Thermogravimetric, Devolatilization Rate, and Differential Scanning Calorimetry Analyses of Biomass of Tropical Plantation Species of Costa Rica Torrefied at Different Temperatures and Times



Abstract
:1. Introduction
2. Material and Methods
2.1. Material Characteristics
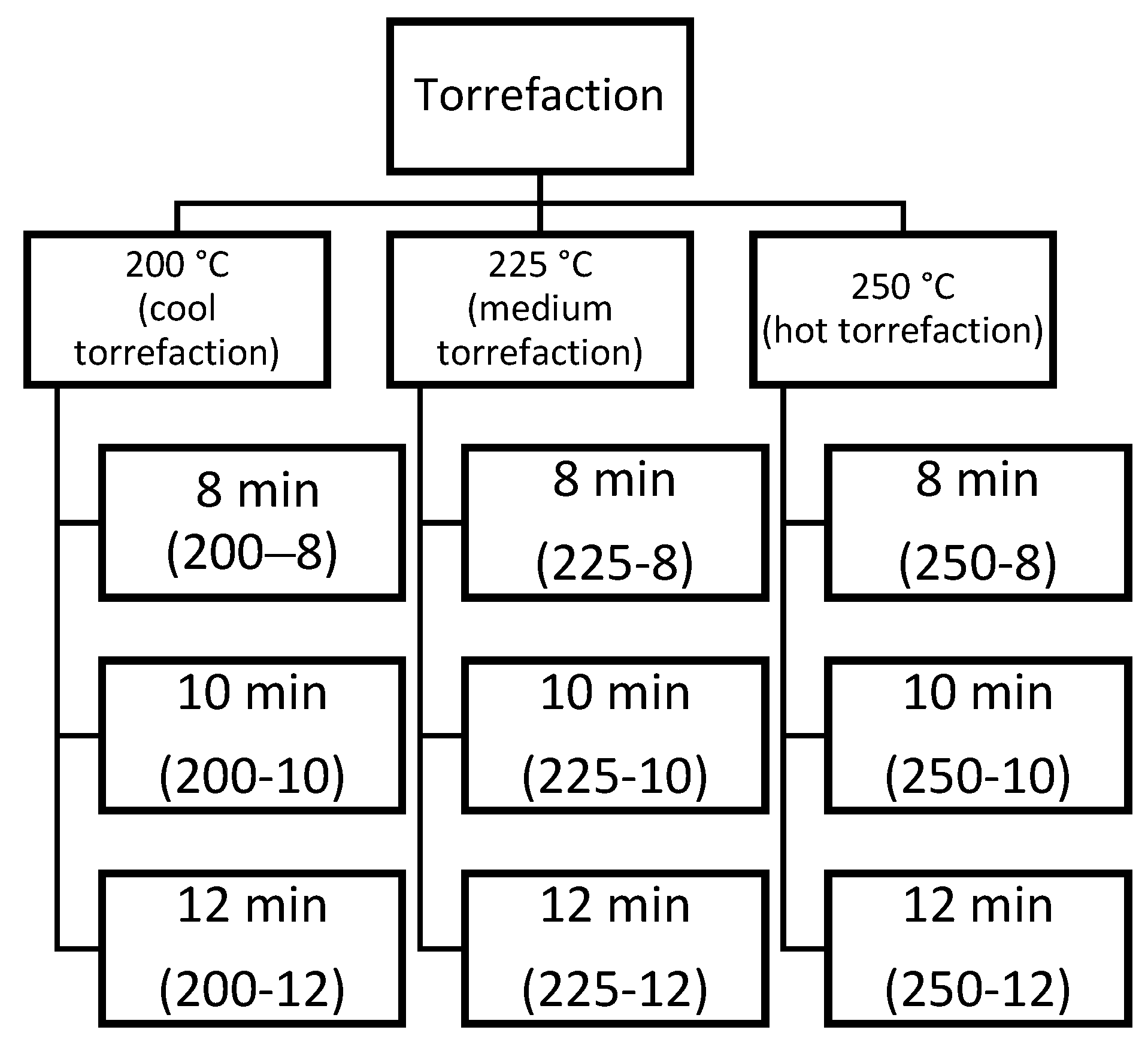
2.2. Torrefaction Process
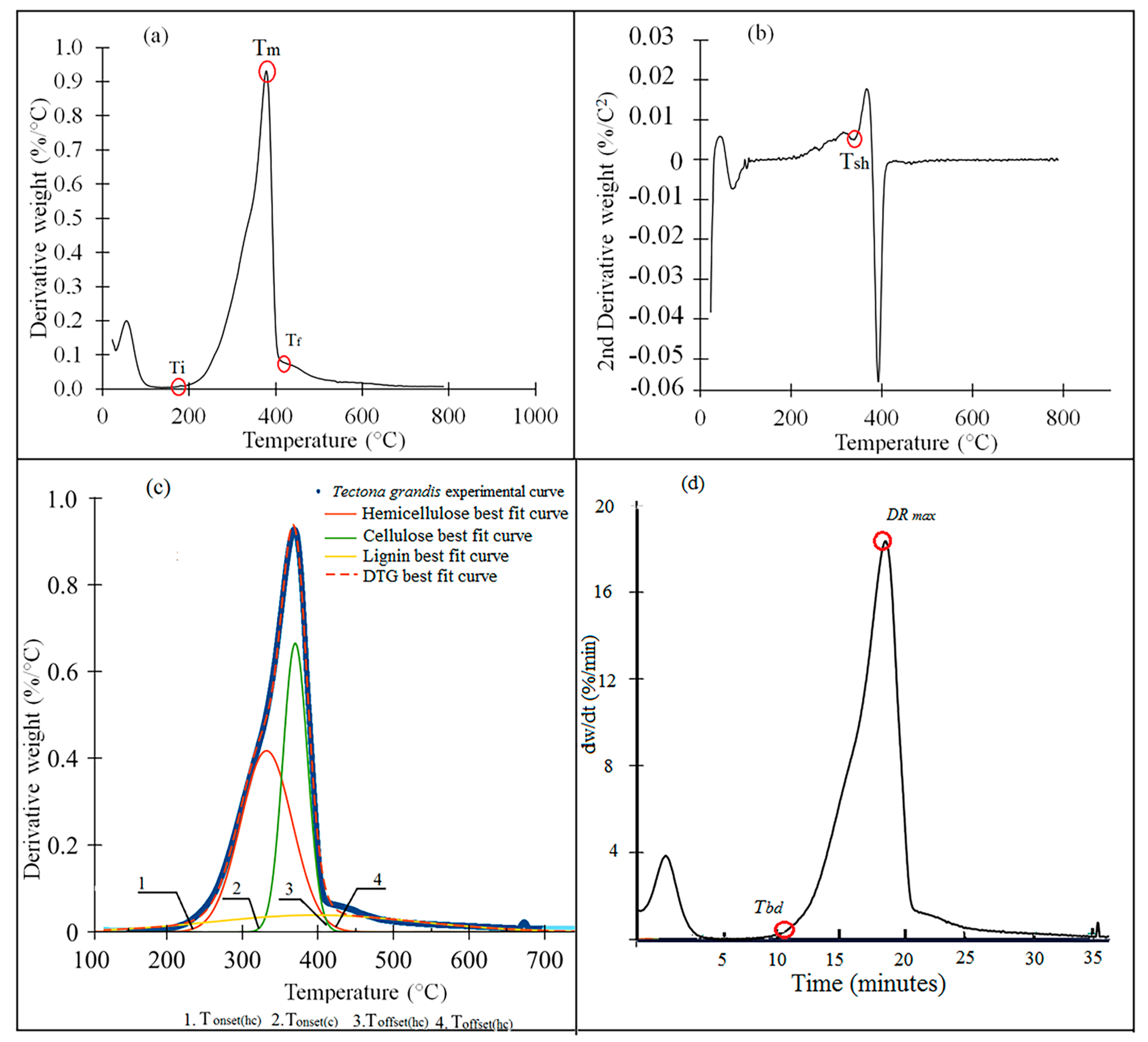
2.3. Thermogravimetric Analysis (TGA)
2.4. Devolatilization Variation
2.5. Statistical Analysis
3. Results
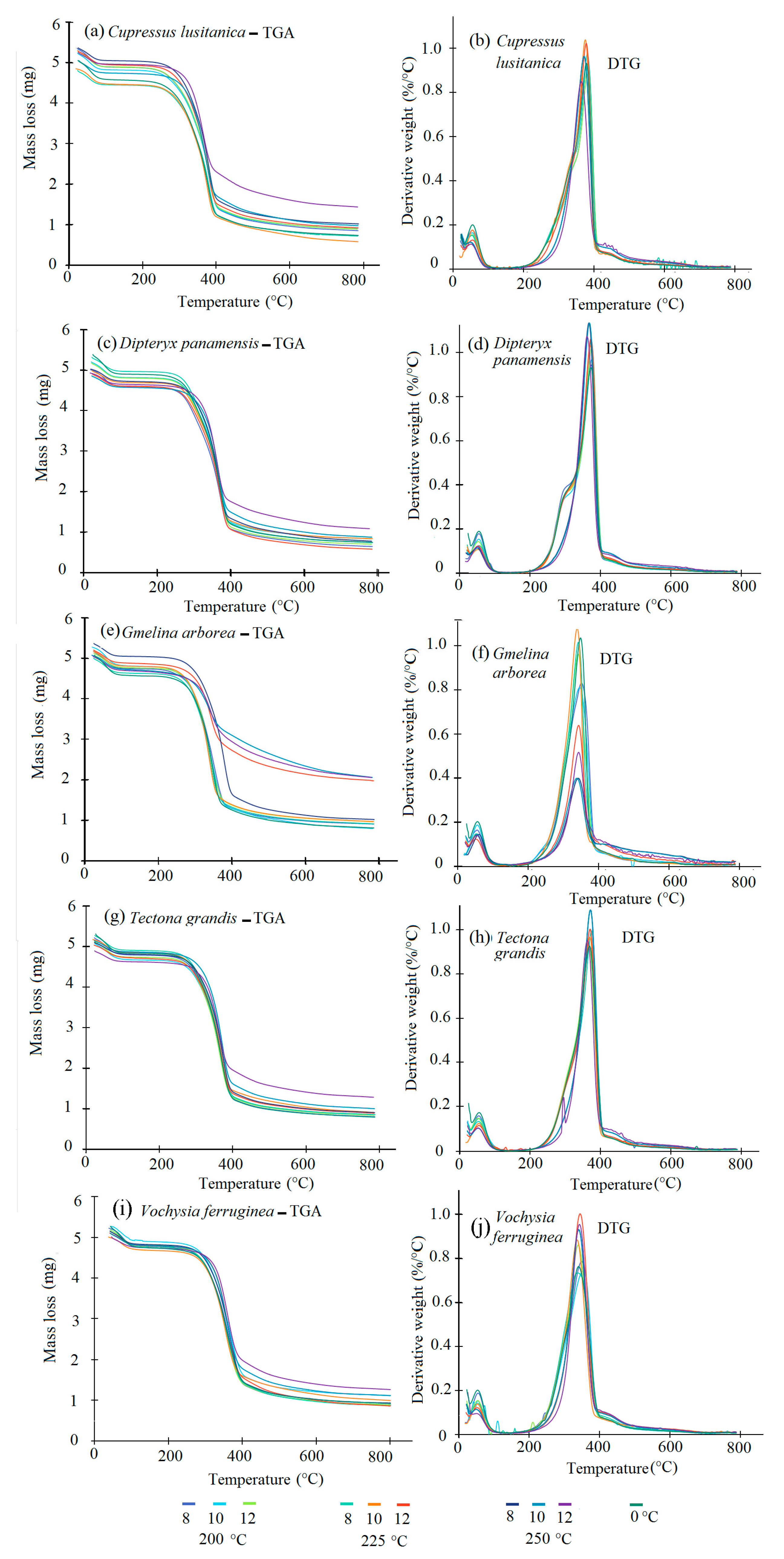
3.1. TGA-DTG Analysis
3.2. Devolatilization
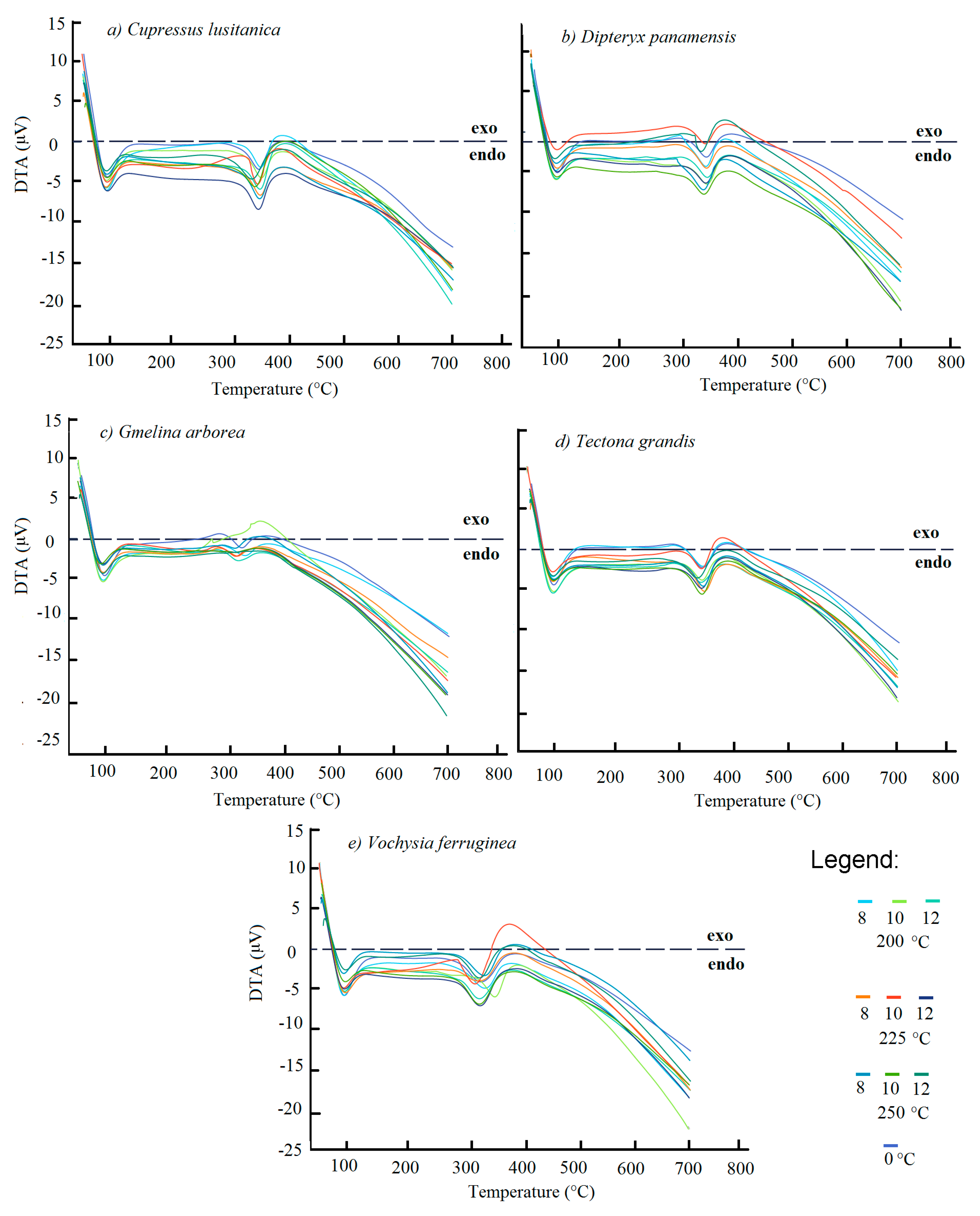
3.3. Differential Scanning Calorimetry Analyses
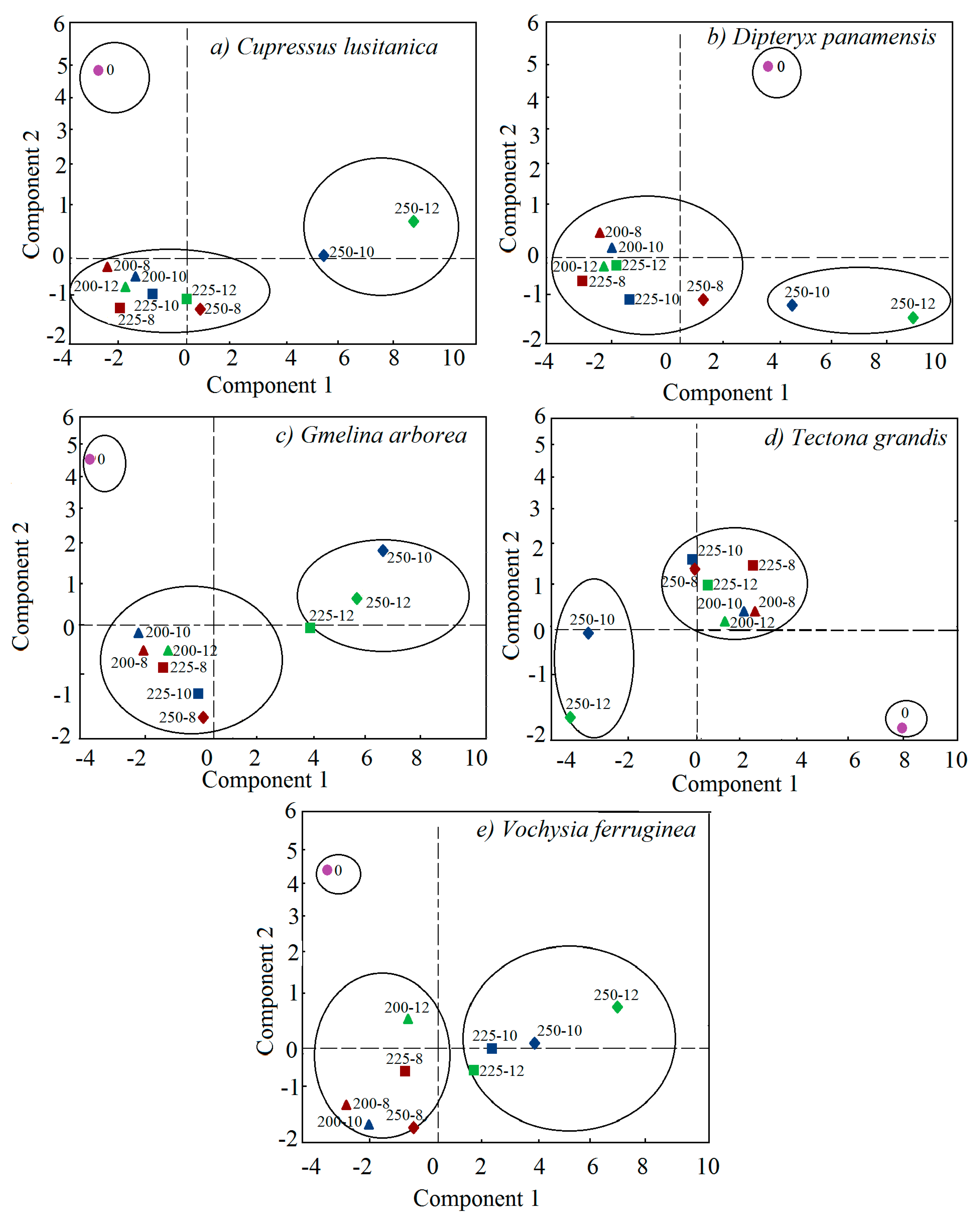
3.4. Multivariate Analysis
4. Discussion
4.1. TGA-DTG Analysis
4.2. Devolatilization
4.3. Differential Scanning Calorimetry Analyses
4.4. Multivariate Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yue, Y.; Singh, H.; Singh, B.; Mani, S. Torrefaction of sorghum biomass to improve fuel properties. Bioresour. Technol. 2017, 232, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Puig-Arnavat, M.; Shang, L.; Sárossy, Z.; Ahrenfeldt, J.; Henriksen, U.B. From a single pellet press to a bench scale pellet mill—Pelletizing six different biomass feedstocks. Fuel Process. Technol. 2016, 142, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Eseltine, D.; Thanapal, S.S.; Annamalai, K.; Ranjan, D. Torrefaction of woody biomass (Juniper and Mesquite) using inert and non-inert gases. Fuel 2013, 113, 379–388. [Google Scholar] [CrossRef]
- Ren, S.; Lei, H.; Wang, L.; Bu, Q.; Chen, S.; Wu, J. Thermal behaviour and kinetic study for woody biomass torrefaction and torrefied biomass pyrolysis by TGA. Biosyst. Eng. 2013, 116, 420–426. [Google Scholar] [CrossRef]
- Chen, W.H.; Peng, J.; Bi, X.T. A state-of-the-art review of biomass torrefaction, densification and applications. Renew. Sustain. Energy Rev. 2015, 44, 847–866. [Google Scholar] [CrossRef]
- Medic, D.; Darr, M.; Shah, A.; Potter, B.; Zimmerman, J. Effects of torrefaction process parameters on biomass feedstock upgrading. Fuel 2012, 91, 47–154. [Google Scholar] [CrossRef]
- Wang, G.; Luo, Y.; Deng, J.; Kuang, J.; Zhang, Y. Pretreatment of biomass by torrefaction. Chin. Sci. Bull. 2011, 56, 1442–1448. [Google Scholar] [CrossRef]
- Peng, J.H.; Bi, X.T.; Sokhansanj, S.; Lim, C.J. Torrefaction and densification of different species of softwood residues. Fuel 2013, 111, 411–421. [Google Scholar] [CrossRef]
- Van der Stelt, M.J.C.; Gerhauser, H.; Kiel, J.H.A.; Ptasinski, K.J. Biomass upgrading by torrefaction for the production of biofuels: A review. Biomass Bioenergy 2011, 35, 3748–3762. [Google Scholar] [CrossRef]
- Chew, J.J.; Doshi, V. Recent advances in biomass pretreatment–Torrefaction fundamentals and technology. Renew. Sustain. Energy Rev. 2011, 15, 4212–4222. [Google Scholar] [CrossRef]
- Ciolkosz, D.; Wallace, R. A review of torrefaction for bioenergy feedstock production. Biofuel Bioprod. Biorefin. 2011, 5, 317–329. [Google Scholar] [CrossRef]
- Gaitán-Álvarez, J.; Moya, R.; Rodríguez-Zúñiga, A.; Puente-Urbina, A. Characterization of torrefied biomass of five reforestation species (Cupressus lusitanica, Dipteryx panamensis, Gmelina arborea, Tectona grandis and Vochysia ferruginea) in Costa Rica. Bioresources 2017, 12, 7566–7589. [Google Scholar] [CrossRef]
- Da Silva, C.M.S.; Carneiro, A.D.; Pereira, B.L.C.; Vital, B.R.; Alves, I.C.N.; de Magalhaes, M.A. Stability to thermal degradation and chemical composition of woody biomass subjected to the torrefaction process. Eur. J. Wood Prod. 2016, 74, 845–850. [Google Scholar] [CrossRef]
