, Q.; Han, J. Schisandra chinensis polysaccharides prevent cardiac hypertrophy by dissociating thioredoxin-interacting protein/thioredoxin-1 complex and inhibiting oxidative stress. Biomed. Pharmacother. 2021, 139, 111688. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sklifasovskaya, A.; Blagonravov, M.; Azova, M.; Goryachev, V. Myocardial Glutathione Synthase and TRXIP Expression Are Significantly Elevated in Hypertension and Diabetes: Influence of Stress on Antioxidant Pathways. Pathophysiology 2023, 30, 248-259. https://doi.org/10.3390/pathophysiology30020021
Sklifasovskaya A, Blagonravov M, Azova M, Goryachev V. Myocardial Glutathione Synthase and TRXIP Expression Are Significantly Elevated in Hypertension and Diabetes: Influence of Stress on Antioxidant Pathways. Pathophysiology. 2023; 30(2):248-259. https://doi.org/10.3390/pathophysiology30020021
Chicago/Turabian StyleSklifasovskaya, Anastasia, Mikhail Blagonravov, Madina Azova, and Vyacheslav Goryachev. 2023. "Myocardial Glutathione Synthase and TRXIP Expression Are Significantly Elevated in Hypertension and Diabetes: Influence of Stress on Antioxidant Pathways" Pathophysiology 30, no. 2: 248-259. https://doi.org/10.3390/pathophysiology30020021
APA StyleSklifasovskaya, A., Blagonravov, M., Azova, M., & Goryachev, V. (2023). Myocardial Glutathione Synthase and TRXIP Expression Are Significantly Elevated in Hypertension and Diabetes: Influence of Stress on Antioxidant Pathways. Pathophysiology, 30(2), 248-259. https://doi.org/10.3390/pathophysiology30020021