- Korošec, R.C.; Lavrič, B.; Rep, G.; Pohleven, F.; Bukovec, P. Thermogravimetry as a possible tool for determining modification degree of thermally treated Norway spruce wood. J. Therm. Anal. Calorim. 2009, 98, 189. [Google Scholar] [CrossRef]
- Aydemir, D.; Gunduz, G.; Altuntas, E.; Ertas, M.; Sahin, H.T.; Alma, M.H. Investigating changes in the chemical constituents and dimensional stability of heat-treated hornbeam and uludag fir wood. BioResources 2011, 6, 1308–1321. [Google Scholar]
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G. An overview of the chemical composition of biomass. Fuel 2010, 89, 913–933. [Google Scholar] [CrossRef]
- Poudel, J.; Ohm, T.I.; Oh, S.C. A study on torrefaction of food waste. Fuel 2015, 140, 275–281. [Google Scholar] [CrossRef]
- Moya, R.; Rodríguez-Zúñiga, A.; Puente-Urbina, A. Thermogravimetric and devolatilisation analysis for five plantation species: Effect of extractives, ash compositions, chemical compositions and energy parameters. Thermochim. Acta 2017, 647, 36–46. [Google Scholar] [CrossRef]
- Gaitán-Álvarez, J.; Moya, R. Characteristics and properties of pellet fabricated with torrefaccioned biomass of Gmelina arborea and Dipterix panamensis at different time. Revista Chapingo 2016, 23, 325–337. [Google Scholar]
- Tenorio, C.; Moya, R. Thermogravimetric characteristics, its relation with extractives and chemical properties and combustion characteristics of ten fast-growth species in Costa Rica. Thermochim. Acta 2013, 563, 12–21. [Google Scholar] [CrossRef]
- Gaitán-Álvarez, J.; Moya, R.; Puente-Urbina, A.; Rodríguez-Zúñiga, A. Physical and compression properties of pellets manufactured with the biomass of five woody tropical species of Costa Rica torrefied at different temperatures and times. Energies 2017, 10, 1205. [Google Scholar] [CrossRef]
- Aragón-Garita, S.; Moya, R.; Bond, B.; Valaert, J.; Tomazello Filho, M. Production and quality analysis of pellets manufactured from five potential energy crops in the Northern Region of Costa Rica. Biomass Bioenergy 2016, 87, 84–95. [Google Scholar] [CrossRef]
- Tenorio, C.; Moya, R.; Tomazello Filho, M.; Valaert, J. Application of the X-ray densitometry in the evaluation of the quality and mechanical properties of biomass pellets. Fuel Process. Technol. 2015, 132, 62–73. [Google Scholar] [CrossRef]
- Tenorio, C.; Moya, R.; Tomazello-Filho, M.; Valaert, J. Quality of pellets made from agricultural and forestry crops in Costa Rican tropical climates. BioResources 2014, 9, 482–498. [Google Scholar] [CrossRef]
- Moya, R.; Rodriguez-Zuñiga, A.; Puente-Urbina, A.; Gaitan-Alvarez, J. Study of light, middle and severe torrefaction and effects of extractives and chemical compositions on torrefaction process by thermogravimetric analyses in five fast-growing plantation of Costa Rica. Energy 2018, 149, 152–160. [Google Scholar] [CrossRef]
- Sbirrazzuoli, N.; Vyazovkin, S.; Mititelu, A.; Sladic, C.; Vincent, L. A Study of Epoxy-Amine Cure Kinetics by Combining Isoconversional Analysis with Temperature Modulated DSC and Dynamic Rheometry. Macromol. Chem. Phys. 2003, 204, 1815–1821. [Google Scholar] [CrossRef]
- Grønli, M.G.; Várhegyi, G.; Di Blasi, C. Thermogravimetric analysis and devolatilization kinetics of wood. Ind. Eng. Chem. Res. 2002, 41, 4201–4208. [Google Scholar] [CrossRef]
- Prins, M.J.; Ptasinski, K.J.; Janssen, F.J. Torrefaction of wood: Part 2. Analysis of products. J. Anal. Appl. Pyrolysis 2006, 77, 35–40. [Google Scholar] [CrossRef]
- Arias, B.; Pevida, C.; Fermoso, J.; Plaza, M.G.; Rubiera, F.; Pis, J.J. Influence of torrefaction on the grindability and reactivity of woody biomass. Fuel Process. Technol. 2008, 89, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Ramiah, M.V. Thermogravimetric and differential thermal analysis of cellulose, hemicellulose, and lignin. J. Appl. Polym. Sci. 1970, 14, 1323–1337. [Google Scholar] [CrossRef]
- Bach, Q.V.; Skreiberg, Ø. Upgrading biomass fuels via wet torrefaction: A review and comparison with dry torrefaction. Renew. Sustain. Energy Rev. 2016, 54, 665–677. [Google Scholar] [CrossRef]
- Yang, H.; Yan, R.; Chen, H.; Lee, D.H.; Zheng, C. Characteristics of hemicellulose, cellulose and lignin pyrolysis. Fuel 2007, 86, 1781–1788. [Google Scholar] [CrossRef]
- Avni, E.; Coughlin, R.W. Kinetic analysis of lignin pyrolysis using non-isothermal TGA data. Thermochim. Acta 1985, 90, 157–167. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kang, I.A.; Doh, G.H.; Kim, W.J.; Kim, J.S.; Yoon, H.G.; Wu, Q. Thermal, mechanical and morphological properties of polypropylene/clay/wood flour nanocomposites. Express Polym. Lett. 2008, 2, 78–87. [Google Scholar] [CrossRef]
- Islam, M.S.; Hamdan, S.; Rahman, M.R.; Jusoh, I.; Ahmen, A.S. Dynamic Young’s modulus, morphological, and thermal stability of 5 tropical light hardwoods modified by benzene diazonium salt treatment. BioResources 2011, 6, 737–750. [Google Scholar]
- Arteaga-Pérez, L.E.; Grandón, H.; Flores, M.; Segura, C.; Kelley, S.S. Steam torrefaction of Eucalyptus globulus for producing black pellets: A pilot-scale experience. Bioresour. Technol. 2017, 238, 194–204. [Google Scholar] [CrossRef] [PubMed]
- McKendry, P. Energy production from biomass (part 2): Conversion technologies. Bioresour. Technol. 2002, 83, 47–54. [Google Scholar] [CrossRef]
- Doddapaneni, T.R.; Konttinen, J.; Hukka, T.I.; Moilanen, A. Influence of torrefaction pretreatment on the pyrolysis of Eucalyptus clone: A study on kinetics, reaction mechanism and heat flow. Ind. Crops Prod. 2016, 92, 244–254. [Google Scholar] [CrossRef]
- Chen, W.H.; Kuo, P.C. Torrefaction and co-torrefaction characterization of hemicellulose, cellulose and lignin as well as torrefaction of some basic constituents in biomass. Energy 2011, 36, 803–811. [Google Scholar] [CrossRef]
- Park, J.; Meng, J.; Lim, K.H.; Rojas, O.J.; Park, S. Transformation of lignocellulosic biomass during torrefaction. J. Anal. Appl. Pyrolysis 2013, 100, 199–206. [Google Scholar] [CrossRef]
- Ramos, L.P. The chemistry involved in the steam treatment of lignocellulosic materials. Quim. Nova 2003, 26, 863–871. [Google Scholar] [CrossRef]
- Biswas, A.K.; Umeki, K.; Yang, W.; Blasiak, W. Change of pyrolysis characteristics and structure of woody biomass due to steam explosion pretreatment. Fuel Process. Technol. 2011, 92, 1849–1854. [Google Scholar] [CrossRef]
- Bach, Q.V.; Tran, K.Q.; Skreiberg, Ø.; Khalil, R.A.; Phan, A.N. Effects of wet torrefaction on reactivity and kinetics of wood under air combustion conditions. Fuel 2014, 137, 375–383. [Google Scholar] [CrossRef]
- Bobleter, O. Hydrothermal degradation of polymers derived from plants. Prog. Polym. Sci. 1994, 19, 797–841. [Google Scholar] [CrossRef]
- Wyman, C.E.; Decker, S.R.; Himmel, M.E.; Brady, J.W.; Skopec, C.E.; Viikari, L. Hydrolysis of cellulose and hemicellulose. Polysaccharides 2005, 1, 1023–1062. [Google Scholar]
- Skreiberg, A.; Skreiberg, Ø.; Sandquist, J.; Sørum, L. TGA and macro-TGA characterisation of biomass fuels and fuel mixtures. Fuel 2011, 90, 2182–2197. [Google Scholar] [CrossRef]
- Ball, R.; McIntosh, A.C.; Brindley, J. Feedback processes in cellulose thermal decomposition: Implications for fire-retarding strategies and treatments. Combust. Theory Model. 2004, 8, 281–291. [Google Scholar] [CrossRef]
- Shen, J.; Igathinathane, C.; Yu, M.; Pothula, A.K. Biomass pyrolysis and combustion integral and differential reaction heats with temperatures using thermogravimetric analysis/differential scanning calorimetry. Bioresour. Technol. 2015, 185, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Stenseng, M.; Jensen, A.; Dam-Johansen, K. Investigation of biomass pyrolysis by thermogravimetric analysis and differential scanning calorimetry. J. Anal. Appl. Pyrolysis 2001, 58, 765–780. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Cupressus lusitanica | Dipterix panamensis | Gmelina arborea | Tectona grandis | Vochysia ferruginea |
|---|---|---|---|---|---|
| Cellulose (%) | 64.7 | 49.9 | 55.6 | 54.4 | 50.9 |
| Lignin (%) | 31.4 | 20.3 | 24.2 | 21.90 | 11.2 |
| Ash (%) | 0.18 | 3.04 | 0.96 | 2.81 | 0.99 |
| Carbon (%) | 50.18 | 48.64 | 48.39 | 49.77 | 49.32 |
| Nitrogen (%) | 0.27 | 0.24 | 0.20 | 0.20 | 0.27 |
| Species | Temperature (°C) | Time (min) | Ti (°C) | Tf (°C) | Residual Mass (%) | Tonset(hc) (°C) | Toffset(hc) (°C) | Tsh (°C) | Tonset(c) (°C) | Toffset(c) (°C) | Tm (°C) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cupressus lusitanica | 0 | 0 | 172.1 | 448.7 | 21.0 | 221.2 | 455.4 | 339.3 | 253.8 | 465.8 | 378.8 |
| 200 | 8 | 177.2 | 449.8 | 22.6 | 231.2 | 454.4 | 345.3 | 345.5 | 418.7 | 383.5 | |
| 10 | 181.7 | 438.0 | 24.1 | 234.0 | 452.3 | 340.1 | 346.4 | 418.1 | 383.5 | ||
| 12 | 191.7 | 453.5 | 23.3 | 231.4 | 454.5 | 345.3 | 346.1 | 418.4 | 382.6 | ||
| 225 | 8 | 215.4 | 436.2 | 22.8 | 230.9 | 452.4 | 346.5 | 344.6 | 416.4 | 380.8 | |
| 10 | 183.5 | 412.6 | 23.6 | 237.1 | 438.2 | 329.7 | 340.1 | 409.7 | 375.3 | ||
| 12 | 173.5 | 445.3 | 25.0 | 245.0 | 439.1 | 337.5 | 341.2 | 413.6 | 378.0 | ||
| 250 | 8 | 161.7 | 437.1 | 28.3 | 247.9 | 440.9 | 342.7 | 343.4 | 415.7 | 379.8 | |
| 10 | 172.6 | 425.3 | 31.3 | 266.5 | 425.7 | 263.5 | 333.5 | 413.8 | 371.7 | ||
| 12 | 213.8 | 476.6 | 36.1 | 273.5 | 415.8 | 275.2 | 323.3 | 410.7 | 364.4 | ||
| Dipteryx panamensis | 0 | 0 | 146.3 | 460.9 | 18.7 | 239.5 | 375.4 | 315.0 | 238.2 | 468.2 | 372.1 |
| 200 | 8 | 194.7 | 431.1 | 19.6 | 206.9 | 438.6 | 310.2 | 330.3 | 413.0 | 373.8 | |
| 10 | 188.2 | 441.3 | 21.5 | 206.4 | 443.0 | 307.6 | 330.4 | 416.7 | 375.1 | ||
| 12 | 201.2 | 425.7 | 20.2 | 205.2 | 443.1 | 306.3 | 332.4 | 415.6 | 375.1 | ||
| 225 | 8 | 212.8 | 430.9 | 20.8 | 211.6 | 440.9 | 316.7 | 333.0 | 411.3 | 373.8 | |
| 10 | 205.1 | 433.5 | 23.4 | 211.0 | 437.9 | 314.1 | 331.2 | 409.2 | 372.5 | ||
| 12 | 206.4 | 446.5 | 18.5 | 212.4 | 441.3 | 311.5 | 331.2 | 411.9 | 372.5 | ||
| 250 | 8 | 212.8 | 433.5 | 24.1 | 237.6 | 428.4 | 308.9 | 325.7 | 409.3 | 367.3 | |
| 10 | 206.4 | 438.7 | 27.2 | 263.6 | 420.2 | 272.6 | 329.8 | 407.9 | 368.6 | ||
| 12 | 196.0 | 424.4 | 33.5 | 266.1 | 413.8 | 272.6 | 232.8 | 404.0 | 362.1 | ||
| Gmelina arborea | 0 | 0 | 172.1 | 471.5 | 20.8 | 249.1 | 385.0 | 305.8 | 258.7 | 417.9 | 349.9 |
| 200 | 8 | 194.7 | 410.2 | 24.4 | 196.7 | 392.7 | 247.9 | 305.6 | 395.6 | 351.7 | |
| 10 | 197.3 | 401.1 | 24.4 | 225.1 | 410.4 | 247.9 | 301.7 | 401.6 | 346.5 | ||
| 12 | 221.9 | 419.2 | 25.5 | 217.9 | 417.2 | 302.4 | 295.9 | 391.7 | 341.4 | ||
| 225 | 8 | 201.2 | 399.8 | 26.6 | 227.0 | 411.4 | 303.7 | 305.3 | 381.4 | 341.4 | |
| 10 | 199.9 | 406.3 | 26.4 | 236.3 | 391.9 | 240.1 | 303.9 | 376.0 | 338.8 | ||
| 12 | 207.7 | 424.4 | 50.0 | 238.5 | 418.1 | 262.2 | 305.2 | 380.7 | 344.0 | ||
| 250 | 8 | 173.9 | 454.3 | 25.8 | 247.9 | 440.9 | 267.4 | 343.4 | 415.7 | 341.4 | |
| 10 | 198.6 | 442.6 | 56.9 | 236.6 | 416.5 | 242.7 | 295.0 | 384.6 | 337.5 | ||
| 12 | 179.1 | 412.8 | 56.9 | 188.0 | 495.9 | 255.7 | 294.7 | 386.6 | 344.0 | ||
| Tectona grandis | 0 | 0 | 164.5 | 473.0 | 19.2 | 262.6 | 398.5 | 321.0 | 257.8 | 454.6 | 368.2 |
| 200 | 8 | 233.6 | 425.7 | 22.1 | 226.6 | 435.2 | 312.8 | 322.0 | 418.8 | 371.2 | |
| 10 | 219.3 | 430.9 | 23.2 | 233.0 | 433.8 | 320.6 | 330.2 | 410.6 | 369.9 | ||
| 12 | 227.1 | 425.7 | 23.1 | 225.7 | 434.4 | 316.7 | 320.4 | 413.4 | 366.0 | ||
| 225 | 8 | 198.6 | 438.7 | 22.1 | 227.2 | 441.1 | 312.8 | 320.4 | 413.4 | 373.8 | |
| 10 | 220.6 | 423.1 | 26.5 | 233.0 | 430.8 | 312.8 | 329.1 | 409.6 | 369.9 | ||
| 12 | 233.6 | 443.9 | 24.2 | 232.0 | 434.8 | 316.7 | 328.6 | 411.9 | 369.9 | ||
| 250 | 8 | 212.8 | 432.2 | 25.1 | 240.2 | 434.9 | 308.9 | 330.8 | 414.9 | 373.8 | |
| 10 | 241.4 | 427.0 | 28.8 | 266.0 | 422.4 | 286.8 | 331.9 | 412.0 | 369.9 | ||
| 12 | 272.6 | 430.9 | 36.9 | 256.6 | 416.1 | 298.5 | 317.6 | 408.6 | 363.4 | ||
| Vochysia ferruginea | 0 | 0 | 161.5 | 439.6 | 22.3 | 233.7 | 369.6 | 301.3 | 227.2 | 442.9 | 339.3 |
| 200 | 8 | 219.3 | 427.0 | 22.5 | 220.1 | 414.8 | 301.1 | 301.9 | 407.7 | 350.4 | |
| 10 | 233.6 | 430.9 | 26.2 | 222.0 | 419.1 | 306.3 | 306.3 | 408.1 | 355.6 | ||
| 12 | 223.2 | 424.4 | 22.9 | 217.1 | 384.1 | 299.8 | 282.2 | 405.5 | 338.8 | ||
| 225 | 8 | 224.5 | 420.5 | 23.7 | 222.3 | 415.6 | 255.7 | 297.9 | 408.4 | 337.5 | |
| 10 | 224.5 | 401.1 | 29.6 | 214.5 | 407.1 | 298.5 | 286.1 | 394.1 | 336.2 | ||
| 12 | 251.8 | 432.2 | 24.8 | 237.9 | 401.2 | 245.3 | 294.2 | 400.5 | 346.5 | ||
| 250 | 8 | 245.3 | 415.4 | 24.7 | 224.4 | 414.8 | 297.2 | 303.4 | 406.7 | 350.4 | |
| 10 | 245.1 | 420.5 | 30.4 | 228.4 | 401.8 | 253.1 | 288.4 | 398.4 | 340.1 | ||
| 12 | 227.1 | 414.1 | 35.3 | 251.7 | 395.4 | 268.7 | 294.8 | 395.7 | 342.7 |
| Specie | Temperature (°C) | Time (min) | WTi (%) | WTf (%) | WTonset(hc) (%) | WToffset (hc) (%) | WTsh (%) | WTonset(c) (%) | WToffset(c) (%) | WTm (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Cupressus lusitanica | 0 | 0 | 90.3 | 21.0 | 89.7 | 20.6 | 65.4 | 88.1 | 20.0 | 38.5 |
| 200 | 8 | 91.2 | 22.6 | 90.4 | 22.3 | 65.3 | 65.3 | 24.7 | 39.8 | |
| 10 | 91.8 | 24.1 | 91.0 | 23.3 | 68.5 | 65.7 | 25.6 | 40.5 | ||
| 12 | 92.0 | 23.3 | 91.3 | 23.2 | 66.2 | 65.7 | 25.7 | 41.7 | ||
| 225 | 88 | 92.0 | 22.8 | 91.7 | 21.7 | 65.1 | 66.1 | 24.2 | 40.9 | |
| 10 | 92.0 | 23.6 | 91.1 | 21.8 | 71.9 | 66.5 | 23.9 | 40.4 | ||
| 12 | 93.3 | 25.0 | 92.3 | 25.5 | 72.9 | 71.0 | 27.5 | 43.8 | ||
| 250 | 8 | 94.0 | 28.3 | 92.8 | 26.8 | 72.1 | 71.6 | 28.8 | 42.5 | |
| 10 | 93.7 | 31.3 | 92.1 | 31.3 | 92.3 | 79.4 | 32.5 | 52.7 | ||
| 12 | 94.1 | 36.1 | 92.7 | 42.1 | 92.6 | 86.1 | 42.7 | 62.2 | ||
| Dipteryx panamensis | 0 | 0 | 90.9 | 18.7 | 89.6 | 35.5 | 72.6 | 89.7 | 18.4 | 38.3 |
| 200 | 8 | 91.4 | 19.6 | 91.3 | 19.3 | 74.0 | 65.6 | 20.6 | 36.5 | |
| 10 | 92.4 | 21.5 | 92.3 | 21.4 | 78.1 | 69.4 | 22.8 | 39.6 | ||
| 12 | 92.6 | 20.2 | 92.6 | 19.2 | 77.8 | 67.6 | 20.8 | 37.7 | ||
| 225 | 8 | 93.1 | 20.8 | 93.1 | 20.2 | 21.7 | 68.4 | 22.1 | 39.6 | |
| 10 | 93.8 | 23.4 | 93.8 | 23.2 | 77.0 | 70.0 | 25.0 | 41.1 | ||
| 12 | 93.4 | 18.5 | 93.3 | 18.8 | 77.4 | 69.1 | 20.8 | 39.5 | ||
| 250 | 8 | 93.4 | 24.1 | 92.8 | 24.4 | 82.9 | 76.8 | 25.9 | 46.4 | |
| 10 | 93.8 | 27.2 | 92.3 | 28.8 | 91.7 | 83.8 | 30.1 | 50.9 | ||
| 12 | 93.9 | 33.5 | 92.3 | 34.5 | 92.0 | 93.4 | 35.3 | 56.4 | ||
| Gmelina arborea | 0 | 0 | 89.8 | 20.8 | 88.0 | 26.0 | 76.1 | 87.2 | 23.5 | 44.1 |
| 200 | 8 | 91.4 | 24.4 | 91.4 | 25.6 | 89.2 | 76.4 | 25.4 | 45.9 | |
| 10 | 90.8 | 24.4 | 90.0 | 23.8 | 88.4 | 75.7 | 24.4 | 46.1 | ||
| 12 | 92.3 | 25.5 | 92.4 | 25.6 | 78.4 | 81.3 | 27.5 | 51.0 | ||
| 225 | 8 | 92.2 | 26.6 | 91.7 | 25.7 | 77.2 | 76.3 | 28.2 | 49.5 | |
| 10 | 92.5 | 26.4 | 91.5 | 27.5 | 91.3 | 76.9 | 29.1 | 49.0 | ||
| 12 | 93.0 | 50.0 | 92.3 | 50.6 | 91.1 | 85.1 | 54.8 | 67.5 | ||
| 250 | 8 | 93.9 | 25.8 | 92.8 | 26.8 | 91.6 | 71.6 | 28.8 | 72.5 | |
| 10 | 92.1 | 56.9 | 91.1 | 59.4 | 90.9 | 86.5 | 62.7 | 74.4 | ||
| 12 | 92.4 | 56.9 | 92.3 | 50.0 | 90.5 | 87.5 | 60.0 | 72.2 | ||
| Tectona grandis | 0 | 0 | 91.0 | 19.2 | 88.8 | 23.9 | 74.4 | 89.2 | 19.9 | 43.2 |
| 200 | 8 | 91.1 | 22.1 | 91.3 | 21.5 | 78.5 | 74.5 | 22.5 | 41.5 | |
| 10 | 91.5 | 23.2 | 91.2 | 23.0 | 76.0 | 71.7 | 24.5 | 44.1 | ||
| 12 | 91.9 | 23.1 | 91.9 | 22.5 | 77.1 | 75.4 | 23.9 | 44.2 | ||
| 225 | 8 | 92.7 | 22.1 | 92.4 | 22.0 | 79.7 | 76.7 | 23.7 | 43.1 | |
| 10 | 93.1 | 26.5 | 92.9 | 26.1 | 80.4 | 73.3 | 27.4 | 45.2 | ||
| 12 | 92.7 | 24.2 | 92.7 | 24.7 | 79.5 | 74.7 | 26.3 | 46.2 | ||
| 250 | 8 | 93.7 | 25.1 | 93.2 | 25.0 | 84.3 | 76.3 | 26.5 | 45.5 | |
| 10 | 93.6 | 28.8 | 92.6 | 29.3 | 91.0 | 80.6 | 30.2 | 52.9 | ||
| 12 | 92.6 | 36.9 | 93.2 | 38.3 | 89.6 | 86.5 | 39.1 | 58.8 | ||
| Vochysia ferruginea | 0 | 0 | 89.6 | 22.3 | 88.1 | 32.3 | 74.9 | 88.3 | 22.2 | 52.1 |
| 200 | 8 | 90.2 | 22.5 | 90.2 | 23.4 | 76.0 | 76.0 | 23.9 | 45.7 | |
| 10 | 90.8 | 26.2 | 91.2 | 26.9 | 76.1 | 76.1 | 27.6 | 46.0 | ||
| 12 | 91.0 | 22.9 | 91.2 | 26.4 | 76.9 | 83.2 | 24.3 | 52.0 | ||
| 225 | 8 | 91.4 | 23.7 | 91.4 | 24.1 | 89.3 | 78.2 | 24.7 | 53.6 | |
| 10 | 92.3 | 29.6 | 92.6 | 29.2 | 79.6 | 84.0 | 30.2 | 54.6 | ||
| 12 | 91.7 | 24.8 | 92.4 | 27.7 | 92.0 | 85.9 | 27.9 | 52.8 | ||
| 250 | 8 | 91.5 | 24.7 | 92.5 | 24.8 | 81.6 | 79.3 | 25.4 | 48.9 | |
| 10 | 92.2 | 30.4 | 92.9 | 32.1 | 91.7 | 87.2 | 32.4 | 58.7 | ||
| 12 | 93.5 | 35.3 | 92.7 | 37.4 | 91.8 | 88.7 | 37.4 | 62.4 |
| Species | Temperature (°C) | Time (min) | Hemicellulose | Cellulose | ||||
|---|---|---|---|---|---|---|---|---|
| A | Ea | R2 | A | Ea | R2 | |||
| Cupressus lusitanica | 0 | 0 | 2 × 109 | 77.9 | 0.999 | 4 × 1019 | 158.3 | 0.955 |
| 200 | 8 | 3 × 109 | 78.5 | 0.995 | 8 × 107 | 68.2 | 0.999 | |
| 10 | 4 × 109 | 79.8 | 0.993 | 9 × 107 | 68.5 | 0.999 | ||
| 12 | 5 × 109 | 80.8 | 0.994 | 5 × 107 | 65.7 | 0.999 | ||
| 225 | 8 | 2 × 1010 | 87.0 | 0.996 | 1 × 109 | 81.4 | 0.999 | |
| 10 | 1 × 1010 | 85.1 | 0.996 | 3 × 1017 | 177.7 | 0.997 | ||
| 12 | 6 × 109 | 84.3 | 0.998 | 9 × 1015 | 160.3 | 0.999 | ||
| 250 | 8 | 3 × 109 | 82.1 | 0.999 | 1 × 1016 | 161.1 | 0.998 | |
| 10 | 5 × 108 | 76.0 | 0.998 | 2 × 1025 | 267.6 | 0.973 | ||
| 12 | 6 × 107 | 68.2 | 0.996 | 6 × 1019 | 201.1 | 0.999 | ||
| Dipteryx panamensis | 0 | 0 | 2 × 108 | 66.3 | 0.979 | 2 × 108 | 324.7 | 0.977 |
| 200 | 8 | 2 × 1012 | 105.1 | 0.997 | 1 × 1012 | 113.4 | 0.997 | |
| 10 | 4 × 1013 | 118.2 | 0.997 | 3 × 1012 | 119.1 | 0.997 | ||
| 12 | 3 × 1013 | 117.6 | 0.997 | 6 × 1013 | 133.8 | 0.997 | ||
| 225 | 8 | 1 × 1014 | 124.5 | 0.998 | 7 × 1015 | 157.1 | 0.994 | |
| 10 | 1 × 1014 | 123.1 | 0.998 | 1 × 1019 | 193.4 | 0.985 | ||
| 12 | 1 × 1013 | 113.5 | 0.998 | 4 × 1016 | 164.9 | 0.991 | ||
| 250 | 8 | 3 × 1010 | 89.7 | 0.999 | 6 × 1018 | 189.2 | 0.989 | |
| 10 | 4 × 108 | 75.0 | 0.999 | 9 × 1021 | 227.4 | 0.982 | ||
| 12 | 2 × 107 | 63.5 | 0.998 | 2 × 1028 | 299.9 | 0.946 | ||
| Gmelina arborea | 0 | 0 | 3 × 1012 | 109.6 | 0.914 | 9 × 1015 | 146.8 | 0.777 |
| 200 | 8 | 7 × 108 | 72.3 | 0.989 | 2 × 109 | 77.6 | 0.993 | |
| 10 | 8 × 109 | 81.8 | 0.994 | 2 × 1011 | 96.8 | 0.986 | ||
| 12 | 1 × 1011 | 94.1 | 0.999 | 4 × 1022 | 223.1 | 0.899 | ||
| 225 | 8 | 3 × 1010 | 89.1 | 1.000 | 4 × 1024 | 244.7 | 0.810 | |
| 10 | 4 × 1010 | 90.2 | 0.999 | 2 × 1032 | 327.9 | 0.729 | ||
| 12 | 6 × 106 | 56.7 | 0.997 | 2 × 1029 | 298.7 | 0.760 | ||
| 250 | 8 | 3 × 108 | 73.1 | 0.997 | 1 × 1016 | 161.1 | 0.998 | |
| 10 | 1 × 106 | 51.5 | 0.999 | 2 × 1012 | 112.7.7 | 0.999 | ||
| 12 | 9 × 102 | 24.5 | 0.998 | 5 × 1027 | 280.2 | 0.856 | ||
| Tectona grandis | 0 | 0 | 2 × 109 | 79.3 | 0.997 | 4 × 1022 | 143.6 | 0.991 |
| 200 | 8 | 4 × 1011 | 100.7 | 1.000 | 9 × 1014 | 144.9 | 0.994 | |
| 10 | 3 × 1011 | 100.1 | 1.000 | 6 × 1016 | 167.4 | 0.990 | ||
| 12 | 1 × 1012 | 105.5 | 1.000 | 5 × 1019 | 198.6 | 0.971 | ||
| 225 | 8 | 3 × 1012 | 109.4 | 0.999 | 4 × 1012 | 118.9 | 0.996 | |
| 10 | 2 × 1012 | 108.0 | 0.999 | 7 × 1012 | 121.4 | 0.994 | ||
| 12 | 2 × 1012 | 108.5 | 0.999 | 4 × 1015 | 153.0 | 0.993 | ||
| 250 | 8 | 4 × 1011 | 101.83 | 0.999 | 4 × 1013 | 130.32 | 0.998 | |
| 10 | 4 × 108 | 75.58 | 0.999 | 7 × 1016 | 168.65 | 0.997 | ||
| 12 | 9 × 108 | 79.99 | 0.949 | 9 × 1028 | 306.93 | 0.935 | ||
| Vochysia ferruginea | 0 | 0 | 9 × 1010 | 93.16 | 0.998 | 3 × 1026 | 225.20 | 0.901 |
| 200 | 8 | 6 × 108 | 71.85 | 0.996 | 7 × 1011 | 104.78 | 1.000 | |
| 10 | 6 × 109 | 81.67 | 0.999 | 2 × 109 | 78.04 | 0.999 | ||
| 12 | 4 × 108 | 69.73 | 0.971 | 4 × 1024 | 244.37 | 0.996 | ||
| 225 | 8 | 4 × 1010 | 88.96 | 0.999 | 1 × 109 | 71.99 | 0.995 | |
| 10 | 5 × 109 | 80.92 | 0.999 | 1 × 1030 | 303.55 | 0.985 | ||
| 12 | 3 × 107 | 63.22 | 0.998 | 1 × 1020 | 198.03 | 0.993 | ||
| 250 | 8 | 2 × 1011 | 83.23 | 0.997 | 1 × 1013 | 105.55 | 0.990 | |
| 10 | 1 × 108 | 68.60 | 1.000 | 1 × 1027 | 274.73 | 0.981 | ||
| 12 | 2 × 106 | 53.10 | 0.999 | 1 × 1036 | 376.89 | 0.958 | ||
| Species | Temperature (°C) | Time (min) | Time Max. (min) | Dmax (% wt/min) |
|---|---|---|---|---|
| Cupressus lusitanica | 0 | 0 | 18.33 | 17.3 |
| 200 | 8 | 18.7 | 14.7 | |
| 10 | 18.8 | 14.1 | ||
| 12 | 18.7 | 14.8 | ||
| 225 | 8 | 18.6 | 16.1 | |
| 10 | 18.3 | 19.2 | ||
| 12 | 18.3 | 18.8 | ||
| 250 | 8 | 18.3 | 18.4 | |
| 10 | 17.0 | 8.2 | ||
| 12 | 17.6 | 15.1 | ||
| Dipteryx panamensis | 0 | 0 | 18.0 | 18.6 |
| 200 | 8 | 18.3 | 18.0 | |
| 10 | 18.3 | 18.2 | ||
| 12 | 18.3 | 19.5 | ||
| 225 | 8 | 18.3 | 20.7 | |
| 10 | 18.3 | 20.7 | ||
| 12 | 18.2 | 21.1 | ||
| 250 | 8 | 18.0 | 22.7 | |
| 10 | 18.0 | 20.5 | ||
| 12 | 17.8 | 16.1 | ||
| Gmelina arborea | 0 | 0 | 16.7 | 16.9 |
| 200 | 8 | 17.0 | 16.6 | |
| 10 | 16.8 | 14.5 | ||
| 12 | 16.5 | 17.3 | ||
| 225 | 8 | 16.6 | 17.9 | |
| 10 | 16.4 | 20.2 | ||
| 12 | 16.6 | 14.0 | ||
| 250 | 8 | 16.5 | 18.1 | |
| 10 | 14.4 | 13.7 | ||
| 12 | 16.6 | 8.4 | ||
| Tectona grandis | 0 | 0 | 17.7 | 15.8 |
| 200 | 8 | 17.9 | 14.6 | |
| 10 | 17.9 | 15.0 | ||
| 12 | 17.7 | 15.8 | ||
| 225 | 8 | 18.2 | 19.6 | |
| 10 | 18.1 | 19.3 | ||
| 12 | 18.0 | 20.1 | ||
| 250 | 8 | 18.1 | 20.2 | |
| 10 | 18.0 | 21.8 | ||
| 12 | 17.6 | 16.5 | ||
| Vochysia ferruginea | 0 | 0 | 16.5 | 14.5 |
| 200 | 8 | 17.0 | 15.7 | |
| 10 | 17.1 | 14.8 | ||
| 12 | 16.4 | 16.2 | ||
| 225 | 8 | 16.4 | 14.5 | |
| 10 | 16.4 | 16.6 | ||
| 12 | 16.8 | 16.4 | ||
| 250 | 8 | 17.0 | 17.3 | |
| 10 | 16.6 | 16.9 | ||
| 12 | 16.4 | 17.9 |
| Variable | Cupressus Lusitanica | Dipteryx panamensis | Gmelina arborea | Tectona grandis | Vochysia ferruginea | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C1 | C2 | C1 | C2 | C1 | C2 | C1 | C2 | |
| Ti (°C) | - | - | - | −0.92 ** | - | - | −0.83 ** | - | - | −0.80 ** |
| Tm (°C) | −0.90 ** | - | −0.89 * | - | - | - | - | - | - | - |
| Tf (°C) | - | - | - | 0.79 ** | - | - | - | −0.71 * | - | - |
| Tsh (°C) | −0.91 ** | - | −0.78 ** | - | - | - | 0.83 ** | - | - | - |
| Toffset(hc) (°C) | −0.96 ** | - | −0.69 ** | −0.69 * | - | - | - | 0.94 ** | - | −0.93 ** |
| Tonset(c) (°C) | - | −0.97 ** | −0.78 ** | - | - | −0.87 ** | - | 0.88 ** | - | −0.94 ** |
| Toffset(c) (°C) | - | 0.89 ** | - | 0.97 ** | - | - | - | −0.78 ** | −0.69 * | - |
| Tonset(hc) (°C) | 0.97 ** | - | 0.95 ** | - | - | - | - | −0.69 * | - | - |
| WTsh (%) | 0.97 ** | - | - | - | - | - | −0.91 ** | - | 0.76 * | - |
| WTi (%) | 0.81 * | - | - | −0.87 ** | - | −0.70 * | - | - | 0.96 ** | - |
| WTm (%) | 0.98 ** | - | 0.85 ** | - | 0.87 ** | - | −0.96 ** | - | 0.87 ** | - |
| WTf (%) | 0.98 ** | - | 0.80 ** | - | 0.96 ** | - | −0.98 ** | - | 0.91 ** | - |
| WTonset(hc) (%) | 0.69 * | - | - | −0.85 ** | - | −0.76 * | −0.66 * | 0.71 * | 0.78 ** | - |
| WToffset(hc) (%) | 0.97 ** | - | 0.91 ** | - | 0.93 ** | - | −0.93 ** | - | 0.68 * | - |
| WTonset(c) (%) | - | 0.79 * | 0.96 ** | - | - | 0.79 * | - | −0.88 * | - | 0.74 * |
| WToffset (c) (%) | 0.96 ** | - | 0.75 * | −0.64 * | 0.96 ** | - | −0.98 ** | - | 0.94 ** | - |
| Ea Hemicellulose | −0.72 * | - | −0.92 ** | - | −0.93 ** | - | - | 0.75 * | −0.73 * | - |
| Ea Cellulose | 0.73 * | - | 0.86 ** | - | 0.69 * | - | −0.74 * | - | 0.75 * | - |
| Residual mass (%) | 0.98 ** | - | 0.80 * | 0.96 ** | - | −0.98 ** | - | 0.91 ** | - | |
| Time max (min) | −0.77 * | - | −0.95 ** | - | −0.64 * | - | - | 0.74 * | - | - |
| Rate max (wt/%) | - | - | - | - | −0.83 ** | - | - | - | 0.79 * | - |
| Percentage of variance | 60.88 | 16.46 | 52.18 | 31.49 | 44.52 | 18.28 | 46.36 | 46.36 | 47.45 | 47.45 |
| Cumulative variance | 60.88 | 77.35 | 52.18 | 83.67 | 44.52 | 62.80 | 32.63 | 78.99 | 26.66 | 74.11 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaitán-Álvarez, J.; Moya, R.; Puente-Urbina, A.; Rodriguez-Zúñiga, A. Thermogravimetric, Devolatilization Rate, and Differential Scanning Calorimetry Analyses of Biomass of Tropical Plantation Species of Costa Rica Torrefied at Different Temperatures and Times. Energies 2018, 11, 696. https://doi.org/10.3390/en11040696
Gaitán-Álvarez J, Moya R, Puente-Urbina A, Rodriguez-Zúñiga A. Thermogravimetric, Devolatilization Rate, and Differential Scanning Calorimetry Analyses of Biomass of Tropical Plantation Species of Costa Rica Torrefied at Different Temperatures and Times. Energies. 2018; 11(4):696. https://doi.org/10.3390/en11040696
Chicago/Turabian StyleGaitán-Álvarez, Johanna, Róger Moya, Allen Puente-Urbina, and Ana Rodriguez-Zúñiga. 2018. "Thermogravimetric, Devolatilization Rate, and Differential Scanning Calorimetry Analyses of Biomass of Tropical Plantation Species of Costa Rica Torrefied at Different Temperatures and Times" Energies 11, no. 4: 696. https://doi.org/10.3390/en11040696
APA StyleGaitán-Álvarez, J., Moya, R., Puente-Urbina, A., & Rodriguez-Zúñiga, A. (2018). Thermogravimetric, Devolatilization Rate, and Differential Scanning Calorimetry Analyses of Biomass of Tropical Plantation Species of Costa Rica Torrefied at Different Temperatures and Times. Energies, 11(4), 696. https://doi.org/10.3390/en11040696